棉花枯萎病菌產角質酶的連續誘導
2013-04-23 11:52:22冉琴琴張效寧張文坤張學俊
中國釀造 2013年8期
冉琴琴,張效寧,張文坤,張學俊,2*
(1.貴州大學 貴州省發酵工程與生物制藥重點實驗室,貴州 貴陽 550003;2.貴州大學 化學與化學工程學院,貴州 貴陽 550003)
角質酶(Cutinase,EC3.1.1.74)是一種多功能酶,具有屬于α/β水解酶折疊的共同結構的蛋白質[1],是絲氨酸水解酶家族中較小的成員[2],可以降解角質產生脂肪酸[3],也可以水解甘油三酯等化合物[4]。角質酶能夠水解短鏈或長鏈酯類,角質酶除了能參與水解反應,也能參與酯化反應[5],能夠催化酸與醇的酯化、脂肪酸鹽與醇的轉酯化等反應,所以角質酶在生物催化[6]、農業化學品工業[7]、農產品深加工[8]、護理用品行業[9]、生物降解及廢水處理[10]、紡織工業[11]等方面有著廣泛的應用,重要的是其還可以用作酯轉換制備生物柴油。現在,對產角質酶的菌株的研究引起了廣大科學工作者廣泛的興趣,研究主要有產酶菌株篩選、角質酶菌株的誘導和誘變過程表達、發酵條件優化、角質酶結構和酶特性、基因重組改良過表達等方面的內容[12]。
人們是在研究植物病菌治病機理時發現角質酶的,角質層生長在植物表面對植物組織起到保護作用,當致病菌在入侵植物時,必須分泌出的角質酶降解角質層,破壞角質層后方可侵入到植物內部利用植物組織營養進一步繁殖。在自然界中,角質酶的主要來源有2個方面:微生物和花粉[13]。目前,國內外眾多學者對角質酶的研究,主要集中在真菌角質酶和細菌角質酶。
角質酶是誘導酶,只有在角質存在條件下激活真菌內的角質酶基因,誘導其轉錄表達角質酶。DAVIES KA等[14]的研究指出,未經過角質誘導的真菌,其基因轉錄表達的角質酶量非常少,但加入角質時會誘導真菌的角質酶基因,使角質酶轉錄量大大增加。另一方面,分子生物學的方法也可以提高真菌產對角質酶的過表達,張芙華等[15]報道的重組Bacillus subtilisWSHB06-07菌株的角質酶產量可達168.8U/mL。誘導法和分子生物學方法相比之下,分子生物學方法投資高很多、周期長、風險大,而誘導法同樣可獲得高表達角質酶的菌株,方法更為簡便直接,低成本高效率。
本實驗以植物病菌為對象,采用誘導法獲得高產角質酶,因為該類病菌的繁殖必須借助于角質酶。棉花枯萎病菌(Fusarium oxysporum)是常見的產角質酶的優良菌株,在未添加角質誘導時產酶量可以達到7.932U/mL,比一般的真菌產酶量高。該菌種可從棉花或其生長的土壤中分離是一種專化性較強、寄主范圍較窄的病菌。本研究在對該菌株進行生長和發酵產角質酶的環境條件研究的基礎上,進行了添加角質對該菌產酶能力的提高以及連續激活真菌的角質酶基因,誘導其轉錄表達的研究。研究表明,誘導作用可以顯著地提高角質酶的產量,產酶量可比無誘導時提高至4倍。
