p38信號通路對SETD4在細胞中的表達和定位的影響*
2012-11-13 07:57:10蔡軍偉付曉霞劉亞偉陳小歡羅海華李煜生劉靖華
中國病理生理雜志 2012年5期
葉 萍, 蔡軍偉, 付曉霞, 崔 航, 劉亞偉, 陳小歡,羅海華, 李煜生, 劉 蕓, 姜 勇, 劉靖華
(重大疾病的轉錄組與蛋白質組學教育部重點實驗室,南方醫科大學基礎醫學院病理生理學教研室,廣東廣州510515)
SET是一高度保守的結構域,因最初在果蠅的3個調節因子suppressor of variegation 3-9[Su(var)3-9]、enhancer of zeste[E(z)]和 trithorax的羧基末端發現而得名。含有SET結構域的蛋白被稱為SET家族蛋白。SET家族蛋白廣泛分布于酵母和動植物細胞中,目前經過分離鑒定以及理論推測的SET家族蛋白有 300多種,其中人類有48種[1-2]。
2000年,Stephen首次發現含有SET結構域的蛋白家族具有組蛋白甲基化轉移酶的功能,即通過對組蛋白的甲基化修飾作用(主要作用于組蛋白H3/H4上的賴氨酸位點)影響染色體結構,調控基因表達[3-4]。近期研究發現,SET家族蛋白不僅具有組蛋白甲基化酶的作用,同時對非組蛋白也有甲基化酶的作用。如SET家族蛋白SET9、SMYD2可甲基化p53的不同賴氨酸位點,SETD6能甲基化核轉錄因子RelA(p65)310位賴氨酸,從而抑制炎癥因子白細胞介素6(interleukin-6,IL-6)、IL-1和 IL-8等釋放[5-10]。越來越多的研究表明SET家族蛋白通過上述甲基化酶作用參與了炎癥及腫瘤的發生發展[11-12]。因此,探討SET家族蛋白的功能對于闡明各種炎癥相關性疾病及腫瘤的發生發展機制具有重要的理論價值。
SETD4(SET domain containing 4)屬于SET家族蛋白中的一種,但是目前對其鮮見報道。為了探討SETD4的生物學功能,本研究首先對SETD4在不同細胞中的表達和分布特點進行了觀察,接下來觀察了亞砷酸鈉(sodium arsenite,NaAsO2)刺激對 SETD4蛋白表達水平和定位的影響,并初步研究上述改變是否與p38 MAPK信號通路有關。
材料和方法
1 材料
CO2培養箱(Heraeus),細胞培養皿(Corning),DMEM培養基(Gibco-BRL),胎牛血清(HyClone),SETD4多克隆抗體(Santa Cruz,編號 sc-133993,可檢測人或鼠SETD4),β-actin抗體(Bioworld Technology),anti-rabbit抗體(Cell Signaling Technology),ReverTra Ace qPCR RT kit(Toyobo),Alexa Fluor 594偶聯的抗兔IgG(Molecular Probes),細胞裂解液(CST),引物合成由深圳華大基因公司完成。小鼠胚胎成纖維細胞(wild-type mouse embryonic fibroblasts, MEFs, C57B1/6 background)p38+/+和p38-/-細胞系由美國Scripps研究所韓家淮博士饋贈。p38+/+細胞表達正常的 p38蛋白,p38-/-細胞則不表達 p38 蛋白[13]。
