果蠅Mef2基因多克隆抗體的制備及檢測
2012-11-22 01:32:46徐臘梅盛立翔吳秀山莫小陽
湖南師范大學(xué)自然科學(xué)學(xué)報(bào) 2012年5期
關(guān)鍵詞:融合
朱 玲,徐臘梅,盛立翔,吳秀山,莫小陽
(湖南師范大學(xué)蛋白質(zhì)化學(xué)及魚類發(fā)育生物學(xué)教育部重點(diǎn)實(shí)驗(yàn)室,心臟發(fā)育研究中心,中國 長沙 410081)
Mef2屬于MADS家族轉(zhuǎn)錄因子基因,MADS家族是指其蛋白質(zhì)結(jié)構(gòu)域含有高度保守的MADS結(jié)構(gòu)域的一類轉(zhuǎn)錄因子的總稱[1].MADS結(jié)構(gòu)域是一種DNA結(jié)合結(jié)構(gòu)域,由55~63個(gè)氨基酸組成,所識別并結(jié)合的DNA保守序列為CArG框[CC(A/T)6GG].
Mef2位于果蠅2號染色體上,mRNA全長5 990 bp,編碼501個(gè)氨基酸.脊椎動物有4種MEF2蛋白(MEF2A-D),而果蠅中只有1種MEF2蛋白(D-MEF2)[2].不管是脊椎動物還是果蠅中的MEF2蛋白都含有一個(gè)高度保守的MADS和MEF2結(jié)構(gòu)域,并且由它們來介導(dǎo)DNA結(jié)合、二聚作用、蛋白質(zhì)-蛋白質(zhì)的相互作用[3].
Mef2最初是在分化的肌肉細(xì)胞中作為一個(gè)肌肉特異結(jié)合的活性因子被鑒定的,1997 年卻發(fā)現(xiàn)Mef2基因在心臟發(fā)育的過程中起重要作用[4].在果蠅中,Mef2首先在原腸胚形成時(shí)在整個(gè)中胚層中表達(dá),隨著胚胎的發(fā)育,它的表達(dá)逐漸限制在體節(jié)、內(nèi)臟和心肌細(xì)胞中[5-6].鑒于Mef2基因在果蠅心臟發(fā)育的過程中起著重要的作用[7],所以制備一個(gè)特異性好的多克隆抗體尤其重要,對進(jìn)一步以果蠅為模型研究Mef2基因具有重要的意義.
1 材料與方法
1.1 實(shí)驗(yàn)材料和試劑
大腸桿菌Rosseta菌種,pET-28a菌種以及coliDH5α菌種為本實(shí)驗(yàn)室提供;限制性內(nèi)切酶EcoRⅠ和XhoⅠ,DNA、蛋白質(zhì)相對分子質(zhì)量Marker,Tag DNA聚合酶購自深圳晶美公司;pMD18-T載體和連接酶購自大連TaKaRa公司;UNIQ-10柱式DNA 膠回收純化試劑盒購自上海生工公司;質(zhì)粒提取試劑盒購自O(shè)MEGA公司;Glutathione SepharoseTM4B蛋白純化試劑盒購自PIERCE公司;弗氏佐劑購自Sigma公司;丙烯酰胺、甲叉雙丙烯酰胺、IPTG(異丙基-β-D-硫代半乳糖苷)等購自上海Sangon公司;新西蘭大白兔購自中南大學(xué)實(shí)驗(yàn)動物學(xué)部.
1.2 引物設(shè)計(jì)與合成
從NCBI數(shù)據(jù)庫(http://www.ncbi.nlm.nih.gov)檢索出果蠅Mef2基因的序列,根據(jù)巢式PCR的原理利用Primer Premier 5.0 軟件設(shè)計(jì)兩對引物:第一對:D-Mef2-S1:5′CGCCAGCTGCCGGAAATCCA 3′;D-Mef2-A1:5′GCGTCTGCAGCGTGACCACT 3′.第二對(分別加入EcoRⅠ和XhoⅠ酶切位點(diǎn)):D-Mef2-S2:5′GAGAATTCATGGGCCGCAAA AAAATTCA 3′EcoRⅠ;D-Mef2-A2:5′GGCTCGAGGTGGACTGGCCTGCAGCAGG 3′XhoⅠ.