1 材料與方法
1.1 菌種與試劑
棉花枯萎病菌(Fusarium oxysporum):天津農學院植保研究室提供。
對硝基苯丁酸酯(p-nitrophenyl butyrate,PNB)購自美國Sigma公司;NaNO3、K2HPO4、MgSO4、KCl、FeSO4·7H2O、瓊脂、葡萄糖等試劑均為國產分析純。
1.2 培養基
種子培養基:PDA培養基,將土豆洗凈去皮,再稱取200g馬鈴薯并切成小塊,加1000mL蒸餾水煮爛,用8層紗布過濾,補足水分至1000mL,加熱,加入17g瓊脂,繼續加熱攪拌混勻,待瓊脂溶解完后,加入20g葡萄糖,攪拌均勻,分裝試管或者錐形瓶,加塞、包扎,121℃滅菌20min左右后取出試管擺斜面或者搖勻,冷卻后貯存備用。
土豆汁:200g土豆,在沸水中浸煮20min,過濾,濾液置于冰箱中,備用。
發酵基礎培養基[16]:葡萄糖1.0g/L,NaNO30.6g/L,K2HPO40.6g/L,MgSO40.2g/L,KCl 0.2g/L,FeSO4·7H2O 0.1g/L,角質0.1%。
礦物質培養基:NaNO30.6g/L,K2HPO40.6g/L,MgSO40.2g/L,KCl 0.2g/L,FeSO4·7H2O 0.1g/L。
1.3 角質的制備
采用蕃茄角質為誘導角質。蕃茄表面有韌性更強的角質,且容易大量獲得。蕃茄角質的制備:將蕃茄于沸水中煮5min,驟冷分離取皮,并將皮于草酸緩沖液(草酸4g/L、草酸銨16g/L,pH3.8)中煮沸1.5h,用蒸餾水洗凈皮上粘性物質。于50℃干燥箱中烘干至質量恒定,磨碎備用。
1.4 培養方法
種子培養:將儲于冰箱中的菌種接入斜面種子培養基上,在30℃培養48h,然后接種于裝有100mL液體PDA的250mL錐形瓶中,30℃、160r/min搖床培養36h。
發酵培養:將發酵培養基分裝于數個250mL錐形瓶中,每瓶裝100mL。取培養36h的種子培養液2mL,加入其中,30℃、160r/min搖床培養。培養完成后將發酵液用12層紗布過濾,濾液再于4℃條件下5500r/min離心30min,上清即為粗酶液,用于酶活測定。
1.5 酶活測定
酶活測定[14]:以對硝基苯丁酸酯(PNB)為底物測定酶的活力。反應體系為1.8mL,包括200μL的粗酶液、200μL 0.4%的Tritonx-100、1380μL 50mmol/L磷酸鹽緩沖液(pH 7.0)和20μL 1.76%的PNB溶液。于37℃反應10min后,在波長405nm處測定吸光度值。酶活定義:在反應條件下,將每毫升酶液每分鐘產生1μg的對硝基苯酚定義為1個酶活單位,U/mL。酶活計算公式:
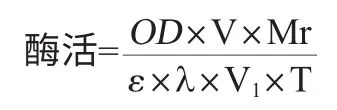
