活性SET7/9融合蛋白的表達與純化
2012-12-25 01:43:38高麗麗余衛平
東南大學學報(醫學版) 2012年5期
關鍵詞:融合
高麗麗,余衛平
(東南大學醫學院病理學與病理生理學系,江蘇南京 210009)
SET7/9含有SET結構域,是蛋白賴氨酸甲基化轉移酶 (proteinlysinemethyltransferases,PLMTs或PKMTs)家族成員。有報道稱之為SET7,也有報道稱之為SET9[1-2]。近年來發現SET7/9催化的非組蛋白甲基化作用能引起蛋白穩定性改變和基因表達變化等多種分子效應,涉及到染色體結構、細胞周期及凋亡等方面。因研究需要,我們建立了活性GST-SET7/9融合蛋白(SET7/9融合蛋白)表達系統與純化方法。
1 材料與方法
1.1 菌株和載體
本實驗所需的大腸桿菌CM1061菌株、大腸桿菌BL21(DE3)表達菌株、pCMV-SET7/9質粒均由本課題組保存;原核表達載體質粒pGEX-6P-3由Amersham Biosciences公司提供。
1.2 主要試劑
PCR產物純化、質粒提取試劑盒為QIAGEN公司產品;PCR酶、限制性內切酶購自 TaKaRa公司;T4 DNA連接酶由New England Biolabs公司提供;Glutathione Agarose吸附柱為Sigma公司產品;人組蛋白H3由 upstate公司提供;Adenosel-L-Methionine,S-K[methyl-3H]由PerkinElmer公司提供;Bio-Rad蛋白測定試劑盒由BIO-RAD公司提供;引物合成、測序均在INVITROGEN公司完成。
1.3 pGEX-SET7/9原核重組表達載體的構建
1.3.1 設計與合成寡聚核苷酸引物 目的基因為含SET結構區的SET7/9基因片段,位于pCMV-SET7/9全基因序列的第268~1 365個核苷酸。通過NCBI GenBank確定pCMV-SET7/9的cDNA序列,選擇EcoRⅠ和XhoⅠ作為酶切位點,采用DNA Star軟件設計SET7/9引物。SET7/9引物上游:5'-ACCGAATTCCAT GGATAGCGACGAC-3',含EcoRⅠ酶切位點及啟始密碼子;SET7/9引物下游:5'-CTCCTCGAGTCATTACTTT TGCTGGGTGGCCTG-3',含終止密碼子及XhoⅠ酶切位點。
1.3.2 擴增目的基因 以pCMV-SET7/9質粒為模板,PCR擴增目的基因。等量去離子水代替模板作為陰性對照。其反應條件為94℃ 13 min熱啟動;94℃1 min,60 ℃ 1 min,72 ℃ 2 min,35個循環;最后72 ℃延伸10 min。PCR產物經1%瓊脂糖凝膠電泳后,純化回收,產物純化后測序確認。
1.3.3 構建pGEX-SET7/9表達載體 將原核表達載體pGEX-6P-3轉入大腸桿菌CM1061菌株。挑取單個菌落培養,大量提取質粒DNA。將純化后的PCR產物和pGEX-6P-3質粒分別用核酸內切酶EcoRⅠ與XhoⅠ進行酶切,電泳鑒定并用PCR產物純化試劑盒純化,氯仿抽提法回收。用T4 DNA連接酶連接含SET7/9的基因片段與表達載體,獲得pGEX-SET7/9重組質粒。
接種轉化有重組質粒的大腸桿菌BL21(DE3)單菌落至10 ml含氨芐青霉素抗性的LB液體培養基中,37℃振蕩培養過夜。次日按1∶100的比例轉接過夜培養菌至15 ml含氨芐青霉素液體培養基中,37℃繼續振蕩培養至OD600nm為0.6左右,均分該培養物為兩部分:一部分加入異丙基β-D硫代半乳糖苷(IPTG)至終濃度為1 mmol·L-1以誘導SET7/9融合蛋白表達,在誘導培養的不同時間(0、1、2、3、4 h)收取菌液,各取1 ml培養物離心后留沉淀置-70℃保存待用;另一部分不加IPTG,相同方法收取不同時間段的細菌沉淀作對照。將收集到的細菌菌體用十二烷基硫酸鈉聚丙烯酰胺凝膠電泳(SDS PAGE)法,檢測與鑒定含pGEXSET7/9質粒的陽性菌落。