1.3 基因克隆和載體構(gòu)建
選取30只野生型果蠅,液氮冷凍30 min,用Trizol法提取總RNA[8],反轉(zhuǎn)錄PCR擴(kuò)增,獲得first-chain cDNA.然后以first-chain cDNA為模板用第一對引物進(jìn)行PCR擴(kuò)增,然后以所得產(chǎn)物為模板,用第二對引物進(jìn)行PCR擴(kuò)增,得到目的片段[9].目的片段經(jīng)純化后克隆入T載體,轉(zhuǎn)化入DH5α感受態(tài)細(xì)胞中,酶切檢測篩選出陽性單克隆,經(jīng)測序鑒定后將DNA片段從pMD18-T-Mef2質(zhì)粒上切下,連入pET-28a載體(EcoRⅠ和XhoⅠ酶切線性化),得到陽性克隆,經(jīng)EcoRⅠ和XhoⅠ雙酶切鑒定后,得到重組表達(dá)質(zhì)粒pET-28a-Mef2.
1.4 Mef2融合蛋白誘導(dǎo)表達(dá)及純化
pET-28a-Mef2重組表達(dá)質(zhì)粒轉(zhuǎn)化入大腸桿菌菌株Rosseta,篩選出陽性菌落,然后接種于LB 培養(yǎng)基(含100 mg/L 氯霉素,卡那霉素)37 ℃ 培養(yǎng)12 h左右,以1∶50轉(zhuǎn)接擴(kuò)大培養(yǎng)至OD600達(dá)0.6,按0.1 mol/L 加IPTG,25 ℃誘導(dǎo),4、5、6、7 h各取1 mL菌液,確定最佳誘導(dǎo)時(shí)間.
收集誘導(dǎo)后大腸桿菌,PBS漂洗后超聲裂解,離心取上清.4 ℃下與經(jīng)Binding Buffer漂洗活化后的Ni-IDA凝膠柱結(jié)合,Washing Buffer洗去雜蛋白,Elution Buffer洗脫目的蛋白,獲得純化的His-Mef2融合蛋白,-80 ℃保存?zhèn)溆茫?/p>
1.5 Mef2多克隆抗體制備
將純化后的His-Mef2蛋白與弗氏完全佐劑按體積1∶1在注射器中推成乳劑,分散約12個(gè)點(diǎn)對同一新西蘭大白兔進(jìn)行背部皮下免疫注射.在第一次注射后14、21、28 d,將His-Mef2蛋白按體積比1∶1與弗氏不完全佐劑在注射器中推成乳劑,進(jìn)行多次免疫.第35天主動脈取血,靜置過夜(4 ℃),3 000 r/min離心處理10 min,取上清分裝保存(-80 ℃).
1.6 效價(jià)測定
獲得免疫兔血清后,用誘導(dǎo)的總蛋白進(jìn)行Western blot實(shí)驗(yàn)檢測抗體效價(jià).將含pET-28a-Mef2重組子的菌株在LB培養(yǎng)液中37 ℃培養(yǎng),將5 mL接種于200 mL新鮮培養(yǎng)基中,用IPTG在25 ℃條件下誘導(dǎo)5 h后,收集細(xì)菌,PBS重懸,然后用SDS-PAGE電泳分離.將Mef2抗體分別以1∶100、1∶500、1∶1 000和1∶2 000的體積比稀釋后,以免疫前兔血清作為對照,采用Western blotting方法檢測抗體的特異性[10].
2 結(jié)果與分析
2.1 重組質(zhì)粒pET-28a-Mef2的構(gòu)建與鑒定
將純化后的果蠅Mef2基因片段克隆到pMD18-T載體中,經(jīng)測序鑒定與基因理論序列100%匹配,然后用EcoRⅠ和XhoⅠ雙酶切,連入pET-28a載體(EcoRⅠ和XhoⅠ酶切線性化),獲得重組質(zhì)粒pET-28a-Mef2.經(jīng)酶切和測序鑒定重組質(zhì)粒正確(圖1).
2.2 Mef2 融合蛋白誘導(dǎo)表達(dá)及純化
將重組子轉(zhuǎn)入Rosseta感受態(tài)細(xì)胞中,以0.1 mmol/L IPTG進(jìn)行誘導(dǎo),30 ℃條件下進(jìn)行時(shí)間梯度誘導(dǎo)1,2,3,4,5,6 h分別取樣檢測蛋白的表達(dá),結(jié)果(如圖2)顯示Mef2蛋白在誘導(dǎo)1 h后就開始表達(dá),隨著時(shí)間推移表達(dá)量增加,在3 h后趨于穩(wěn)定.