2 方法
2.1 細胞培養 NIH 3T3(小鼠成纖維細胞)、293T(人胚胎腎細胞)、Raw 264.7(小鼠巨噬細胞)、QSG(人正常肝臟細胞)、HepG2(人肝臟腫瘤細胞)、SMMC(人源肝癌細胞)、U2-OS(小鼠骨肉瘤細胞)(以上細胞由本室保存)、p38+/+和 p38-/-細胞均用含有10%FBS的DMEM培養基在CO2培養箱(5%CO2,37 ℃)中培養。
2.2 NaAsO2刺激細胞實驗 細胞接種在6 cm培養皿中,待細胞長至80%融合時,無血清培養6 h,再給予100 μmol/L NaAsO2刺激,分別在刺激后 0、0.5、1、2、3和6 h收集細胞,按下述方法提取細胞總蛋白。
2.3 Western blotting檢測SETD4蛋白的表達 將收集的細胞用細胞裂解緩沖液(20 mmol/L Tris-HCl pH7.5,150 mmol/L NaCl,1 mmol/L Na2EDTA,1 mmol/L EGTA,10%Triton,2.5 mmol/L 焦磷酸鈉,1 mmol/Lβ-磷酸甘油,1 mmol/L釩酸鈉)裂解細胞,提取細胞總蛋白。總蛋白用Bradford法蛋白定量,取等量的總蛋白行12%SDS-PAGE蛋白凝膠電泳,電泳結束后將蛋白電轉移至PVDF膜。經5%脫脂奶粉TBST溶液室溫封閉2 h后,再與1∶1 000稀釋的I抗4℃孵育過夜,然后與1∶1 000稀釋的HRP(辣根過氧化物酶)偶聯的抗兔IgG室溫孵育1 h,采用Pierce公司的SuperSignal West Pico化學發光底物進行顯色,利用Kodak IS4000R圖像工作站[Carestream公司(原Kodak)]進行化學發光檢測。
2.4 細胞免疫熒光實驗 接種1×105細胞至Petri皿中,6 h后補液,生長12~18 h,待細胞密度達到80%時換用無血清DMEM培養6 h,NaAsO2刺激組給予100μmol/L NaAsO2刺激1 h。棄去細胞上清,PBS洗滌細胞3次,以4%多聚甲醛固定細胞10 min,PBS 洗3 次,5 min/次,然后用含 0.1%Triton X-100的 PBS(PBST)透化細胞膜5 min,PBS洗3次,5 min/次,用質量分數 0.1%硼氫化鈉(NaBH4)孵育細胞5 min,PBST洗3次,5 min/次,用3%BSA封閉1 h,PBST洗3次后用抗-SETD4特異性抗體(1∶500稀釋)4°孵育過夜,次日取出室溫孵育1 h,PBST洗3次后,用 Alexa Flour-594偶聯的抗兔IgG(1∶1 000稀釋)室溫孵育1 h,DAPI染核10 min,PBS洗3次,5 min/次,然后PBS封片,于Zessi熒光顯微鏡下觀察和圖像采集。
3 SETD4的結構預測
小鼠SETD4(accession number:P58467)和人的SETD4(accession number:NP_059134)一級結構信息來源于美國國家生物技術信息中心(the National Center for Biotechnology Information,NCBI);小鼠SETD4蛋白的二級結構采用 PSIPRED 3.0預測(http://bioinf.cs.ucl.ac.uk/psipred/)。
