戊糖乳桿菌素pentocin LPEM818的初步純化及特性研究*
2013-05-05 11:27:20于雷雷王超施波馬妙蓮段慧張明
食品與發酵工業 2013年2期
關鍵詞:實驗
于雷雷,王超,施波,馬妙蓮,段慧,張明
(安徽農業大學生命科學學院,安徽合肥,230036)
細菌素(bacteriocin)是一些細菌產生的抗菌多肽或蛋白[1],因其能被人類胃腸道分泌的蛋白酶降解,對人體無毒副作用,所以具有作為食品生物防腐劑的潛在應用價值,乳酸乳球菌素nisin在乳制品及罐頭制品中已得到了廣泛的應用[2]。戊糖乳桿菌 (Lactobacillus pentosus)[3]是公認的食品安全菌,因而對戊糖乳桿菌細菌素戊糖乳桿菌素進行了大量研究,有關戊糖乳桿菌素的報道最早見于1999年有關pentocin TV35b的分離純化[4],我國在2004年首次報道了pentocin 31-1的分離純化[5-9]。已報道的產細菌素戊糖乳桿菌有 L.pentosus TV35b[4]、L.pentosus 31 -1[5-9]、L.pentosus ST151BR[10]、L.pentosus B96[11]、L.pentosus ST712BZ[12]、L.pentosus S-PT84[13]、L.pentosus b240[14]、L.pentosus IG1[15]、L.pentosus[16]、L.pentosus C50-6[3]、L. pentosus 191 和 L. pentosus 204[17]等。
戊糖乳桿菌素多屬于II類細菌素,分子量較小、熱穩定性高、主要抑制革蘭氏陽性菌[4],但pentocin C50-6還能抑制革蘭氏陰性菌[3],在N-末端具有 IIa類細菌素的保守序列YGNGV,對單核細胞增生李斯特氏菌(Listeria monocytogenes)ATCC19115有較強的抑制作用[5]。L.pentosus B96和L.pentosus IG1所產的IIb類細菌素均由2個高度疏水、對熱穩定的小分子肽組成,2個組分在抑菌功能上具有協同作用[11,15]。Pentocin C50-6 由于沒有進行序列測定,僅根據分子量和抑菌譜的分析推測其可能屬于抑菌譜較廣的I類或II類細菌素[3],暫未發現屬于IIc類、IId類、III類和IV類的戊糖乳桿菌素。細菌素的抑菌方式一般分為抑菌、殺菌不溶菌和殺菌且溶菌3種類型,研究表明現已報道的pentocin TV35b和pentocin 31-1對指示菌的的作用方式均為殺菌不溶菌[4-5]。戊糖乳桿菌素由于可耐受高溫處理,且大部分在酸性或弱堿性條件下穩定[2-9],因而在食品領域具有廣泛的應用前景。本文主要探討了戊糖乳桿菌LPEM818細菌素的生物學特性以及對指示菌的作用方式等,以待為該細菌素的應用和進一步研究提供理論依據。
1 材料和方法
1.1 實驗材料
1.1.1 菌株
產細菌素菌株:戊糖乳桿菌(Lactobacillus pentosus)LPEM818為安徽農業大學生命科學學院實驗室篩選保藏;指示菌株見表2。
1.1.2 培養基
MRS培養基和 LB培養基[18]。
1.1.3 主要藥品試劑
胃蛋白酶(BBI),木瓜蛋白酶(Sangon),脂肪酶(Sigma),過氧化氫酶(Sigma),凝乳蛋白酶(BBI),胰蛋白酶(BBI),蛋白酶K(Sigma),Sephadex G-25(GE Healthcare),BSA(BBI),Folin-酚試劑(Sangon),尿素(BBI),EDTA(BBI),SDS(BBI)等。
1.2 實驗方法
1.2.1 Pentocin LPEM818的活性及效價的測定
采用牛津杯雙層平板法[19]和雙層平板二倍稀釋法[20]。
1.2.2 蛋白質濃度的測定