1.4 SET7/9融合蛋白的表達與提純
挑取單一陽性菌落接種在10 ml含青霉素抗性的LB液體培養基中,37℃振蕩培養過夜,次日按1∶100的比例轉接于500 ml含青霉素抗性的培養基中,于37℃振蕩培養至OD600nm為0.6左右,加入IPTG至終濃度為1 mmol·L-1以誘導GST-SET7/9表達,繼續培養5.0 h。離心收取菌體用5 ml PBS重新懸浮細胞于4℃超聲破碎以裂解細菌,利用Glutathione Agarose吸附柱提純SET7/9融合蛋白,產物進行SDS-PAGE電泳檢測,確定目的蛋白所在部位。
1.5 SET7/9融合蛋白的組蛋白甲基化作用分析
采用Bio-Rad蛋白測定試劑盒檢測純化的SET7/9融合蛋白濃度,組蛋白甲基化轉移酶(HMT)測定法分析蛋白活性。通過LS6000IC型液閃儀(BECKMAN公司)測得的每分鐘脈沖數(CPM)反映蛋白質HMT活性。
2 結 果
2.1 pGEX-SET7/9重組質粒的PCR鑒定
pGEX-SET7/9重組質粒通過PCR擴增,其目的基因長度為1 123 bp,與分子質量參照物(DNA Ladder)相比,PCR擴增產物的大小與預測結果一致(圖1)。
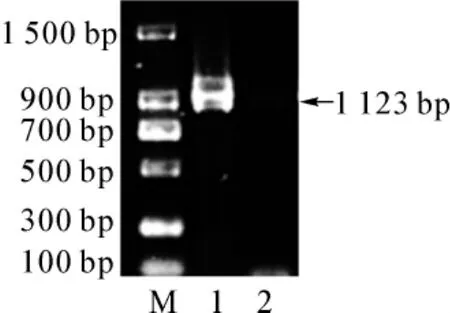
圖1 PCR擴增SET7/9結果(1%瓊脂糖凝膠電泳)Fig 1 Amplification of SET7/9 with PCR(1%agarose gel)
2.2 SET7/9融合蛋白的表達與純化
SET7/9融合蛋白進行SDS-PAGE蛋白電泳,經考馬斯亮藍R-250染色發現,含重組質粒的誘導菌在分子質量大約77 kD位置出現一蛋白條帶。已知目的基因表達的多肽分子質量為50 kD,與融合蛋白共同表達分子質量約為77 kD,故獲得的純化蛋白分子質量與理論估計值相一致(圖2)。
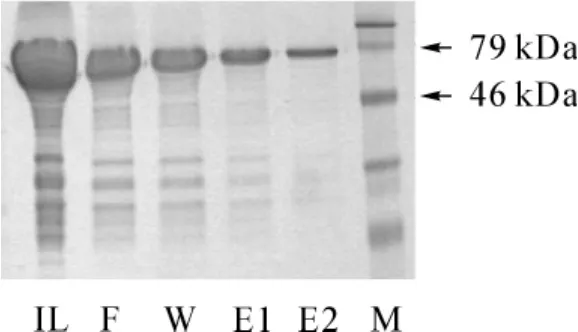
圖2 提純SET7/9融合蛋白的SDS-PAGE結果Fig 2 SDS-PAGE showing the purification of SET7/9 protein
2.3 SET7/9融合蛋白的組蛋白甲基化作用
采用HMT測定法可見,孵育30、60、120 min后的樣本,純化SET7/9融合蛋白組CPM值均明顯高于對照組(GST蛋白質組),與對照組相比分別增加21.7%、19.8%、22.1%,且與作用時間呈正相關,表明融合蛋白SET部分具有HMT活性(表1)。

表1 純化SET7/9蛋白的組蛋白甲基化作用分析Tab 1 Histone methylation analysis of the purified SET7/9 protein
3 討 論
SET7/9蛋白分子質量大約為50 kD,其表達基因位于第4號染色體4q28。SET7/9所含的SET結構域具有由β折疊環繞成的類似繩結的三級結構[3-5]。最初發現SET7/9是靶向組蛋白H3第4位賴氨酸(H3K4)的甲基化轉移酶,但進一步研究發現SET7/9只對游離的H3K4而非核小體H3K4有高度活性[1-2,4]。此外,SET7/9 還能使膜相關受體、轉錄因子、腫瘤抑制因子等蛋白賴氨酸甲基化,從而調節這些非組蛋白的功能[6-7]。
SET7/9對非組蛋白賴氨酸的甲基化作用會產生復雜的生物學效應,如甲基化p53的K372位點可增加p53穩定性及其對靶基因的活化作用;甲基化衍生因子(elongation factor,E2F1)的K185位點可抑制該蛋白的乙酰化和磷酸化,并激活蛋白泛素化,抑制凋亡;還可使成視網膜母細胞瘤蛋白(retinoblastoma protein,pRb)、雌激素受體 α (estrogen receptor α,ERα)等蛋白甲基化,影響它們的生物學效應[6-7]。可見SET7/9催化的非組蛋白賴氨酸甲基化作用可能與腫瘤等疾病的發病機制相關[6-7]。
