核桃優良品種枝條抗寒性生理生化指標分析
2012-12-23 06:25:04宋艷波喬永勝石美娟武彥霞劉朝紅樊新萍
山西農業科學 2012年10期
王 勇,宋艷波,喬永勝,石美娟,武彥霞,劉朝紅,田 鑫,樊新萍,安 棟
(1.山西省農業科學院果樹研究所,山西太谷030815;2.山西農業大學生命科學學院,山西太谷030801)
核桃(Juglans regia L.)屬核桃科落葉喬木[1],在山西分布廣泛,南北均有栽培,尤其以太行、呂梁等丘陵地區分布最多[2]。我國北方地區核桃幼樹普遍存在著不同程度的低溫傷害,主要包括冬季低溫凍害和早春霜害[3]。核桃幼樹遭受低溫凍害后,部分主枝、多年生大枝及1 年生枝會死亡,當年大量減產,甚至絕收。國內外許多學者已在果樹抗寒機理、抗寒性鑒定、抗寒資源評價及抗寒性遺傳育種等領域進行了大量研究,并取得了良好的進展[4-9]。王勇等[10]對核桃枝條電解質滲出率與核桃樹的抗寒性關系進行了研究,結果表明,品種間和不同處理溫度間的電解質滲出率均存在極顯著差異,核桃枝條電解質滲出率隨處理溫度的降低而逐漸升高;不同樹齡間的電解質滲出率存在顯著差異,核桃成齡樹1 年生枝條抗寒性略高于幼齡樹。王勇等[10]還對核桃的1 年生枝條的抗寒性指標進行了測定分析,結果表明,元豐、遼核4 號、遼核1 號比阿7、扎343 品種比較抗寒;對抗寒性指標進行分析比較,品種間抗寒性差異較大時可用電導率法測定,此法既簡便又節省費用;抗寒性差異小時可用總酚、黃酮類物質、可溶性糖和淀粉含量進行抗寒性綜合鑒定。
本試驗在前人研究基礎上[11-16],以禮品1 號、禮品2 號、中林1 號、中林5 號、遼核1 號、遼核3號和晉龍1 號7 個山西主推的核桃品種為試材,對其生育后期和休眠期的新梢和1 年生枝條的抗寒性生理生化指標,包括可溶性糖、可溶性蛋白、脯氨酸含量及超氧化物歧化酶(SOD)、過氧化物酶(POD)活性等進行了分析測定,旨在明確各指標的變化規律,分析其與核桃抗寒性之間的關系,并對各個品種的抗寒性進行比較,為核桃品種的選擇及合理栽培管理提供理論依據。