式中:OD為吸光值;V為反應體系的體積,1.8mL;Mr為產物對硝基苯酚的分子量,139.11g/mol;ε為摩爾消光系數,6830L/(mol·cm);λ為比色皿的光程,0.5cm;V1為酶液體積,200μL;T為反應時間10min。
1.6 連續誘導馴化實驗方法
連續誘導操作工藝流程:
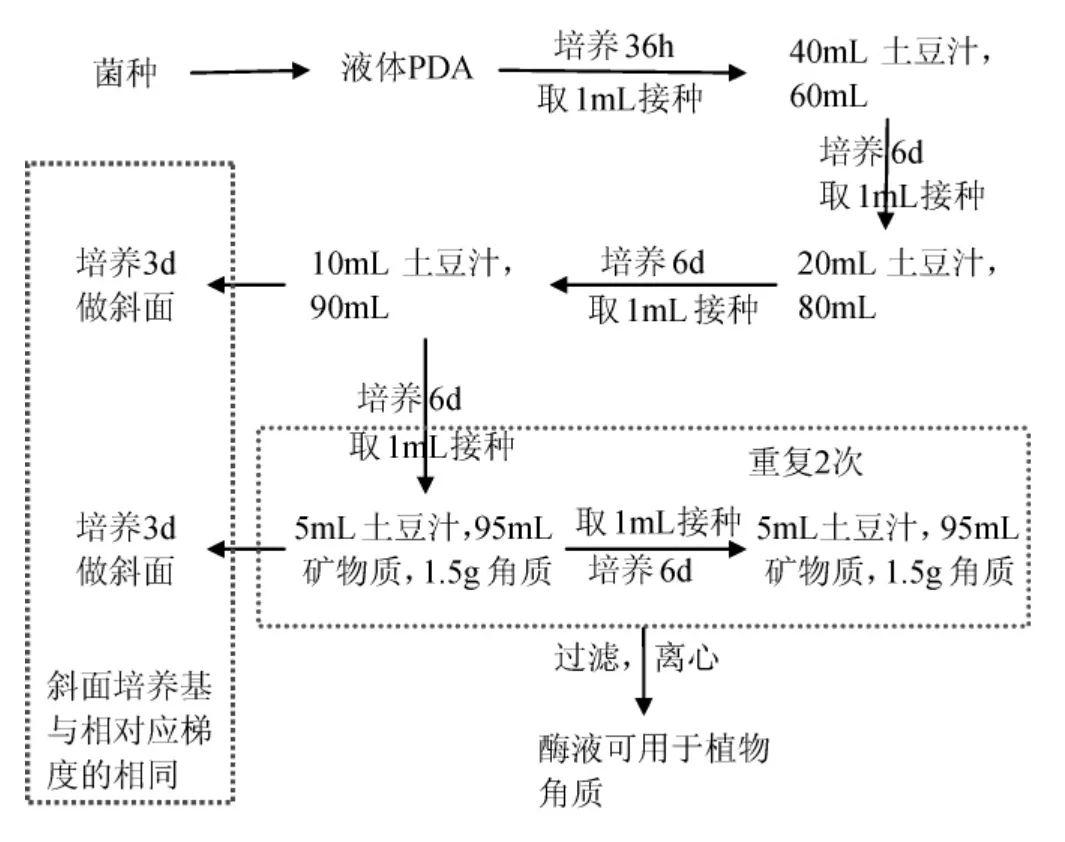
逐漸降低培養基中土豆汁的含量,逐漸提高角質的含量,形成培養基中碳源角質與土豆汁之間的含量比值變化的梯度環境。使得棉花枯萎病菌(F.oxysporum)在該環境條件下,被迫分泌產生更多的角質酶以適應新的生長環境。這樣可以使得真菌的角質酶基因一直處于亢奮的激活狀態,角質酶基因的轉錄量維持在逐步升高的水平,角質酶的產量也就隨之逐步升高。
發酵培養基中土豆汁的添加量分別為40mL、20mL、10mL、5mL,礦物質溶液的添加量分別是60mL、80mL、90mL、95mL,而角質的添加量分別為0.5g、0.8g、1.0g、1.5g。將棉花枯萎病菌接種于第一梯度培養基中,培養6d后,接入下一梯度培養基中,依次作相同處理。
2 結果與討論
2.1 菌種接種種齡的確定
為了確定最佳接種時間,按照1.4所述的種子培養方法,分別培養12h、24h、36h、48h、60h、72h,培養液以5000r/min離心30min,傾去上清液,加入蒸餾水100mL,再離心。重復3次。傾去上清液,烘干至質量恒定,稱質量,得細胞干質量與時間的關系見圖1。
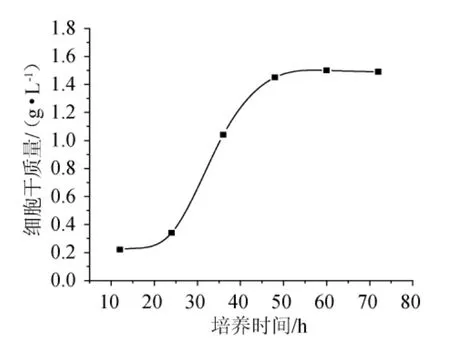
圖1 時間對菌體生長的影響Fig.1 Effect of time on bacteria growth
由圖1可知,顯示了72h內菌體生長的變化情況,由生物量變化可知,菌體培養至25h~50h時處于對數生長期,對數生長期時菌體以一個恒定的最大的比生長速度生長,細胞數量呈指數遞增,生長繁殖旺盛因此對數期可以獲得健壯且數量眾多的發酵種子。本實驗選擇菌體培養36h的處于對數生長期的菌體接種。
2.2 發酵溫度及培養基初始pH值的確定
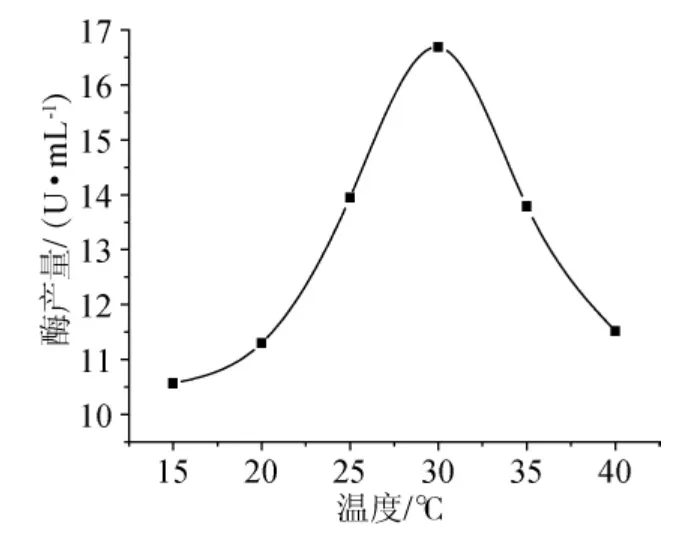
圖2 培養溫度對產角質酶的影響Fig.2 Effect of temperature on producting cutinase
為了考察溫度對菌株產角質酶的影響,以基礎培養基為營養,分別于15℃、20℃、25℃、30℃、35℃、40℃等溫度條件下發酵培養,培養6d后測酶活,結果見圖2。從圖2可以發現,30℃發酵培養時可以獲得最高的角質酶酶活,說明此溫度條件下棉花枯萎病菌表達的角質酶量達到最大。因此,可該菌確定產角質酶酶的最適溫度為30℃。