為了開展SET7/9對非組蛋白甲基化修飾及其效應的研究,我們需要創建SET7/9蛋白表達系統。本項工作所構建的pGEX-SET7/9重組質粒在原核細胞中能夠表達SET7/9融合蛋白。純化的SET7/9融合蛋白通過HMT活性測定有明顯的組蛋白甲基化作用,表明pGEX-SET7/9能夠表達高活性的SET7/9融合蛋白[8]。這會為我們后續相關工作提供幫助。
[1]WANG H B,CAO R,XIA L,et al.Purification and functional characterization of a histone H3-lysine 4-specific methyltransferase[J].Mol Cell,2001,8(6):1207-1217.
[2]NISHIOKA K,CHUIKOV S,SARMA K,et al.Set9,a novel histone H3 methyltransferase that facilitates transcription by precluding histone tail modifications required for heterochromatin formation[J].Genes Dev,2002,16(4):479-489.
[3]QIAN C,ZHOU M M.SET domain protein lysine methyltransferases:structure,specificity and catalysis[J].Cell Mol Life Sci,2006,63(23):2755-2763.
[4]WILSON J R,JING C,WALKER P A,et al.Crystal structure and functional analysis of the histone methyltransferase SET7/9[J].Cell,2002,111(1):105-115.
[5]XIAO B,JING C,WILSON J R,et al.Structure and catalytic mechanism of the human histone methyltransferase SET7/9[J].Nature,2003,421(6923):652-656.
[6]高麗麗,余衛平.PLMT家族成員SET7/9的非組蛋白甲基化作用[J].生命科學,2011,23(8):773-778.
[7] EA C K,BALTIMORE D.Regulation of NF-κB activity through lysine monomethylation of p65[J].Proc Natl Acad Sci USA,2009,106(45):18972-18977.
[8]余衛平,方娟娟,DONG W F,等.含RIZ1 PR結構域融合蛋白的原核表達、提純與功能分析[J].東南大學學報:醫學版,2006,25(4):240-243.
猜你喜歡
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
數學年刊A輯(中文版)(2022年4期)2022-02-16 08:17:34
今日農業(2021年19期)2022-01-12 06:16:36
中老年保健(2021年11期)2021-08-22 03:15:44
無線電通信技術(2021年4期)2021-07-13 08:58:28
無線電通信技術(2021年3期)2021-06-08 03:33:48
中學生數理化(高中版.高考數學)(2021年1期)2021-03-19 08:28:38
無線電工程(2020年11期)2020-10-29 01:25:46
現代出版(2020年3期)2020-06-20 07:10:34
福利中國(2015年4期)2015-01-03 08:03:38