1 材料和方法
1.1 試驗材料
試驗在山西省農業科學院果樹研究所進行,供試品種為禮品1 號、禮品2 號、中林1 號、中林5 號、遼核1 號、遼核3 號和晉龍1 號,樹齡5 年生,每個品種選取生長狀況基本相同的植株6 株,2010 年9 月底至2011 年2 月底,每隔1 個月采樣一次,從每株樹樹體中上部四周隨機采取整齊均勻、粗細相近的新梢和1 年生枝條1 個組成混合樣,用保鮮袋裝好帶回實驗室。用自來水和去離子水沖洗多次,然后用潔凈的紗布擦干,最后用保鮮袋分裝備用。每個處理3 次重復。
主要試劑有:磷酸、愈創木酚、甲基氨酸(met)、氮藍四唑(nbt)、核黃素、EDTA、考馬斯亮藍G250、蒽酮- 乙酸乙酯、90%的乙醇、茚三酮、冰醋酸、磺基水楊酸。試驗器材有:UV-2450 分光光度計、PRZMOR 多功能離心機、BT 244S 電子天平。
1.2 試驗方法
1.2.1 可溶性糖的測定 參照喬富廉[17]的方法測定各樣品可溶性糖含量。
1.2.2 可溶性蛋白含量測定 采用考馬斯亮藍G250 法測定各樣品可溶性蛋白含量[18]。
1.2.3 脯氨酸含量測定 參照喬富廉[17]的方法測定各樣品脯氨酸含量。
1.2.4 酶活性的測定 參照李合生等[19]的方法測定各樣品酶活性。
2 結果與分析
2.1 可溶性糖含量的變化
從圖1 可以看出,隨著核桃生育進程的繼續以及氣溫的變化,除中林5 號和遼核3 號核桃枝條的可溶性糖含量一直呈上升趨勢外,其余5 個品種枝條的可溶性糖含量整體呈先升高后降低的明顯變化。在12 月多數品種可溶性糖含量大幅增加,在最低溫之前均達到最大,這是由于初期的低溫鍛煉,激發了植物體內可溶性糖的積累,以此來提高植物的抗寒性。之后除中林5 號、遼核3 號枝條的可溶性糖含量繼續上升外,其余5 個品種均開始出現小幅下降趨勢,可能是由于這5 個品種正在逐步解除自然休眠,而中林5號、遼核3 號可能仍處在自然休眠狀態。

不同品種間可溶性糖含量比較如表1 所示。
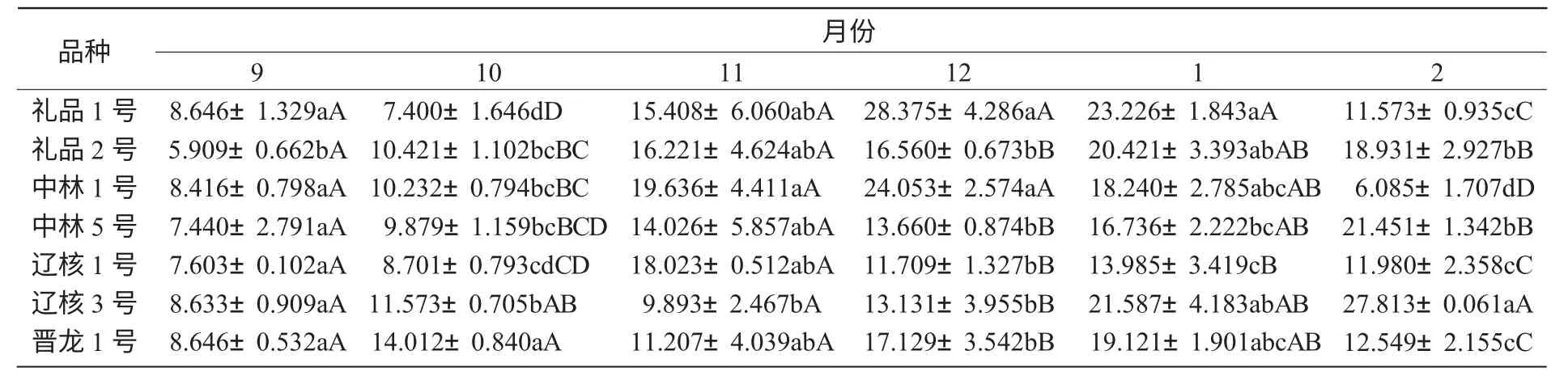