菌株和酶的活性和其活性狀態都與環境的pH值有著密切的關系,培養基的pH值是影響微生物菌種生長和生產的一個重要的環境因素。在研究中將基礎培養基的初始pH值分別調至6.5、7.0、7.5、8.0、8.5、9.0、9.5進行發酵培養,培養6d后測酶活,結果見圖3。

圖3 初始pH值對產角質酶的影響Fig.3 Effect initial pH value on cutin enzyme production
圖3確定了該菌產角質酶的最佳初始pH值,表明在pH 6.5~8.5范圍內,隨著pH值升高,酶活也逐漸升高,pH值>8.5以后,隨pH值的升高酶活力降低,所以pH 8.5是該菌產酶最佳酸堿度。
2.3 培養時間及角質對酶的產生的影響
將斜面的菌種活化,接入裝有100mL發酵培養基的250mL錐形瓶中培養至對數期,再按2%接種量接入發酵培養基中,30℃、160r/min搖床振蕩培養,分別在1d、2d、3d、4d、5d、6d、7d、8d取樣、過濾、離心、測酶活,且每個樣品做3個平行樣。以不加角質樣作為對照,結果見圖4。
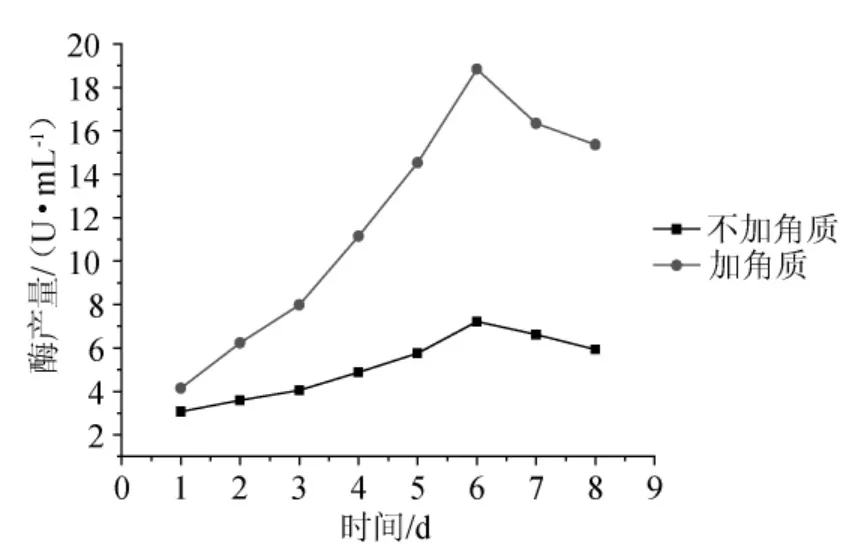
圖4 培養時間及角質對產酶的影響Fig.4 Effect of training time and cutin on enzyme production
圖4表明,棉花枯萎病菌經發酵培養,角質酶的活性在第6d時達到最高,之后隨著發酵時間的增加活性逐漸降低,因此可確定產酶最佳時間為6d。從圖4還可以看出,在培養基中添加0.2%的蕃茄角質后,菌種產角質酶的量明顯有所提高,比未添加角質時提高了3倍,產酶量最高達到29.46U/mL,比未誘導時提高至4倍。因為其產酶量提高4倍的情況還不夠穩定,因此本文選擇了產酶量提高的平均水平,即19.06U/mL,比未添加時提高3倍。這充分說明角質酶是誘導酶,微量的角質就對菌種產角質酶起到了明顯的誘導作用,利用角質誘導棉花枯萎病菌(F.oxysporum)產角質酶,這也與角質酶是一種誘導酶的結論是相符的,也說明誘導法是簡便可行的有效方法。這充分說明角質酶是誘導酶,微量的角質就對菌種產角質酶起到了明顯的誘導作用。
2.4 連續誘導
菌種的馴化是通過人工措施使微生物逐步適應某一環境,而定向選育微生物的方法,馴化獲得具有較高耐受力及活動能力的菌株。目前報道較多的例子是用于廢水處理中的微生物,通過馴化得到對某種污染物具有較高降解能力和耐受力的高效菌株。
酶的合成受到基因表達的調控,也就是說,生物在生長發育過程中,基因表達可按一定時間程序發生改變,而且隨著內外環境的變化而加以調控,這就是基因表達的時序調節和適應調節,基因表達的調節可以在不同水平(包括轉錄前、轉錄和轉錄后)或在翻譯的水平(包括翻譯和翻譯后)[17]。當在培養基中加入誘導物,誘導物能夠激活其基因的轉錄表達,合成有關的信使核糖核酸(mRNA),并翻譯成誘導酶。