結 果
1 SETD4的結構特征
小鼠SETD4由439個氨基酸組成,人SETD4是440個氨基酸,兩者序列的一致性為77%(338/440),保守性為87%(382/440)。SETD4主要由2個結構域組成,位于羧基末端的SET結構域(小鼠:第33~278位氨基酸;人:第34~279位氨基酸)和位于氨基端末端的底物結合結構域(小鼠:第306~424位氨基酸;人:第307~425位氨基酸),見圖1A。小鼠SETD4的二級結構圖有16個α螺旋(α-helix,粉紅色)、8個 β折疊結構(β -strand,黃色)以及不規則卷曲組成,見圖1B。
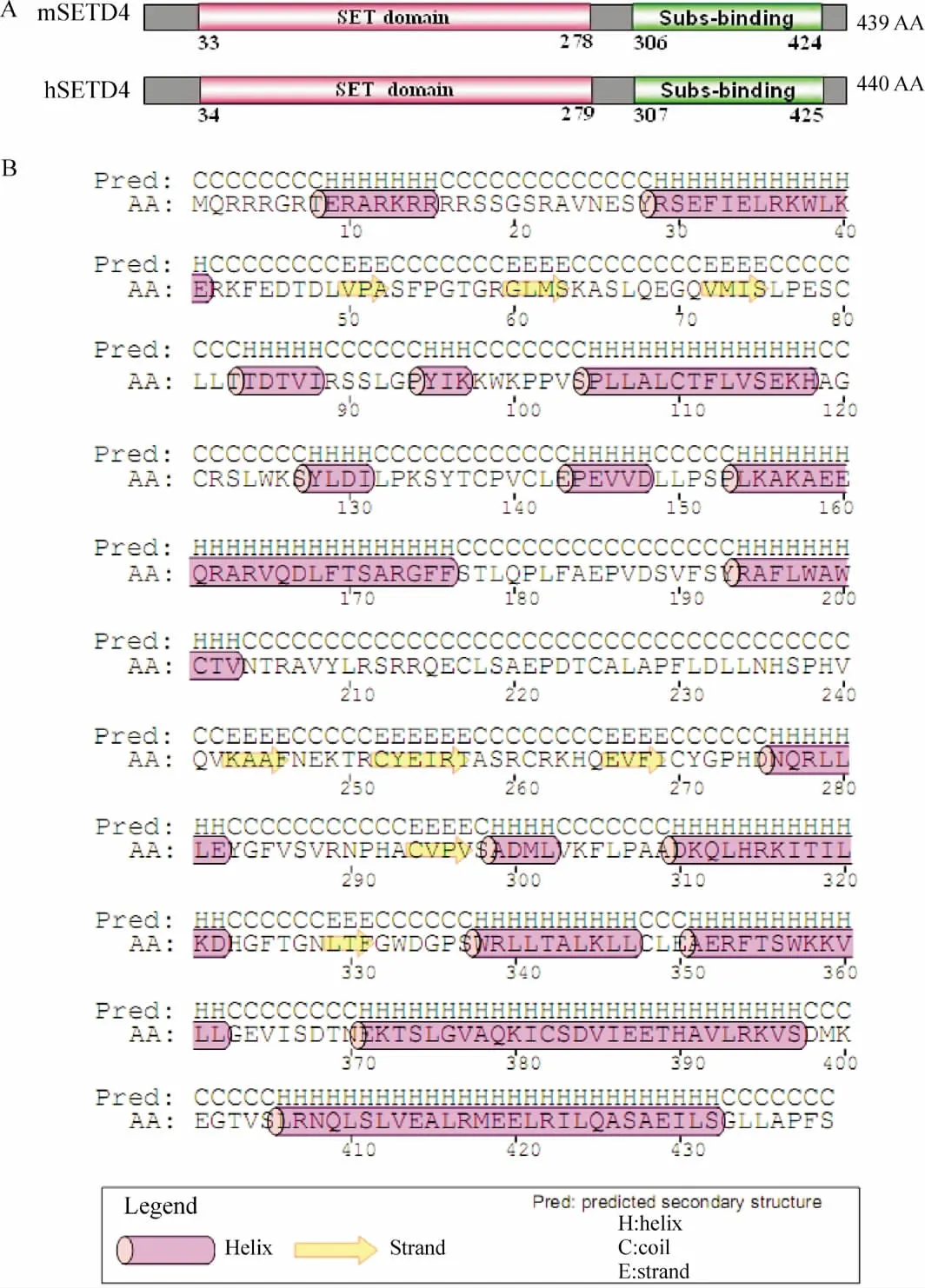
Figure 1.Characterization of SETD4.A:the domains of mouse SETD4(mSETD4)and human SETD4(hSETD4);B:the secondary structure of mouse SETD4.Subs-binding:substrate-binding domain.圖1 SETD4的結構特征
2 SETD4在不同細胞中的表達
SETD4蛋白在鼠源(Raw 264.7、U2 - OS、NIH 3T3和MEFs)和人源(293T、QSG、HepG2和 SMMC)的細胞中均有表達,在總蛋白量相同的情況下鼠源細胞的表達水平較人源的高,并且鼠源細胞在40 kD附近有2條帶,而人源的只有分子量較大的1條帶,見圖2。這提示盡管SETD4在不同種屬、不同組織來源的細胞均有表達,但是其表達特征有一定區別。
3 SETD4在細胞中的定位
細胞免疫熒光結果顯示SETD4在整個細胞中分布,以胞質較多,見圖3。
4 p38 MAPK信號通路對NaAsO 2誘導的細胞SETD4蛋白表達的影響
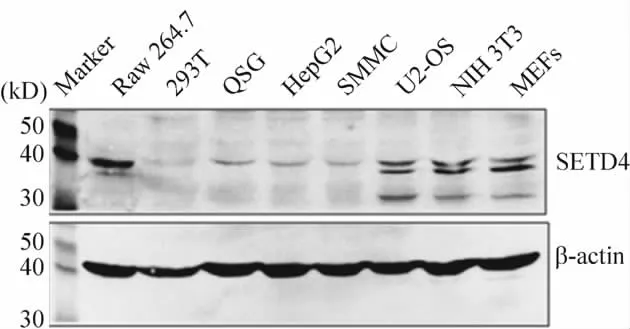
Figure 2.Expression of SETD4 in different cells.圖2 SETD4蛋白在不同細胞中的表達
p38+/+細胞受NaAsO2刺激后其SETD4蛋白的表達在0.5 h、1 h、2 h、3 h 沒有明顯變化,但在6 h 時明顯下降;p38-/-細胞受 NaAsO2刺激后其 SETD4蛋白的表達趨勢與p38+/+一致,即均在6 h時有明顯減少,提示p38信號通路可能沒有參與NaAsO2誘導的SETD4表達的改變,見圖4。
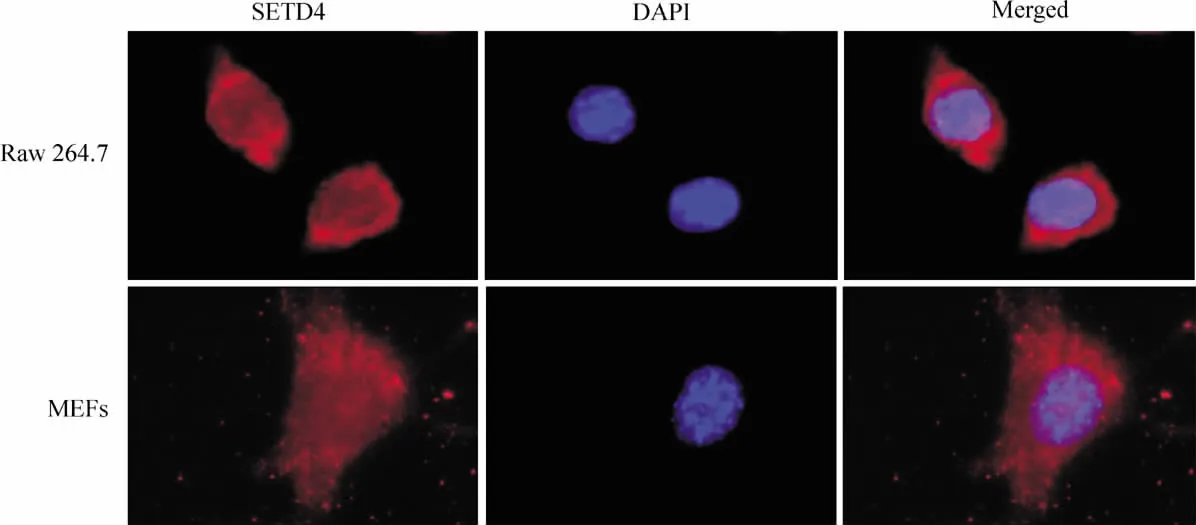
Figure 3.The location of SETD4 in Raw 264.7 cells and MEFs(immunofluorescent staining,×600).圖3 SETD4在NIH 3T3、Raw264.7細胞中的定位