Lowry 法[21]。
1.2.3 Pentocin LPEM818 的初步純化[22]
接種6環菌株LPEM818于MRS培養基中,37℃培養14 h作為種子液。按4%的體積比將種子液接種于4 L MRS培養基中,37℃培養24 h。將發酵液離心,過濾(0.45 μm),上清液進行80% 硫酸銨沉淀,4℃過夜,離心取沉淀,重懸于 ddH2O中,用透析袋(500 u)透析脫鹽,經氯化鋇檢測不含硫酸銨,收集透析液,測定其效價并冷凍干燥,得到 pentocin LPEM818干粉。將pentocin LPEM818干粉重懸于ddH2O,得到效價為1 060AU/mL的樣品,取5 mL上Sephadex G-25(1.6 cm×100 cm)凝膠層析,流速為0.6 mL/min,流動相為ddH2O。收集各峰進行活性檢測,確定其活性峰。
1.2.4 Pentocin LPEM818的生物學特性研究
1.2.4.1 pH值和熱穩定性實驗
樣品用1 mol/L HCl和1 mol/L NaOH分別調pH值至 2.0、4.0、6.0、8.0、10.0 和 12.0,補加 ddH2O 使樣品終體積相等,以ddH2O同比稀釋的樣品作為對照,每組設3個平行,37℃水浴6 h后,調 pH值至5.0,檢測其抑菌活性。
樣品分別進行 4、37、60、80、100 和 121℃水浴 15 min或30 min,以未經處理的樣品為對照,每組設3個平行,冰浴10 min后檢測其抑菌活性。
1.2.4.2 酶和有機溶劑敏感性實驗
樣品用1 mol/L HCl和1 mol/L NaOH分別調至各酶最適pH值,使各酶的最終濃度為5 mg/mL,以ddH2O同比稀釋的酶和同比稀釋的樣品分別作為對照,每組設3個平行,37℃水浴8h,沸水浴10 min,檢測抑菌活性。
樣品分別加入1%體積比的 SDS、尿素、NaCl、EDTA、Triton X-100、吐溫-80和乙腈,以 ddH2O 同比稀釋的有機溶劑和同比稀釋的樣品分別作為對照,每組設3個平行,37℃水浴4h,檢測抑菌活性。
1.2.4.3 抑菌譜的測定
采用牛津杯雙層平板法,分別對供試G+菌株和G-菌株做抑菌實驗。
1.2.4.4 作用方式的測定
將菌株533接種于LB培養基中,37℃培養14 h(OD600=2.89)作為種子液,按4%的體積比接種于LB培養基,分成兩組,一組在8h后加入 pentocin LPEM818樣品,另一組加等體積的無菌生理鹽水作為對照,37℃培養26 h,每2 h取出5 mL,測定OD600,計算菌落數(CFU/mL),每次測3個平行。
1.2.4.5 生理曲線的測定
將菌株LPEM818的種子液按4%的體積比接種于MRS培養基中,37℃培養。每隔2 h取樣,測定OD600,pH值和上清液抑菌活性,各設3個平行。
2 結果與分析
2.1 Pentocin LPEM818的初步純化
為了對pentocin LPEM818進行初步純化,本實驗以80%硫酸銨沉淀樣品為材料,進行了Sephadex G-25(1.6 cm×100 cm)凝膠層析分析(圖1)。上樣量為5 mL,流速為0.6 mL/min,流動相為 ddH2O,收集各峰并測定活性。
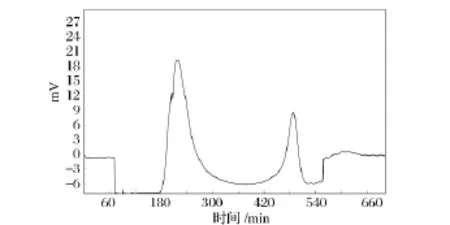
圖1 Sephadex G-25層析圖譜Fig.1 The chromatogram of G-25
經G-25凝膠層析后,樣品有2個峰(圖1),抑菌實驗證實第2個峰為活性峰,而第1個峰無抑菌活性,為雜蛋白峰,達到了一定的分離效果。說明硫酸銨沉淀分離法用于初步分離該細菌素是可行的。經過硫酸銨沉淀和G-25層析分離純化后,該細菌素的純化倍數為11.09倍,回收率為6.4%(表1)。經Tricine-SDS-PAGE電泳后,出現多條蛋白質條帶(結果未顯示),分子質量在6.5~27 ku,說明該細菌素為小分子肽。可能由于樣品上樣量太少,導致未檢測到有抑菌活性的條帶;要獲取單一條帶,有待進一步優化分離純化條件。