按照操作流程,每一梯度發酵后,培養液以5000r/min轉速離心30min,傾去上清液,再加入蒸餾水100mL,再離心。重復3次。傾去上清,烘干至質量恒定,稱質量,結果見圖5~圖7。

圖5 菌種在各梯度下生長情況Fig.5 Bacteria growth in the gradient
在研究中設計了一個梯度環境,此環境的培養基中碳源角質與土豆汁之間的含量對應變化。逐漸降低培養基中土豆汁的含量,并逐漸提高角質的含量,使得較易被棉花枯萎病菌(F.oxysporum)利用的碳源土豆汁的含量不斷減少,而角質含量逐漸增加,使棉花枯萎病菌需要分泌產生更多的角質酶來適應新的環境。在這過程中如果枯萎病菌能利用角質作為碳源,這將更有利于角質酶的表達。梯度環境可以使得棉花枯萎病菌(F.oxysporum)的角質酶基因一直處于激活狀態,角質酶基因的轉錄量隨著環境中角質含量的增加而逐步升高,角質酶的產量也會維持在一個逐步升高的水平上。
從圖6可以看出,隨著培養基中土豆汁含量的減少,菌體細胞干質量隨之減少,說明土豆汁與棉花枯萎病菌(F.oxysporum)生長量有著密切相關的關系,也說明了棉花枯萎病菌不能像利用土豆汁那樣利用角質進行菌體的生長繁殖,角質還不能完全替代土豆汁作為碳源。但從結果(圖7)可以看出,培養基中產的角質酶并沒有隨真菌生物量的減少而降低,反而有明顯的增加。說明了連續的誘導可以提高棉花枯萎病菌(Fusarium oxysporum)產酶的能力。加入角質連續誘導后,角質酶的產量有了明顯的提高,一般情況可以提高3倍,最高可以達到29.46U/mL,比未誘導時產酶能力提高至4倍。
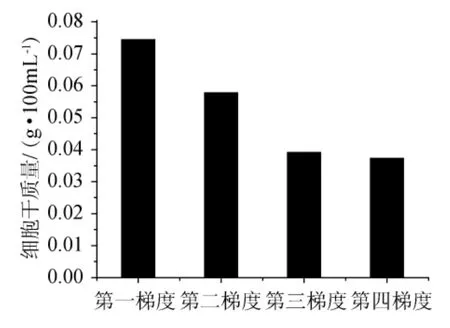
圖6 土豆汁含量對菌體干質量的影響Fig.6 Effect of potato juice content on dry cell weight
由圖6、圖7可知,當營養相對豐富時,真菌的生物量能夠得到一定的積累,經過角質誘導后,產角質酶的量就會大大的提高;角質酶是一種誘導酶,角質能夠激活角質酶基因的轉錄表達,不斷降低其他易被利用的碳源含量并同時提高角質在其中的含量,使得棉花枯萎病菌始終處于產酶較高的水平。
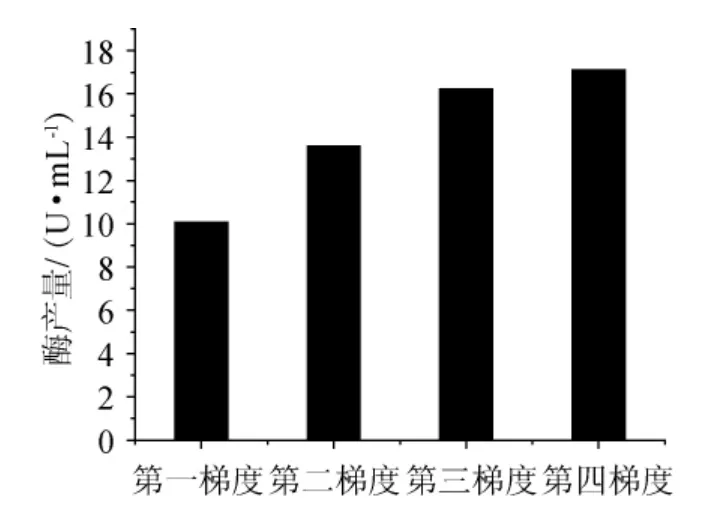
圖7 真菌生物量對酶活的影響Fig.7 Effect of fungal biomass on the enzyme activity
在發酵產酶過程中,為了提高酶的產量,除了選育優良的產酶細胞,保證發酵工藝條件并根據需要和變化情況進行及時調節控制以外,還可以采取某些行之有效的措施如添加誘導物、控制阻遏物濃度、添加表面活性劑或其他產酶促進劑等[18]。
