微小RNA-26b在乳腺癌中的作用及其分子機制研究
2013-04-20 03:47:00徐麗平
中國全科醫學 2013年33期
徐麗平,李 佳,房 林
乳腺癌是女性最常見的惡性腫瘤之一,根據美國2010年發布的數據,2009年全美女性新發乳腺癌207 090例,高居榜首,遠超過第2位消化系統腫瘤125 790例,乳腺癌死亡共39 840例[1]。2003—2007年我國乳腺癌發病率41.64/10萬,居女性癌癥發病的第1位,死亡率 9.63/10萬,居女性腫瘤死亡率的第6位[2]。因此乳腺癌已成為嚴重威脅全世界女性生命的疾病之一。研究表明微小RNA(microRNAs)是一組非編碼RNA,通過作用于mRNA使其降解或抑制翻譯,調控基因的表達,從而參與細胞的增殖、凋亡、細胞分化、腫瘤的侵襲轉移以及腫瘤干細胞調控等[3-6]。微小RNA-26b(miR-26b)在肝細胞癌、肺癌、乳腺癌和膀胱癌等多種腫瘤中表達異常,其在不同腫瘤中扮演著癌基因或者抑癌基因的角色[7-11]。本研究采用熒光定量反轉錄聚合酶鏈式反應(RT-PCR)檢測miR-26b在乳腺癌及正常乳腺組織中表達水平,MTT法分析其對于乳腺癌MBA-MD-231細胞增殖的影響,并尋找其可能靶基因。
1 資料與方法
1.1 一般資料 選取上海市同濟大學附屬第十人民醫院甲乳外科2010年1月—2012年5月術后病理確診為乳腺癌的女性患者38例,年齡35~82歲,中位年齡52歲;術前均未行化療、放療及內分泌治療。取乳腺癌組織,并取距離腫瘤組織>5 cm正常乳腺組織作為對照。隨訪時間為行手術治療后至2013年4月,患者術后均接受化療,目前均無復發轉移。人乳腺癌MBA-MD-231細胞株購于中國科學院。
1.2 microRNAs提取和反轉錄 按照Tiangen公司miRcute microRNA提取分離試劑盒說明書進行操作。microRNAs反轉錄參照Chen等[12]的方法,通過特異莖環引物,按照下列順序在PCR試管中加入反應物(體積為8.6 μl):超純水 2 μl,5×buffer 1.7 μl,microRNAs莖環引物(各個microRNAs及內參U6)1.9 μl,反轉錄酶(Primescript RT Enzyme MixI) 1 μl,microRNAs sample 2 μl。反應條件為:16 ℃ 30 min,42 ℃ 30 min,85 ℃ 8 min,4 ℃ 10 min,反應結束后置于4 ℃冰箱保存。
1.3 實時定量RT-PCR 按照Takara公司SYBR Premix Ex Taq(Perfect Real Time) DRR041A說明書完成加樣,反應條件為95 ℃預變性2 min;變性95 ℃ 30 s,退火、延伸60 ℃ 45 s,重復35個循環,70~95 ℃繪制熔解曲線。反應結束后,RQ Manager分析軟件統計處理,獲取Ct值,并根據熔解曲線判斷產物的純度,結果分析根據Livak等[13]的描述,采用半定量分析。
1.4 細胞增殖檢測 按照每孔5 000個細胞的密度將乳腺癌MBA-MD-231細胞種入96孔板。將接種好的96孔板細胞放入培養箱中培養,至細胞單層密度達到50%~60%。分組:采用50、100 nmol/L miR-26b mimics轉染為50 nmol/L組和100 nmol/L組,參照Invitrogen公司lipofectamine說明書完成96孔板細胞轉染;脂質體處理為空對照組,小分子RNA片段轉染組為負對照組。按轉染后24、48、72、96 h 4個時間段分別進行如下操作:每孔加入20 μl MTT溶液(5 mg/L),繼續培養4 h。吸取上清棄掉,每孔加入150 μl二甲基亞砜(DMSO),置搖床上低速震蕩10 min,使結晶物充分溶解。在酶聯免疫檢測儀490 nm處測量各孔的吸光值(OD)。