2.2 Pentocin LPEM818的pH值和熱穩定性實驗
對80%硫酸銨沉淀樣品分別在不同的pH值條件下,37℃水浴6 h后,調pH值至5.0,檢測抑菌活性,以獲得該細菌素的pH穩定性范圍。相同樣品經不同的溫度處理15 min或30 min,冰浴10 min,檢測抑菌活性,以獲得該細菌素的熱穩定性范圍。
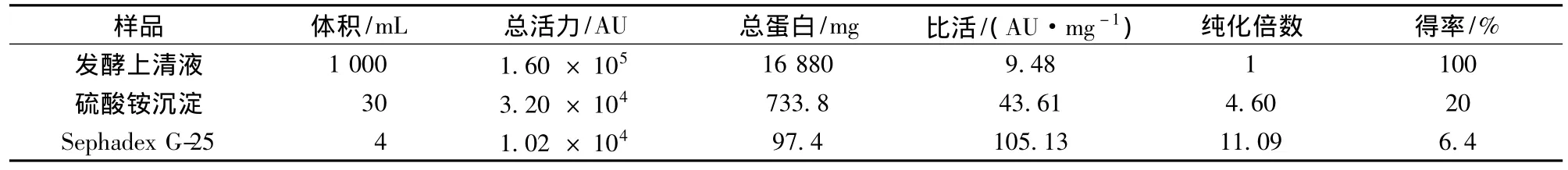
表1 Pentocin LPEM818的純化表Table 1 Purification table for pentocin LPEM818
pH穩定性實驗結果(圖2)表明,樣品在pH 4.0~6.0條件下抑菌活性最高,其保留值約為90%,在pH 2.0和8.0條件下抑菌活性仍保留在80%,當pH在10.0或12.0時抑菌活性完全喪失。因此,該細菌素具有較好的酸和弱堿穩定性。熱穩定性實驗結果(圖3)表明,樣品經4~80℃處理后,抑菌活性保留在90%以上。經100℃處理30 min和121℃處理15 min后,抑菌活性保留在80%以上,而經121℃處理30 min后,其抑菌活性仍保留在66.98%。因此該細菌素是一種熱穩定型的細菌素。
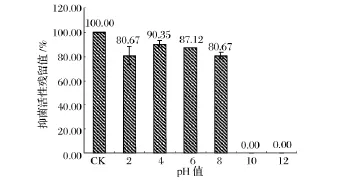
圖2 不同pH對樣品抑菌活性的影響Fig.2 The antibacterial activity of sampleat the different pH
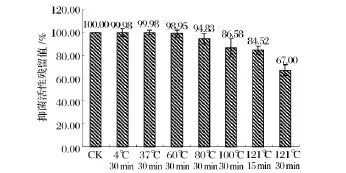
圖3 不同溫度對樣品抑菌活性的影響Fig.3 The antibacterial activity of sample at the different temperature
2.3 Pentocin LPEM818對酶和有機溶劑的敏感性實驗
對80%硫酸銨沉淀樣品分別經不同的酶在37℃條件下處理8 h,沸水浴10 min,檢測抑菌活性,以確定該細菌素的酶敏感性。相同樣品經不同的有機溶劑在37℃條件下處理4 h,檢測其抑菌活性,以確定該細菌素的有機溶劑敏感性。
對酶敏感性實驗結果(圖4)表明,樣品經蛋白酶K和胰蛋白酶處理后抑菌活性完全喪失,所以對其敏感;樣品經凝乳蛋白酶處理后抑菌活性下降15%,所以對其較敏感;樣品經脂蛋白酶、胃蛋白酶、木瓜蛋白酶處理后抑菌活性基本不變,所以對其不敏感,因此該細菌素為蛋白類抑菌物質。對有機溶劑敏感性實驗結果(圖5)表明,樣品經尿素處理后抑菌活性下降14%,所以對其較敏感;樣品經 Triton X-100、NaCl、吐溫-80和乙腈處理后抑菌活性不變,所以對其不敏感;該細菌素與SDS或EDTA共同作用時,抑菌效果明顯好于單一作用,因此二者具有協同抑菌作用。
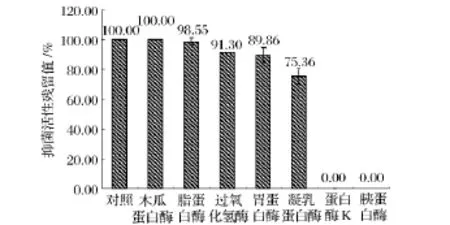
圖4 不同酶對樣品抑菌活性的影響Fig.4 The antibacterial activity of sample at the different enzymes
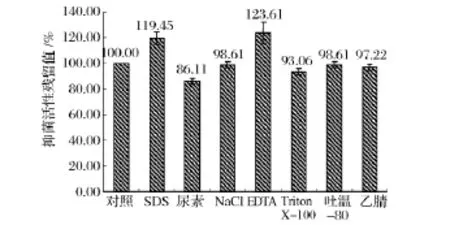
圖5 不同有機溶劑對樣品抑菌活性的影響Fig.5 The antibacterial activity of sample at the different organic solvent
2.4 Pentocin LPEM818抑菌譜的測定