表1 不同品種間可溶性糖含量比較μg/g
從表1 可以看出,各品種在越冬過程中枝條可溶性糖含量存在明顯差異,依其枝條可溶性糖含量的高低,推斷其抗寒性強弱順序依次為:遼核3 號>禮品1 號>禮品2 號>中林1 號>中林5 號>晉龍1 號>遼核1 號。
2.2 可溶性蛋白含量的變化
植物體內可溶性蛋白含量是一個重要的生理生化指標,當遭受低溫脅迫時,體內蛋白質含量就會有所積累,以調節植物抗寒作用。
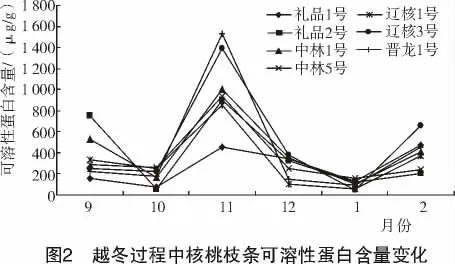
由圖2 可知,在整個越冬過程中供試核桃枝條的可溶性蛋白含量變化比較一致,呈降—升—降—升的“W”趨勢。隨著氣溫下降,7 個品種可溶性蛋白質含量大幅增加,到11 月上旬,可溶性蛋白質含量最高。從整體上看,以晉龍1 號增加幅度最大,含量最高,可見其對寒冷的調節作用最強;禮品1 號增幅最小,對寒冷的調節作用最弱。
從表2 可以看出,各品種在越冬過程中枝條可溶性蛋白含量存在差異,遼核3 號和禮品1 號之間的可溶性蛋白含量差異顯著,其他各品種間差異不顯著。依各品種枝條可溶性蛋白含量的高低,推斷其抗寒性強弱順序依次為:遼核3 號>晉龍1 號>中林1 號>禮品2 號>中林5 號>遼核1 號>禮品1 號。
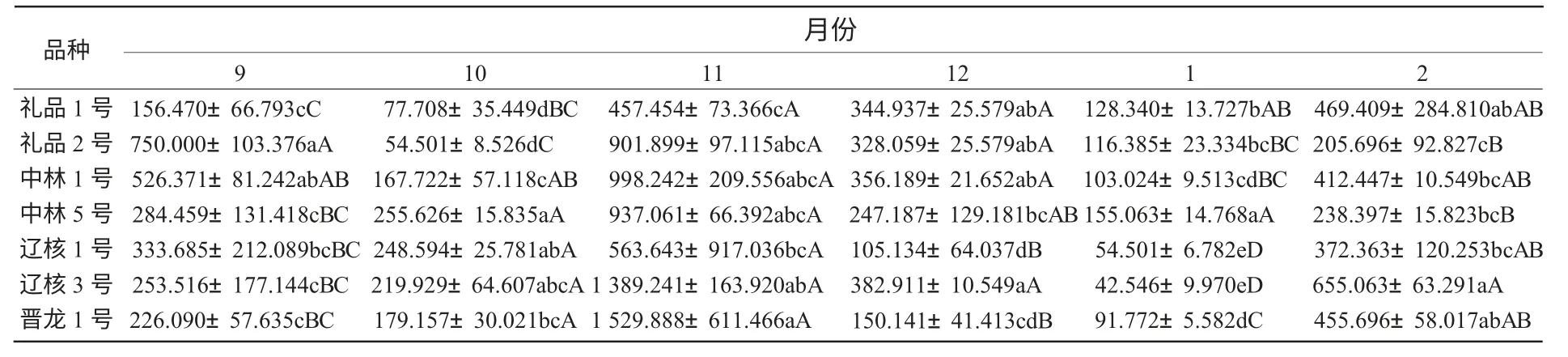
表2 不同品種間可溶性蛋白含量比較μg/g
2.3 脯氨酸含量的變化
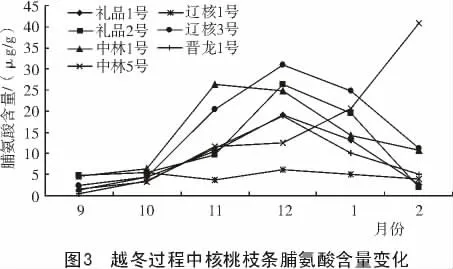
由圖3 可知,2010 年9—12 月,溫度逐漸降低的過程中,各品種脯氨酸含量在逐漸升高。有6 個核桃品種在越冬過程中枝條脯氨酸含量基本呈先升高后降低的狀態,而中林5 號枝條中脯氨酸含量一直呈升高的狀態。但是在整個過程中,脯氨酸的平均含量仍然是遼核3 號最高,遼核1 號最低。然后隨著核桃生育進程的繼續以及氣溫的變化,供試品種除中林5 號的脯氨酸含量繼續上升外,其他品種都有不同幅度的下降。這可能是由于中林5 號仍然處于自然休眠狀態,而其他品種已經解除自然休眠的原因。