對于誘導發酵生產,在發酵培養基中添加適量誘導物,使產酶量顯著提高。誘導物一般可分為3類:酶的作用底物、酶的催化反應產物、酶的底物類似物。根據文獻報道,角質酶是一種誘導酶,其誘導物是角質酶的作用底物角質。本實驗也證實了這一點,相對于無角質存在的發酵過程,添加角質后的產酶量得以顯著的提高。對照圖6、圖7,可以得出這樣的結論,角質酶活性的提高并不是因為棉花枯萎病菌繁殖量的提高,而是角質加入量的增加(圖6、圖7中的第四梯度)提高了菌株對角質酶的表達。
3 結論
角質酶的基因在受到角質的誘導作用下被激活,會表達產生更多的角質酶。本實驗采用連續誘導法培養菌株,控制培養基中的土豆汁與角質的含量,使得菌株一直處于一個有角質存在的環境中。而且土豆汁的含量越來越少,角質越來越多,這就說明角質酶活的增加不是菌株生物量的增加帶來的,而是角質誘導的作用使菌株的角質酶表達能力提高了。在這樣的環境下角質酶基因的表達會因角質的存在保持在一個較高的水平上,從而達到保持菌種產角質酶能力的一個較高水平。
角質酶的應用十分廣泛,其在食品工業、化工工業及印染工業等許多領域都有應用。近幾年還發現,角質酶可用于化纖面料的表面修飾及棉散纖維前處理[11]。但沒有大規模的工業生產角質酶,因此提高角質酶產量的研究具有廣泛的理論與實踐意義。
[1]HOLMQUIST M.Alpha/Beta-hydrolase fold enzymes:structures,functions and mechanisms[J].Curr Protein Pept Sc,2000,1(2):209-235.
[2]EGMOND MR,De VLIEG J.Fusarium solani pisi cutinase[J].Biochimie,2000,82:1015-1021.
[3]KOLATTUKUDY PE,PURDY RE,MAITI IB.Cutinases from fungi and pollen[M].//BORG STR?M B.,BROCKMAN H.(eds.)Lipases.Amsterdam:Elsevier,1984,66:71-504.
[4]MURPHY CA,CAMERON JA,HUANG SJ,et al.Fusarium polycaprolactone depolymerase is cutinase[J].Appl Environ Microb,1996,62(2):456-460.
[5]MARTINEZ C,P De GEUS,LAUWERWYS M,et al.Fusarium solani cutinaseis a lipolytic enzyme with a catalytic serine accessible to solvent[J].Nature,1992,356:615-618.