用80%硫酸銨沉淀樣品分別對供試的G+菌株和G-菌株做抑菌實驗(表2),測定抑菌圈直徑,以確定該細菌素抑菌譜。

表2 Pentocin LPEM818的抑菌譜Table 2 Antibacterial spectrum of pentocin LPEM818
硫酸銨沉淀樣品的抑菌譜測定結果(表2)表明,pentocin LPEM818對供試的G-菌株變形桿菌、雞白痢沙門氏菌、綠膿桿菌和G+菌株單核細胞增生李斯特氏菌、金黃色葡萄球菌、藤黃微球菌、四聯球菌的抑制作用明顯;對糞腸球菌、發酵乳桿菌、保加利亞乳桿菌、短乳桿菌等近源乳酸菌也有較好的抑制作用。
2.5 作用方式的測定
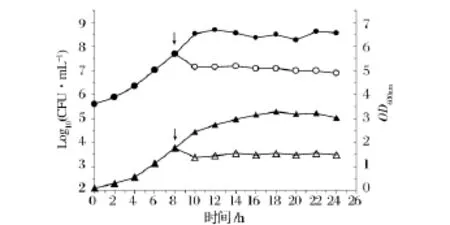
圖6 Pentocin LPEM818對S.pullorum533的作用方式Fig.6 The effect of pentocin LPEM818 on the growth of S.pullorum 533
將接種過菌株533的LB培養基置于37℃培養,每隔2 h取樣,測定其OD600值,監測菌體濃度的變化,同時測定CFU監測活菌數量,獲得菌體的生長情況(圖6),以確定該細菌素對指示菌的作用方式。
對菌株533作用方式的研究(圖6)表明,菌體在12 h后進入生長穩定期。實驗組菌株533的活菌數在10 h,從1×108降至1×107,隨后趨勢略有下降。說明細菌素在短時間內殺死了大量指示菌,該細菌素對指示菌的作用方式為殺菌。而對照組活菌數則呈持續上升趨勢,10h后保持穩定;實驗組菌株533的OD600在10 h后基本保持恒定,說明該細菌素對指示菌無溶菌現象;而對照組OD600一直呈上升趨勢,直至16 h后穩定。因此,該細菌素對指示菌的作用方式僅為殺菌。
2.6 生理曲線的測定
將接種過菌株 LPEM818的 MRS培養基置于37℃培養,每隔2 h取樣,測定OD600值,pH值和上清液抑菌活性,以測定該菌株的生理曲線。
菌株LPEM818生理曲線的測定結果(圖7)表明,菌株LPEM818置于37℃培養,6 h進入對數中期,16 h進入穩定期,此時OD600值最大,22 h進入衰亡期。發酵上清液的pH值從6.24降至4.50,18 h后保持恒定。在對數期中期開始產生細菌素,且在穩定期后期產量達到最大值,此時抑菌活性達到最高,隨后保持穩定,整個過程受pH的影響較小。
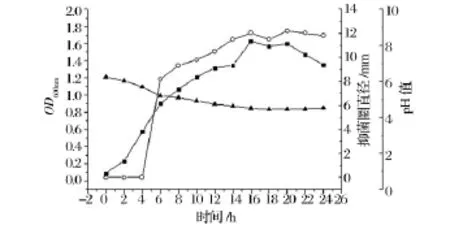
圖7 L.pentosus LPEM818的生理曲線Fig.7 Production of pentocin LPEM818 during growth of L.pentosus LPEM818
3 討論與結論
3.1 Pentocin LPEM818的初步純化
細菌素的分離純化與其特性研究密切相關。細菌素的初步分離方法有鹽析法、有機溶劑沉淀法和pH吸附法等,進一步純化方法有凝膠過濾層析、離子交換層析、疏水相互作用層析和反相高效液相色譜等。本研究采用硫酸銨沉淀和Sephadex G-25凝膠層析對pentocin LPEM818進行了初步純化,最終純化倍數為11.09倍,但回收率僅為6.4%,說明純化過程中目的蛋白損失較大。今后應繼續對pentocin LPEM818的分離純化條件進行優化,為將來的精細純化及應用奠定基礎,但是根據其目的蛋白峰,確定了硫酸銨沉淀法樣品用于生物學特性研究的可行性。
3.2 Pentocin LPEM818的作用方式