由表3 可知,遼核3 號與禮品1 號、晉龍1 號、遼核1 號間差異顯著;遼核3 號、中林5 號與遼核1 號間差異極顯著;其他各品種間差異不顯著。依7 個品種枝條脯氨酸含量的高低,推斷其抗寒性順序依次為:遼核3 號>中林5 號>中林1 號>禮品2 號>禮品1 號>晉龍1 號>遼核1 號。
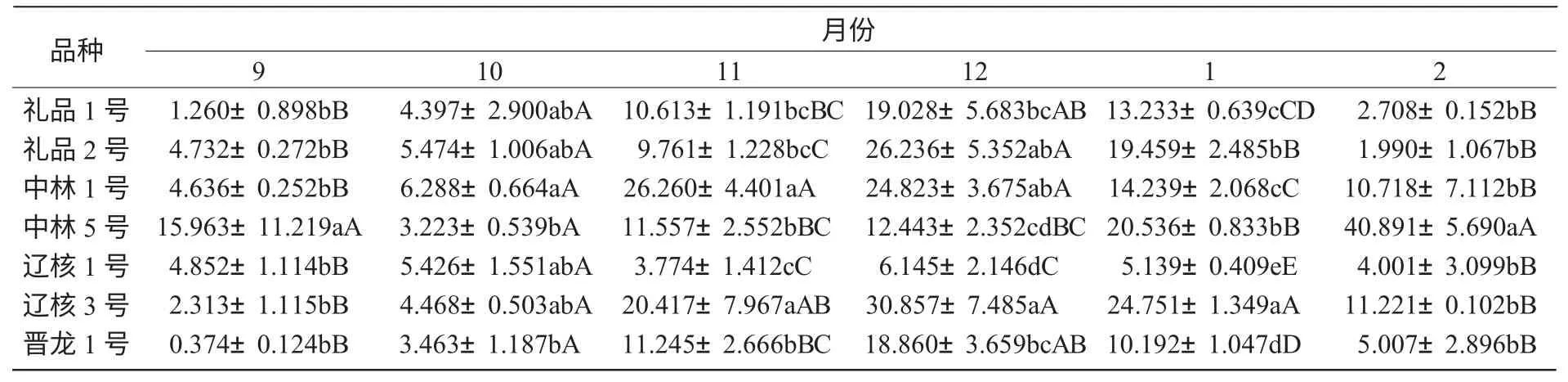
表3 不同品種間脯氨酸含量比較μg/g
2.4 保護酶活性的變化
2.4.1 越冬過程中POD 酶活性變化 由圖4 可知,各個核桃品種枝條POD 酶活性隨著越冬過程中氣溫變化及生理發育過程,總體呈初期有小幅下降然后逐漸走高的趨勢。在溫度較低時,各品種枝條所含的POD 酶活性的變化幅度不大,但隨著溫度的升高,POD 酶的活性迅速升高。而且到測定后期,各個品種的POD 酶活性都有較大幅度增加。這可能是各個品種在測定后期都已經解除了自然休眠,導致POD 酶活性的大幅變化。氣溫最低時,中林1 號、禮品2 號、禮品1 號的POD 酶活性較高,而遼核3 號的POD 酶活性相對較低。不同品種間POD 酶活性如表4 所示。
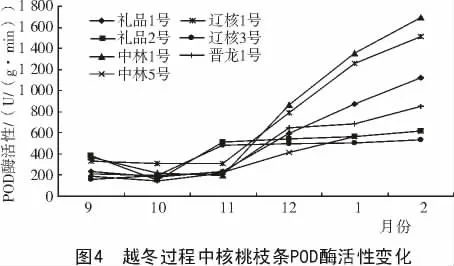
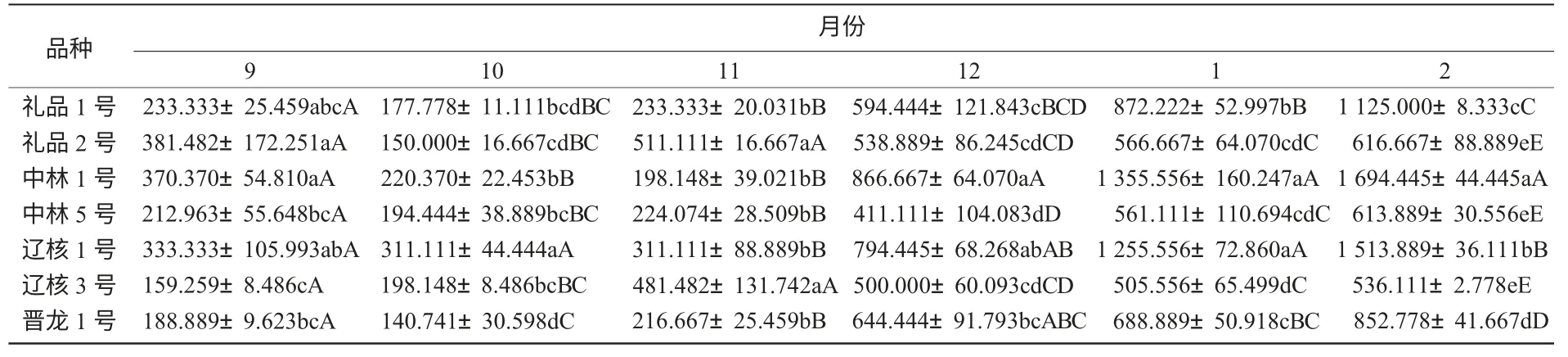
表4 不同品種間POD 酶活性比較U/(g·min)
由表4 可知,各品種枝條POD 酶活性存在顯著差異,中林1 號與禮品2 號、晉龍1 號、遼核3 號、中林5 號之間差異極顯著。依其枝條POD酶活性的高低,推斷其抗寒性強弱順序依次為:中林1 號>遼核3 號>禮品1 號>禮品2 號>晉龍1 號>遼核1 號>中林5 號。
2.4.2 越冬過程中SOD 酶活性變化 從圖5 可以看出,禮品1 號、禮品2 號、中林1 號、遼核3 號、晉龍1 號在整個越冬過程中呈降—升—降的變化趨勢,而中林5 號和遼核1 號呈升—降—升—降的變化趨勢。在降溫初期,各品種SOD 酶活性均有小幅下降,經過初期的低溫鍛煉后,進入12 月底時,各品種又有不同程度的升高,其中以遼核3 號的活性最大。到1 月份隨著氣溫回升,SOD 酶活性表現為降低的趨勢。在整個越冬過程中,遼核3 號的SOD 活性平均值最高,為243.55 U/(g·min)。