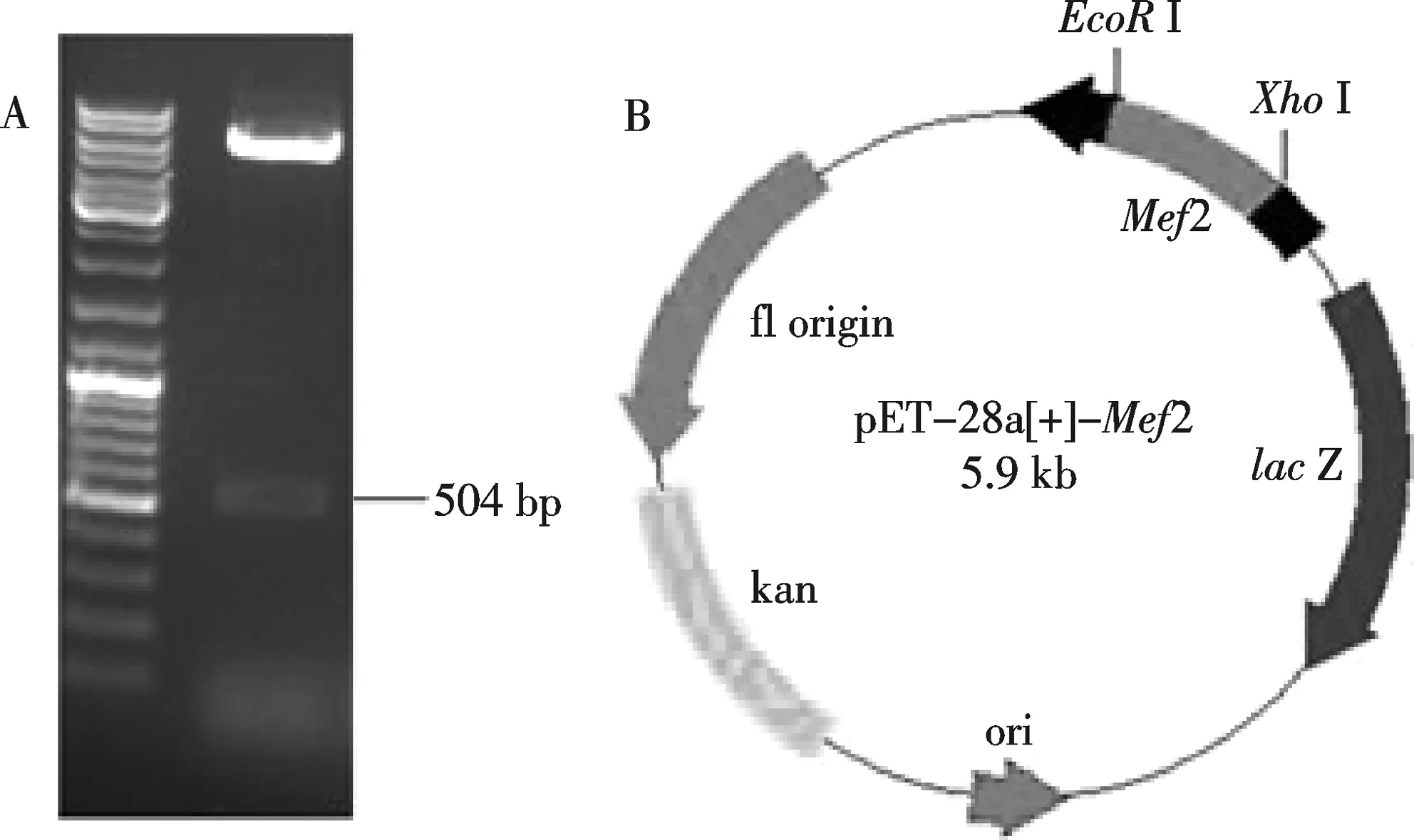
A: pET-28a-Mef2質(zhì)粒雙酶切鑒定,B:pET-28a-Mef2質(zhì)粒示意圖圖1 pET-28a-Mef2質(zhì)粒酶切鑒定圖譜和示意圖

His-Mef2融合蛋白時(shí)間梯度誘導(dǎo), IPTG 0.1 mmol/L, 溫度30 ℃. 1:Marker; 2:pET-28a-Mef2誘導(dǎo)前;3~8:0.1 mmol/L IPTG誘導(dǎo)pET-28a-Mef2 蛋白表達(dá)1~6 h.圖2 His-Mef2 融合蛋白的表達(dá)
選定最佳誘導(dǎo)條件(30 ℃、0.1 mmol/L IPTG 濃度、3 h),大量誘導(dǎo)獲得濃度較高的蛋白,超聲裂解后經(jīng)Ni-IDA凝膠柱親和純化,獲得比較純的蛋白(圖3),可以進(jìn)行蛋白免疫[11].