[6]TADAYUKI S[P].J P pat,08092023,1996-09-04.
[7]KIM YH,AHN JY,MOON SH,et al.Biodegradation and detoxification of organophosphate insecticide malathion byFusarium oxysporumf.sp.pisi cutinase[J].Chemosphere,2005,60:1349-1355.
[8]FETT WF,GéRARD HC,MOREAU RA.Cutinase production byStreptomycesspp[J].Curr Microbiol,1992,25(3):165-171.
[9]JOHAN S.Laundry detergent and/or fabric care compositions comprising a modified transferase[P].US Patent:1999057254,1999.
[10]SHIMAO M.Biodegradation of plastics[J].Curr Opin Biotech,2001,12(3):242-247.
[11]郭 森.重組大腸桿菌產角質酶-CBM[D].無錫:江南大學碩士論文,2012.
[12]查理德N,斯特蘭特.彭友良譯.植物病理學導論[M].北京:化學工業出版社,2007.
[13]張守亮,陳 堅.角質酶產生菌Thermobifida fuscaWSH03-11 誘變及高產突變株發酵條件優化[J].化工進展,2006,25(5):533-537.
[14]DAVIES KA,DE LORONO I,FOSTER SJ,et al.Evidence for a role of cutinase in pathogenicity ofPyrenopeziza brassicaeon brassicas[J].Physiol Mol Plant P,2000,57(2):63-75.
[15]張芙華,陳 晟,張東旭,等.pH 兩階段控制策略發酵生產重組角質酶[J].中國生物工程雜志,2008,28(5):59-64.
[16]SHERYL F,GOTE J,RAJNI HK,Purification of recombinant cutinase by extraction in an aqueous two-phase system facilitated by a fatty acid substrate[J].Biotechnol Bioeng,73(6):465-475.
[17]楊志敏,蔣立科.生物化學[M].北京:高等教育出版社,2005.
[18]刑淑婕,夏新奎.酶工程[M].北京:高等教育出版社,2008.