1.5 熒光素酶報告實驗 通過TargetScan軟件尋找miR-26b可能靶基因,前列腺素內過氧化物酶2(PTGS-2)3′UTR存在一個潛在miR-26b靶位結合點(257-264)。以人乳腺癌MBA-MD-231細胞cDNA為模板,按照Primer star kit(Takara)說明書PCR擴增包括miR-26b結合靶點在內的PTGS-2基因部分3′UTR區。PCR引物:正義引物5′-TAGGCGATCGCTCGAGCTGTTGCGGAGAAAGGAGTC-3′,反義引物 5′-AATTCCCGGGCTCGAGTAGTTACTTCTAATGCATCATGG-3′。將目的片段亞克隆到psiCHECK-2質粒上,酶切鑒定和測序分析其正確性。在96孔板中培養人乳腺癌MBA-MD-231細胞,當細胞密度為60%左右時進行轉染,分別轉染psiCHECK-2 PTGS-2 3′-UTR 和100 nmol/L miR-26b mimics(PTGS-2組),psiCHECK-2/PTGS-2 3′-UTR 突變體 (200 ng) 和 miR-26b (100 nmol/L)(突變體對照組),200 ng psiCHECK-2空載體和100 nmol/L miR-NC (空載體對照組),每組設5個復孔。轉染后繼續培養48 h。根據Dual-luciferase report assay system (Promega,USA)說明書完成熒光素酶報告實驗。
1.6 Western blotting 實驗組為100 nmol/L miR-26b mimics轉染組,空對照組為脂質體處理組,負對照組為小分子RNA片段轉染組。轉染后48 h分別收集實驗組和對照組細胞,用預冷的RIPA裂解液裂解后取上清,以Bradford法蛋白定量,10%十二烷基硫酸鈉聚丙烯酰胺凝膠(SDS-PAGE)電泳,然后電轉移至硝酸纖維素(NC)膜上,分別加入1∶1 000稀釋的PTGS-2一抗(BioVision,USA),4 ℃孵育過夜,1∶1 000稀釋的二抗稀釋液中孵育,磷酸鹽吐溫緩沖液(PBST)洗膜后進行奧德賽系統掃描。
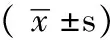
2 結果
2.1 實時定量RT-PCR結果 乳腺癌組織中miR-26b ΔCt為(15.25±0.98),正常乳腺組織中為(12.37±1.23),正常乳腺組織中miR-26b表達水平為乳腺癌組織的7.33倍,乳腺癌組織中miR-26b表達低于正常乳腺組織,差異有統計學意義(t=12.21,P=0.007)。
2.2 MTT實驗結果 MTT實驗結果顯示,50 nmol/L組、100 nmol/L組、空對照組和負對照組轉染后24、48 h相對細胞增殖比較,差異均無統計學意義(F值分別為1.314和1.830,P值分別為0.336和0.220);4組轉染后72、96 h相對細胞增殖比較,差異均有統計學意義(F值分別為11.037和5.470,P值分別為0.003和0.024);其中轉染后96 h 50 nmol/L組與空對照組比較,差異有統計學意義 (q=-3.048,P=0.038);轉染后72 h和96 h 100 nmol/L組與空對照組比較,差異均有統計學意義 (q值分別為-3.248和-4.254,P值分別為0.031和0.013,見圖1)。

圖1 不同組乳腺癌MBA-MD-231細胞相對細胞增殖比較
Figure1 Comparison of the proliferation of MBA-MD-231 cancer cells in different groups
2.3 熒光素酶報告實驗 PTGS-2 mRNA在PTGS-2組中ΔCt為(3.91±0.21),突變體對照組及空載體對照組中分別為(2.25±0.31)、(2.53±0.48)。熒光素酶報告實驗結果顯示,PTGS-2組、突變體對照組及空載體對照組熒光素酶活性比較,差異有統計學意義(F=99.88,P=0.000);其中PTGS-2組較突變體對照組和空載體對照組均降低,差異有統計學意義(q值分別為-10.314和-12.108,P值分別為0.000和0.000,見圖2)。

圖2 不同組熒光素酶活性比較