2.3 抗血清的制備與效價(jià)檢測
用純化的目的蛋白免疫新西蘭大白兔獲得血清,用誘導(dǎo)的總蛋白進(jìn)行效價(jià)測定,抗體稀釋體積比分別為1∶100、1∶500、1∶1 000和1∶2 000,免疫前兔血清(control)為對照.結(jié)果發(fā)現(xiàn)抗體的效價(jià)較好,滿足實(shí)驗(yàn)所需 (圖4).

圖3 His-Mef2 融合蛋白的純化
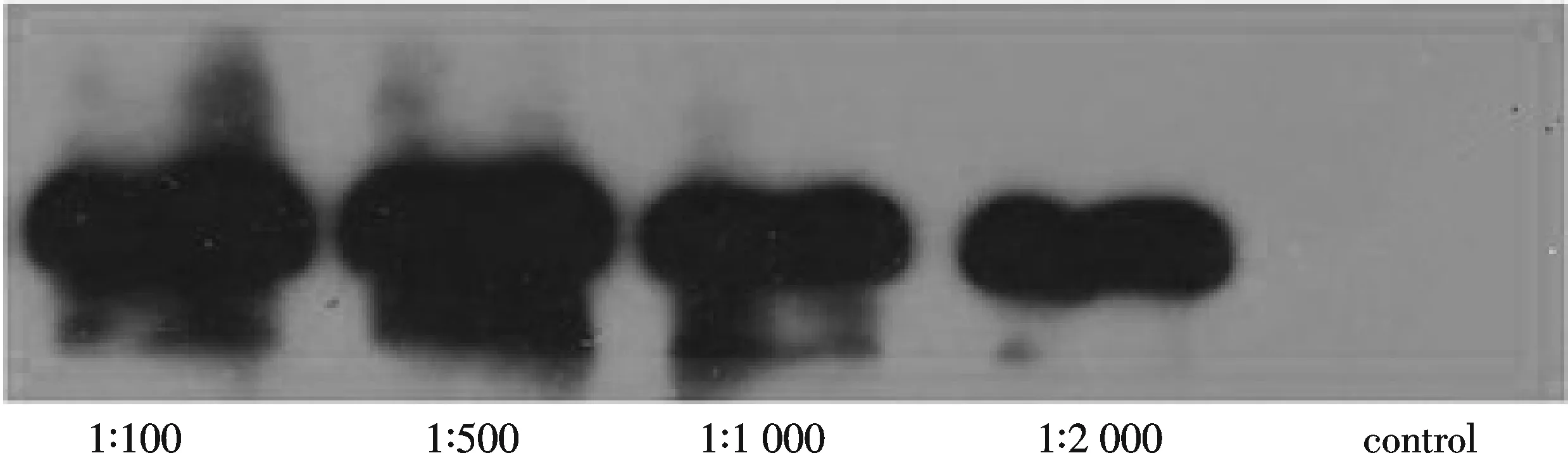
圖4 Mef2 抗體效價(jià)測定
3 討論
為了以果蠅為模型進(jìn)一步研究Mef2基因的功能,本文通過試驗(yàn)獲得了Mef2原核表達(dá)融合蛋白,制備了高效價(jià)和特異性好的兔抗Mef2血清.在獲得融合蛋白的過程中,先通過軟件分析選取了親水性好而又含結(jié)構(gòu)域少的一段序列,為獲得特異性好的抗體提供了前提條件[12].在誘導(dǎo)蛋白的過程中,IPTG 的終濃度選取的是0.1 mmol/L ,蛋白表達(dá)量相對高,又不至于因IPTG 濃度太高對細(xì)胞的生長產(chǎn)生毒性作用,試驗(yàn)相對容易成功.
pET-28a(+)載體是一種常用的原核表達(dá)載體,具有Kan抗性,質(zhì)粒大小為5.3 kb.載體的N端攜帶His標(biāo)簽、凝血酶基因及T7啟動子標(biāo)簽序列;在C端具有His標(biāo)簽序列, pET-28a(+)載體可在BL21(DE3)、Rosstea等表達(dá)菌株中高表達(dá)融合有His標(biāo)簽的蛋白,便于下游蛋白純化.pET-28a這種載體沒有本身溶解性高的多肽融合蛋白,也沒有催化二硫鍵形成的酶融合蛋白,而且不含信號肽序列,所以同一種蛋白用這個(gè)載體,形成包涵體的可能性更大些[13].