細菌素通常對敏感菌株的作用方式表現為3種類型:抑菌、殺菌不溶菌和殺菌且溶菌。本研究在S.pullorum 533的菌懸液中加入80%硫酸銨沉淀樣品后,在短時間內對S.pullorum 533的活菌數產生了有效的抑制作用,并且在加入該細菌素后OD600無明顯變化,因此該細菌素對S.pullorum 533的作用方式僅為殺菌。具有與本實驗結果相同作用方式的細菌素有 bacTN635[23]和 plantaricin MG[24],其中 bacTN635的指示菌活菌數下降了6個單位,而OD600無明顯變化;plantaricin MG的指示菌活菌數下降了96.3%,而OD600也無明顯變化。而 plantaricin TF711[25]的指示菌活菌數在3h內下降了1.3個單位,隨后有所上升,9h后保持穩定,其作用方式為抑菌。所以pentocin LPEM818比bacTN635或plantaricin MG的抑菌效果稍差,但優于plantaricin TF711。
3.3 Pentocin LPEM818的生物學特性
細菌素的生物學特性決定其潛在的應用價值。Pentocin LPEM818在pH 2.0~8.0條件下穩定,具有酸和弱堿穩定性的特點,與enterocin LR/6在pH 2.0-8.0條件下抑菌活性保留80%[26]基本相同,但其熱穩定性不及pentocin LPEM818。Pentocin LPEM818在121℃處理30 min仍保留67%的抑菌活性,具有較好的熱穩定性。具有相似特性的pentocin 31-1在100℃處理 15 min只保留 50%的抑菌活性[7]。pentocin LPEM818熱穩定性明顯優于pentocin 31-1。pentocin LPEM818對酶的敏感性的特點與Pediococcus pentosaceus CFR SIII所產的細菌素相似,僅對胰蛋白酶和蛋白酶K敏感[27],但其pH穩定性范圍為3.0~5.0明顯差于本細菌素。對吐溫-80、NaCl、乙腈和 Triton X-100不敏感,對尿素敏感的特點與sakacin LSJ618[28]相一致,進一步表明了該細菌素具有蛋白屬性。EDTA或 SDS本身對S.pullorum 533具有抑制作用,pentocin LPEM818與SDS或EDTA共同作用時,抑菌效果明顯好于單一作用,因此二者有協同抑菌作用[28-30]。在實驗條件下,菌株 LPEM818在對數中期開始產生細菌素,在穩定期后期產量達到最大值,隨后保持穩定,產細菌素過程受環境pH的影響較小。該細菌素對供試的大部分G+菌株和G-菌株均具有較強抑制作用,其抑菌譜廣,對多數致病菌和食品腐敗菌抑菌作用明顯,特別是對Listeria monocytogenes ATCC19115和G-菌株較強的抑制作用,所以其具有作為食品生物防腐劑的潛在應用價值。
[1] 郭興華.益生乳酸細菌:分子生物學及生物技術[M].北京:科學出版社,2008.
[2] 呂燕妮,李平蘭,孫成虎,等.戊糖乳桿菌31-1菌株所產細菌素的理化及生物學特性[J].中國農業大學學報,2006,11(1):39-43.
[3] 劉書亮,敖靈,周佳,等.戊糖乳桿菌素 C50-6的純化及特性研究[J].食品與發酵工業,2010,36(5):36-40.
[4] Okkers D J,Dicks L M,Silvester M,et al.Characterization of pentocin TV35b,a bacteriocin-like peptide isolated from Lactobacillus pentosus with a fungistatic effect on Candida albicans[J].Journal of Applied Microbiology,1999,87(5):726-734.