Figure 4.Expression of SETD4 protein in p38+/+and p38 -/-cells following NaAsO2 treatment.圖4 NaAsO2刺激對p38+/+和p38-/-細胞 SETD4蛋白表達的影響
5 NaAsO2誘導的SETD4蛋白移位入核依賴于p38信號通路
實驗發現,靜息狀態下 SETD4在 p38+/+、p38-/-細胞的胞漿和胞核中均有分布,但主要分布于胞漿中;100μmol/L NaAsO2刺激1 h后,p38+/+細胞核內 SETD4明顯增多,但是 p38-/-細胞中SETD4的分布沒有明顯改變,提示NaAsO2能誘導SETD4蛋白移位入核,并且依賴于p38信號通路。另外,NaAsO2刺激后p38+/+和p38-/-細胞中SETD4蛋白在胞漿呈現點狀聚集,見圖5。
討 論
我們的實驗結果發現SETD4蛋白表達水平(含量)與種屬有關,即在總蛋白量相同的情況下鼠源細胞SETD4蛋白表達水平(含量)較人源的高,而與細胞是否是腫瘤細胞關系不大,提示SETD4參與腫瘤的發生發展可能主要是通過改變活性形式而不是上調表達水平。SETD4的理論分子量是49 kD,但是我們在40 kD附近檢測到鼠源細胞有2條帶,而人源的只有分子量較大的1條帶,提示SETD4的遷移率發生了改變,這可能與其剪切形式發生了改變或者發生了翻譯后修飾有關。生物信息學分析發現,SETD4可能有多種不同的剪切形式,本實驗觀察到的結果屬于哪種情況還有待實驗的進一步證實。
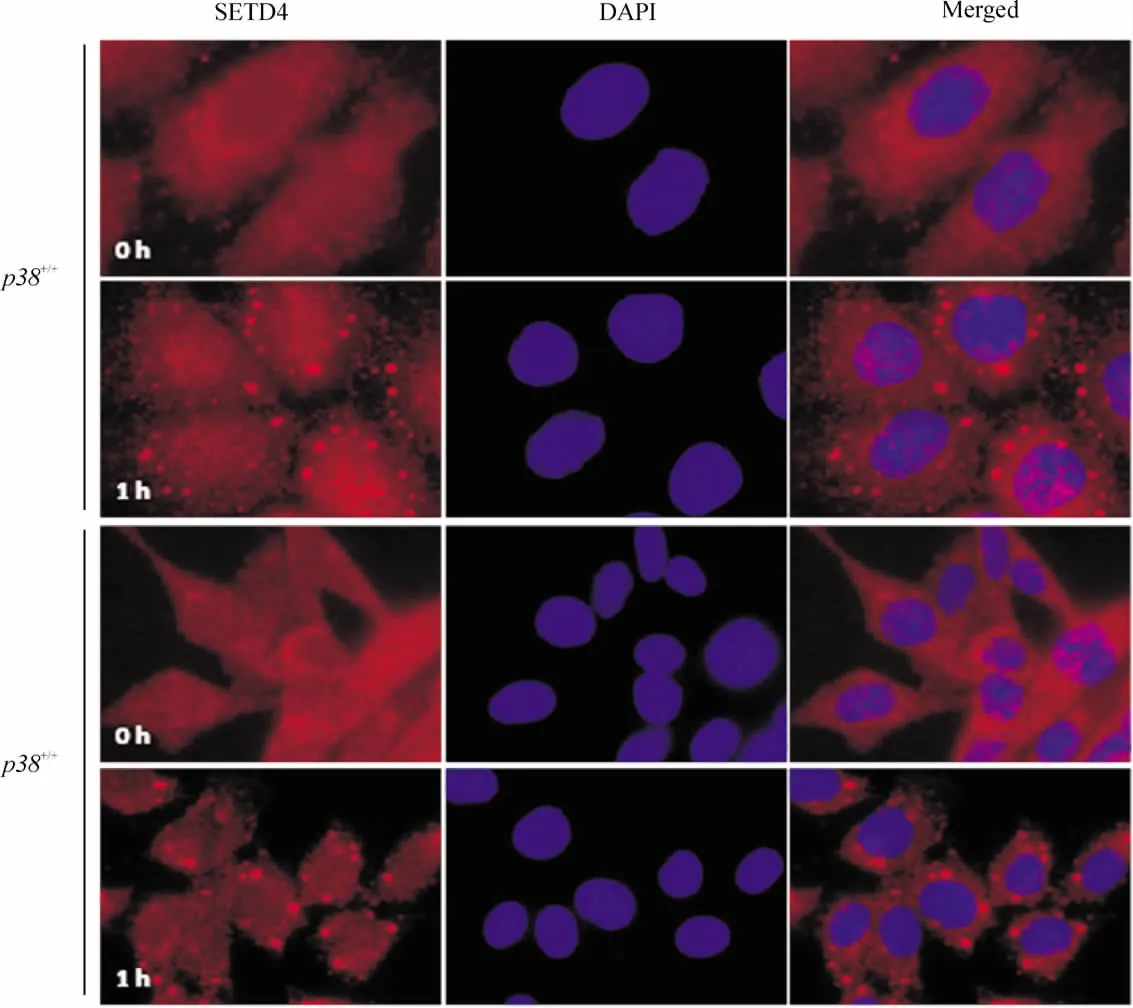
Figure 5.Localization and translocation of SETD4 in p38+/+and p38 -/- cells following NaAsO2 treatment(immunofluorescent staining,×600) .圖5 NaAsO2對p38+/+和p38-/-細胞SETD4蛋白的定位及移位的影響
p38是絲裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)家族中的一個重要成員,參與多種生理和病理過程的調節[14]。在不同刺激條件下,p38 MAPK可通過不同方式活化來介導不同細胞的生物學功能。例如,p38可分別通過磷酸化轉錄因子、細胞骨架相關蛋白及酶等多種底物來調節應激、凋亡、炎癥和細胞生長、分化等過程,而且還參與細胞惡性轉化(malignant transformation)和腫瘤細胞侵襲轉移(tumor invasion and metastasis)等病理過程的調控[15]。由于SET家族蛋白與炎癥和腫瘤的發生發展有很大關系,因此,本研究用p38+/+和p38-/-細胞首先探討了p38 MAPK對亞砷酸鹽刺激后細胞SETD4表達的影響。結果發現無論是否有p38的存在,當細胞受到NaAsO2刺激后SETD4蛋白表達趨勢一致,提示p38 MAPK對SETD4的表達可能不發揮主要的調控作用。接下來,我們研究了p38 MAPK對亞砷酸鹽刺激后細胞SETD4定位及其移位的影響。