Figure2 Comparison of luciferase activities of MBA-MD-231 cancer cells in different groups
2.4 Western blotting結果分析 與空對照組及負對照組相比,實驗組作用乳腺癌MBA-MD-231細胞株48 h后,PTGS-2蛋白的表達量明顯減少(見圖3)。

圖3 不同組PTGS-2蛋白表達情況
Figure3 PTGS-2 protein expression of MBA-MD-231 cancer cells in different groups
3 討論
microRNAs是一類進化上高度保守的單鏈RNA,通常長度在22 nt左右,結合于靶基因mRNA的3′-UTR區域,阻遏翻譯或者直接降解mRNA,抑制基因表達。在腫瘤發生和發展中,miRNA表達譜發生改變,抑制腫瘤的microRNAs表達下調,而促進腫瘤的microRNAs過表達。本實驗發現miR-26b在乳腺癌組織和正常乳腺組織中差異表達,在乳腺癌組織中表達明顯下調,這與在肝癌[7]、鼻咽癌[8]、原發性肺鱗狀細胞癌[9]中的報道相一致。目前的研究表明miR-26b在多種腫瘤中具有多元的作用,如高表達的miR-26b可通過作用于COX-2(cyclooxygenase-2)明顯抑制鼻咽上皮細胞癌細胞的增殖[8],相反下調垂體腫瘤細胞中miR-26b的表達水平可抑制克隆形成、腫瘤侵襲和小鼠體內成瘤能力[10]。miR-26b通過抑制SLC7A11表達,促進乳腺癌細胞的凋亡,從而參與凋亡的調控過程[11]。本實驗發現miR-26b能夠抑制MBA-MD-231細胞的增殖,這些研究結果表明miR-26b在多種腫瘤的發生和演進中發揮著重要的作用。
PTGS-2催化花生四烯酸向前列腺素(PGs)和其他類花生酸類物質轉化,其在正常組織中含量極少,缺氧、炎性細胞因子、促癌物、生長因子等可誘發其表達[14-15]。PTGS-2參與致癌、免疫反應抑制、細胞凋亡抑制、血管生成、腫瘤細胞的侵襲和轉移等一系列生理病理過程。最近的研究表明,PTGS-2基因變異的女性好發乳腺癌[16-17],此外,PTGS-2過度表達的乳腺癌患者預后較差[18]。這些研究表明PTGS-2參與腫瘤的形成和發展,其在乳腺癌中扮演著原癌基因的角色。本實驗通過TargetScan預測miR-26b的下游靶基因,發現PTGS-2 3′UTR與miR-26b有著一處潛在的互補結合序列(257-264),這為PTGS-2是miR-26b可能的靶基因提供了理論依據。熒光定量RT-PCR和Western blotting實驗結果顯示100 nmol/L miR-26b mimics抑制PTGS-2 mRNA和蛋白表達水平,且熒光素酶報告實驗證明miR-26b能夠下調PTGS-2組熒光素酶活性,這從實驗基礎上證明PTGS-2即是miR-26b的靶基因,miR-26b通過作用于PTGS-2基因參與乳腺癌細胞的增殖過程,這與Ji等[8]的報道相一致。
傳統的乳腺癌治療策略,主要包括外科手術、放療、化療和內分泌治療等。microRNAs介導的腫瘤分子治療將是根治腫瘤的重要發展方向。miR-26b在乳腺癌中的表達水平降低,能夠抑制細胞增殖,提示其在乳腺癌中扮演著抑癌基因的角色,因此未來可通過上調miR-26b的表達,實現治療乳腺癌的目的。梅玫等[19]研究發現聚酰胺-胺型樹枝狀高分子(PAMAM)微粒可以實現5氟尿嘧啶(5-Fu)和miR-21的同載,聯合應用5-Fu與miR-21抑制劑可有效抑制乳腺癌細胞的體外生長。因此未來可將5-Fu/PAMAM/miR-26b復合物應用于乳腺癌的臨床治療,這為克服腫瘤提供了新的方向。
1 王深明,林穎.關注乳腺癌早期診斷和治療的進展[J].中華實驗外科雜志,2011,28(5):647-649.
2 黃哲宙,陳萬青,吳春曉,等.中國女性乳腺癌的發病和死亡現況——全國32個腫瘤登記點2003—2007年資料分析報告[J].腫瘤,2012,32(6):435-439.