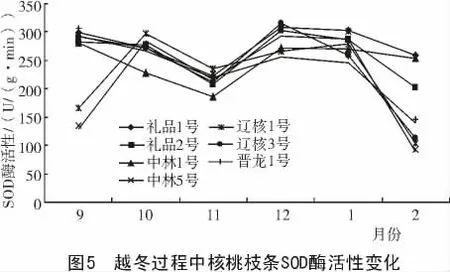
從表5 可以看出,各品種枝條SOD 酶活性存在差異,依其枝條SOD 酶活性的高低,推斷其抗寒性強弱順序依次為:禮品2 號>晉龍1 號>禮品1 號>中林1 號>遼核3 號>中林5 號>遼核1 號。
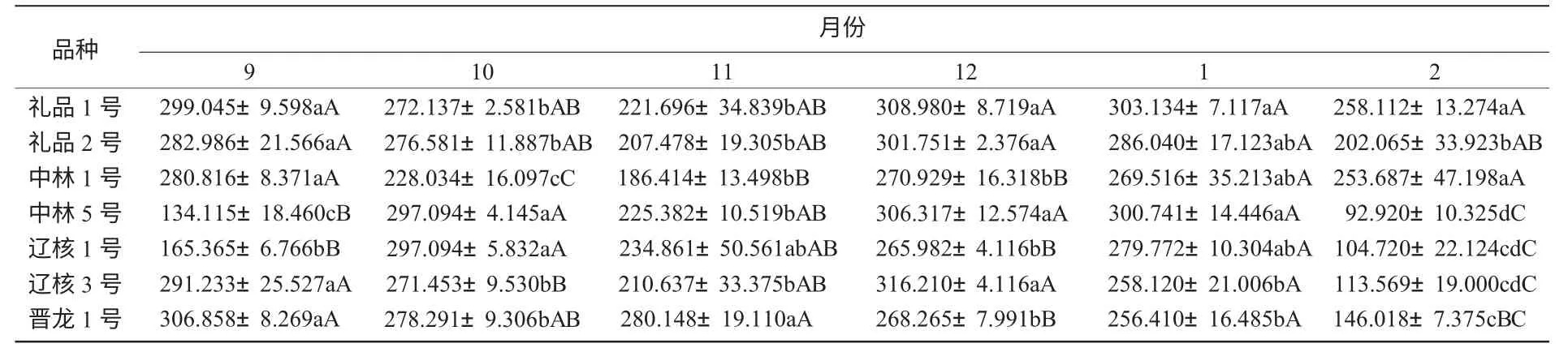
表5 不同品種間SOD 酶活性比較U/(g·min)
3 結論
本試驗結果表明,隨著核桃生育進程的繼續以及氣溫的變化,可溶性糖、可溶性蛋白、脯氨酸含量、POD 酶活性在核桃枝條內含量越高,抗寒性越強,而SOD 酶活性越高,其抗寒性越弱。
綜合比較各品種核桃枝條抗寒性生理生化指標,依各指標與抗寒性可能的相關性,推測7 個品種的抗寒性強弱可能為:遼核3 號、晉龍1號抗寒性較強,中林1 號、禮品2 號和禮品1 號的抗寒性次之,而中林5 號和遼核1 號的抗寒性較差。
由于條件和時間的限制,本試驗只對山西省農業科學院果樹研究所核桃園7 個品種的部分核桃枝條抗寒性生理生化指標進行了測定分析,而核桃生產實踐中栽培應用的品種和影響果樹抗寒性的因素及相關指標非常多,還需進行更深入的研究。
[1]潘瑞熾,董愚得.植物生理學[M].北京:高等教育出版社,1995.
[2]Kataoka,K Sumitomo,T Fudano,et al.Changes in sugar content of Phalaenopsis leaves before floral transition[J].Scientia Horticulturae,2004,102:121-132.
[3]劉祖祺,張石誠.植物抗性生理學[M].北京:中國農業出版社,1994.
[4]陳新華,郭寶林,趙靜,等.休眠期內甜櫻桃不同品種枝條的抗寒性[J].河北農業大學學報,2009,3(6):37-40.