本試驗(yàn)中,作者通過構(gòu)建pET-28a-Mef2重組質(zhì)粒、誘導(dǎo)表達(dá)His-Mef2融合蛋白獲得了高純度的蛋白并免疫新西蘭大白兔制備了Mef2抗體.通過效價(jià)檢測證明制備的Mef2多克隆抗體具有較好的特異性,能滿足試驗(yàn)要求. 因此,它為將來運(yùn)用染色質(zhì)免疫沉淀、免疫共沉淀、免疫組化等手段深入研究Mef2基因的功能奠定了基礎(chǔ).
參考文獻(xiàn):
[1] SOLER C, HAN J, TAYLOR M V,etal.The conserved transcription factor Mef2 has multiple roles in adult Drosophila musculature formation[J]. Development, 2012,139(7):1270-1275.
[2] MILLIGAN J N, JOLLY E R. Identification and characterization of a Mef2 transcriptional activator in schistosome parasites.[J]. PloS Negl Trop Dis, 2012,6(1):e1443.
[3] FLAVELL S W, COWAN C W, KIM T K,etal. Activity-dependent regulation of MEF2 transcription factors suppresses excitatory synapse number[J]. Science, 2006,311(5763):1008-1012.
[4] BLANCHARD F J, COLLINS B, CYRAN S A,etal. The transcription factor Mef2 is required for normal circadian behavior in Drosophila[J]. J Neurosci, 2010,30(17):5855-5865.
[5] 吳秀山,袁婺洲,王躍群,等.心臟發(fā)育研究[M].長沙:湖南科學(xué)技術(shù)出版社, 2004:214-302.
[6] 吳秀山.心臟發(fā)育概論[M]. 北京:科學(xué)出版社, 2006:86-103.
[7] WESSELLS R J, BODMER R. Age-related cardiac deterioration: insights from drosophila [J]. Biosci, 2007,12:39-48.
[8] WU X, GOLDEN K, BODMER R,etal. Heart development in drosophila requires the segement polarity genewingless [J]. Dev Biol, 1996,177:104-116.
[9] LI S, ZHOU D Y, LU M M,etal. Advanced cardiac morphogenesis does not require heart tube fusion[J].Science, 2004,305(5690):1619-1622.
[10] 龔 琳,吳秀山,李永青,等.果蠅Nulp1特異性多克隆抗體的制備及檢測[J].湖南師范大學(xué)自然科學(xué)學(xué)報(bào), 2011,34(4):69-73.
[11] 閔 璐, 周云雷, 周 煌,等.lrrc10蛋白的表達(dá)、純化及多克隆抗體的制備[J].湖南師范大學(xué)自然科學(xué)學(xué)報(bào), 2011,34(6):68-71.
[12] 劉華友,朱 玲,李 帆,等.斑馬魚Foxp4基因多克隆抗體的制備及檢測[J].激光生物學(xué)報(bào), 2011,20(4):556-559.
[13] 梁 艷,宋懷婷,李競超,等.myh6蛋白的表達(dá)、純化及多克隆抗體制備[J].激光生物學(xué)報(bào), 2010,19(5):63-67.
猜你喜歡
中學(xué)生數(shù)理化·中考版(2022年8期)2022-06-14 06:55:24
數(shù)學(xué)年刊A輯(中文版)(2022年4期)2022-02-16 08:17:34
今日農(nóng)業(yè)(2021年19期)2022-01-12 06:16:36
中老年保健(2021年11期)2021-08-22 03:15:44
無線電通信技術(shù)(2021年4期)2021-07-13 08:58:28
無線電通信技術(shù)(2021年3期)2021-06-08 03:33:48
中學(xué)生數(shù)理化(高中版.高考數(shù)學(xué))(2021年1期)2021-03-19 08:28:38
無線電工程(2020年11期)2020-10-29 01:25:46
現(xiàn)代出版(2020年3期)2020-06-20 07:10:34
福利中國(2015年4期)2015-01-03 08:03:38