[5] Zhang J,Liu G,Shang N,et al.Purification and partial amino acid sequence of pentocin 31-1,an anti-listeria bacteriocin produced by Lactobacillus pentosus 31-1[J].Journal of Food Protection,2009,72(12):2 524-2 529.
[6] Liu G R,Griffiths M W,Shang N,et al.Applicability of bacteriocinogenic Lactobacillus pentosus 31-1 as a novel functional starter culture or coculture for fermented sausage manufacture[J].Journal of Food Protection,2010,73(2):292-298.
[7] Liu G R,Lv Y,Li P L,et al.Pentocin 31-1,an anti-Listeria bacteriocin produced by Lactobacillus pentosus 31-1 isolated from Xuan-Wei Ham,a traditional China fermented meat product[J].Food Control,2008,19(4):353 -359.
[8] Zhang J L,Liu G R,Li P L,et al.Pentocin 31-1,a novel meat-borne bacteriocin and its application as biopreservative in chill-stored tray-packaged pork meat[J].Food Control,2010,21(2):198 -202.
[9] Zhou K,Zhou W,Li P,et al.Mode of action of pentocin 31-1:An antilisteria bacteriocin produced by Lactobacillus pentosus from Chinese traditional ham[J].Food Control,2008,19(8):817-822.
[10] Todorov S D,Dicks L M T.Effect of medium components on bacteriocin production by Lactobacillus pentosus ST151BR,a strain isolated from beer produced by the fermentation of maize,barley and soy flour[J].World Journal of Microbiology& Biotechnology,2004,20(6):643-650.
[11] Delgado A,Brito D,Peres C,et al.Bacteriocin production by Lactobacillus pentosus B96 can be expressed as a function of temperature and NaCl concentration[J].Food Microbiol,2005,22(6):521 -528.
[12] Todorov S D,Dicks L M T.Bacteriocin production by Lactobacillus pentosus ST712BZ isolated from boza[J].Brazilian Journal of Microbiology,2007,38(1):166 -172.