[5]胡春霞,王麗,湯杰.低溫對南果梨的生理生化指標的影響[J].沈陽農業大學學報,2009,40(3):349-352.
[6]周蘊薇.翠南報春抗寒生理生態學研究[D].儋州:華南熱帶農業大學,2003.
[7]黃義江,王宗清.蘋果屬果樹抗寒性的細胞學鑒定[J].園藝學報,1982,9(3):23-30.
[8]鄒喻蘋.葡萄抗寒生理特性的分析[J].植物學集刊,1989,1(1):64-68.
[9]周碧燕,梁立峰,黃輝白,等.低溫和多效唑對香蕉及大蕉超氧化物歧化酶和脫落酸的影響[J].園藝學報,1995,22(4):331-335.
[10]王勇,田建保,程恩明,等.核桃枝條電解質滲出率與抗寒性的關系[J].江西農業大學報,2007(2):230-233.
[11]王麗雪,李榮富,張富仁,等.葡萄枝條中蛋白質、過氧化物酶活性變化與抗寒性的關系[J].內蒙古農牧學院學報,1996,17(1):45-50.
[12]石雪暉,陳祖玉,楊會卿.低溫脅迫對柑橘離體葉片中SOD及其同工酶活性的影響 [J]. 園藝學報,1996,23(4):384-386.
[13]王華,王飛,陳登文,等.低溫脅迫對杏花SOD 活性和膜脂過氧化的影響[J].果樹科學,2000,17(3):197-201.
[14]吳經柔.應用過氧化物酶同Ⅰ酶譜測定蘋果的抗寒性[J].果樹科學,1990,7(1):41-44.
[15]沙廣利,郭長城,睢薇,等.梨抗寒性遺傳的研究[J].果樹科學,1996,13(3):167-170.
[16]金萬梅,董靜,尹淑萍,等.冷誘導轉錄因子CBF1 轉化草莓及其抗寒性鑒定 [J]. 西北植物學報,2007,27(2):223-227.
[17]喬富廉.植物生理學實驗分析測定技術[M].北京:高等教育出版社,2000.
[18]鄒崎.植物生理學實驗指導 [M].北京:中國農業出版社,2000.
[19]李合生,孫群,趙世杰.植物生理生化實驗原理和技術[M].北京:高等教育出版社,2000.