結果表明,如果p38缺失,細胞被激活后SETD4蛋白就不能從細胞質向細胞核移位,提示SETD4的移位有賴于p38 MAPK通路。信號蛋白的亞細胞定位和激活后移位已成為細胞信號轉導研究中的重要內容。目前認為,信號蛋白的定位和激活后移位以及伴隨的與其它蛋白的可逆性結合是信號在細胞內進行有效傳遞的關鍵所在。在很多情況下,信號蛋白需要錨定于適配體蛋白、細胞骨架及細胞質膜結構,或與其上游激酶和下游底物結合來發揮正常功能。信號分子的特異性定位有助于細胞高效地完成各種正常功能和病理反應。因此,進一步明確p38 MAPK如何影響SETD4的移位及SETD4結合蛋白對闡明SETD4的功能,進而探索其在炎癥和腫瘤發生發展中扮演的角色有非常重要的意義。
[1] Xiao B,Wilson JR,Gamblin SJ.SET domains and histone methylation [J].Curr Opin Struct Biol,2003,13(6):699-705.
[2] Eom GH,Kim KB,Kim JH,et al.Histone methyltransferase SETD3 regulates muscle differentiation[J].J Biol Chem,2011,286(40):34733 -34742.
[3] Rea S,Eisenhaber F,O'Carroll D,et al.Regulation of chromatin structure by site-specific histone H3 methyltransferase[J].Nature,2000,406(6796):593 -599.
[4] Levy D,Kuo AJ,Chang Y,et al.Lysine methylation of the NF-κB subunit RelA by SETD6 couples activity of the histone methyltransferase GLPat chromatin to tonic repression of NF - κB signaling[J].Nat Immunol,2011,12(1):29-36.
[5] Chen X,El Gazzar M,Yoza BK,et al.The NF - κB factor RelB and histone H3 lysine methyltransferase G9a directly interact to generate epigenetic silencing in endotoxin tolerance[J].JBiol Chem,2009,284(41):27857 -27865.
[6] Bai Q,Shen Y,Yao X,et al.Modeling a new water channel that allows SET9 to dimethylate p53[J].PLoS One,2011,6(5):e19856.
[7] Yang XD,Huang B,Li M,et al.Negative regulation of NF-κB action by Set9-mediated lysine methylation of the RelA subunit[J].EMBO J,2009,28(8):1055 -1066.