3 Zhang J,Lu Y,Yue X,et al.MiR-124 suppresses growth of human colorectal cancer by inhibiting STAT3[J].PLoS One,2013,8(8):e70300.
4 馮燕,楊暢,劉瓊,等.微小RNA-148b在乳腺癌中的表達及意義[J].中國全科醫學,2012,15(11):3848.
5 Pizzini S,Bisognin A,Mandruzzato S,et al.Impact of microRNAs on regulatory networks and pathways in human colorectal carcinogenesis and development of metastasis[J].BMC Genomics,2013,14(1):589.
6 Yu F,Deng H,Yao H,et al.MiR-30 reduction maintains self-renewal and inhibits apoptosis in breast tumor-initiating cells[J].Oncogene,2010,29(29):4194-4204.
7 Ji J,Shi J,Budhu A,et al.MicroRNA expression,survival,and response to interferon in liver cancer[J].N Engl J Med,2009,361(15):1437-1447.
8 Ji Y,He Y,Liu L,et al.MiRNA-26b regulates the expression of cyclooxygenase-2 in desferrioxamine-treated CNE cells[J].FEBS Lett,2010,584(5):961-967.
9 Gao W,Shen H,Liu L,et al.MiR-21 overexpression in human primary squamous cell lung carcinoma is associated with poor patient prognosis[J].J Cancer Res Clin Oncol,2011,137(4):557-566.
10 Palumbo T,Faucz FR,Azevedo M,et al.Functional screen analysis reveals miR-26b and miR-128 as central regulators of pituitary somatomammotrophic tumor growth through activation of the PTEN-AKT pathway[J].Oncogene,2013,32(13):1651-1659.
11 Liu XX,Li XJ,Zhang B,et al.MicroRNA-26b is underexpressed in human breast cancer and induces cell apoptosis by targeting SLC7A11[J].FEBS Lett,2011,585(9):1363-1367.
12 Chen C,Ridzon DA,Broomer AJ,et al.Real-time quantification of microRNAs by stem-loop RT-PCR[J].Nucleic Acids Res,2005,33(20):e179.
13 Livak KJ,Schmittgen TD.Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method[J].Methods,2001,25(4):402-408.
14 Schmedtje JF Jr,Ji YS,Liu WL,et al.Hypoxia induces cyclooxygenase-2 via the NF-kappa B p65 transcription factor in human vascular endothelial cells[J].J Biol Chem,1997,272(1):601-608.
15 Liu XH,Kirschenbaum A,Yao S,et al.Upregulation of vascular endothelial growth factor by cobalt chloride-simulated hypoxia is mediated by persistent induction of cyclooxygenase-2 in a metastatic human prostate cancer cell line[J].Clin Exp Metastasis,1999,17(8):687-694.
16 Dossus L,Kaaks R,Canzian F,et al.PTGS2 and IL6 genetic variation and risk of breast and prostate cancer:results from the Breast and Prostate Cancer Cohort Consortium (BPC3)[J].Carcinogenesis,2010,31(3):455-461.
17 Langsenlehner U,Yazdani-Biuki B,Eder T,et al.The cyclooxygenase-2 (PTGS2) 8473T>C polymorphism is associated with breast cancer risk[J].Clin Cancer Res,2006,12(4):1392-1394.
18 Festa-Vasconcellos JS,Piranda DN,Amaral LM,et al.Polymorphisms in cycloxygenase-2 gene and breast cancer prognosis:association between PTGS2 haplotypes and histopathological features[J].Breast Cancer Res Treat,2012,132(1):251-258.
19 梅玫,任玉,周旋,等.聚酰胺-氨納米微粒介導5-氟尿嘧啶聯合小RNA-21抑制乳腺癌細胞生長的研究[J].中華實驗外科雜志,2010,27(6):691-693.
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中老年保健(2022年6期)2022-08-19 01:41:48
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
中國生殖健康(2019年2期)2019-08-23 08:11:42
中國生殖健康(2019年6期)2019-01-06 09:20:12
中國生殖健康(2019年5期)2019-01-06 09:16:40
祝您健康(2018年5期)2018-05-16 17:10:16
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