[8] Chang Y,Levy D,Horton JR,et al.Structural basis of SETD6-mediated regulation of the NF-κB network via methyl- lysine signaling[J].Nucleic Acids Res,2011,39(15):6380-6389.
[9] Kim DW,Kim KB,KM JY,et al.Characterization of a novel histone H3K36 methyltransferase setd3 in zebrafish[J].Biosci Biotechnol Biochem,2011,75(2):289 -294.
[10] Huang J,Dorsey J,Chuikov S,et al.G9a and Glp methylate lysine 373 in the tumor suppressor p53[J].J Biol Chem,2010,285(13):9636 -9641.
[11] Bayarsaihan D.Epigenetic mechanisms in inflammation[J].J Dent Res,2011,90(1):9 -17.
[12] Wilson AG.Epigenetic regulation of gene expression in the inflammatory response and relevance to common diseases[J].JPeriodontol,2008,79(8 Suppl):1514 -1519.
[13] 劉愛華,龔小衛,魏 潔,等.P38 MAPK基因敲除對亞砷酸鹽誘導細胞凋亡的影響[J].中國病理生理雜志,2008,24(5):892-895.
[14] 龔小衛,姜 勇.絲裂原活化蛋白激酶(MAPK)生物學功能的結構基礎[J].中國生物化學與分子生物學報,2003,19(1):5-11.
[15] Lee JC,Young PR.Role of CSB/p38/RK stress response kinase in LPS and cytokine signaling mechanisms[J].J Leukoc Biol,1996,59(2):152-157.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
學苑創造·A版(2020年9期)2020-10-13 09:41:02
中國生殖健康(2019年3期)2019-02-01 06:12:26
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
海軍航空大學學報(2015年3期)2015-11-11 17:20:00
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