[13] Izumo T,Izumi F,Nakagawa I,et al.Influence of Lactobacillus pentosus S-PT84 ingestion on the mucosal immunity of healthy and salmonella typhimurium-infected mice[J].Bioscience and Microflora,2011,30(2):27-35.
[14] Kotani Y,Shinkai S,Okamatsu H,et al.Oral intake of Lactobacillus pentosus strain b240 accelerates salivary immunoglobulin A secretion in the elderly:A randomized,placebo-controlled,double-blind trial[J].Immunity and Ageing,2010(7):11.
[15] Maldonado-Barragan A,Caballero-Guerrero B,Lucena-Padros H,et al.Genome sequence of Lactobacillus pentosus IG1,a strain isolated from Spanish-style green olive fermentations[J].Journal of Bacteriology,2011,193(19):5 605.
[16] Nieto-Lozano J,Reguera-Useros J,Pelaez-Martinez M,et al.Bacteriocinogenic activity from starter cultures used in Spanish meat industry[J].Meat Science,2002,62(2):237-243.
[17] Con A H,Gokalp H Y.Production of bacteriocin-like metabolites by lactic acid cultures isolated from sucuk samples[J].Meat Science,2000,55(1):89 -96.
[18] 張剛.乳酸細菌:基礎,技術和應用[M].北京:化學工業出版社,2007.
[19] 張艾青,劉書亮,敖靈.產廣譜細菌素乳酸菌的篩選和鑒定[J].微生物學通報,2007,34(4):753-756.
[20] 方佳琪,竺德強,柯芳芳,等.pH吸附法純化乳酸乳球菌素 Lacticin LLC518[J].食品與發酵工業,2011,37(10):52-56.
[21] 汪家政,范明.蛋白質技術手冊[M].北京:科學出版社,2000:42-91.
[22] Todorov S D,Rachman C,Fourrier A,et al.Characterization of a bacteriocin produced by Lactobacillus sakei R1333 isolated from smoked salmon[J].Anaerobe,2011,17(1):23-31.
[23] Smaoui S,Elleuch L,Bejar W,et al.Inhibition of fungi and gram-negative bacteria by bacteriocin BacTN635 produced by Lactobacillus plantarum sp.TN635[J].Applied Biochemistry and Biotechnology,2010,162(4):1 132-1 146.
[24] Gong H S,Meng X C,Wang H.Plantaricin MG active against Gram-negative bacteria produced by Lactobacillus plantarum KLDS1.0391 isolated from“Jiaoke”,a traditional fermented cream from China[J].Food Control,2010,21(1):89-96.
[25] Hernandez D,Cardell E,Zarate V.Antimicrobial activity of lactic acid bacteria isolated from Tenerife cheese:ini-tial characterization of plantaricin TF711,a bacteriocinlike substance produced by Lactobacillus plantarum TF711[J].Journal of Applied Microbiology,2005,99(1):77-84.
[26] Kumar M,Tiwari S K,Srivastava S.Purification and characterization of enterocin LR/6,a bacteriocin from Enterococcus faecium LR/6[J].Applied Biochemistry and Biotechnology,2010,160(1):40-49.
[27] Halami P M,Badarinath V,Manjulata Devi S,et al.Partial characterization of heat-stable,antilisterial and cell lytic bacteriocin of Pediococcus pentosaceus CFR SIII isolated from a vegetable source[J].Annals of Microbiology,2011,61:1-8.
[28] Jiang J,Shi B,Zhu D,et al.Characterization of a novel bacteriocin produced by Lactobacillus sakei LSJ618 isolated from traditional Chinese fermented radish[J].Food Control,2012,23(2):338 -344.
[29] Kopermsub P,Mayen V,Warin C.Potential use of niosomes for encapsulation of nisin and EDTA and their antibacterial activity enhancement[J].Food Research International,2011,44(2):605-612.
[30] Parente E,Giglio M A,Ricciardi A,et al.The combined effect of nisin,leucocin F10,pH,NaCl and EDTA on the survival of Listeria monocytogenes in broth[J].International Journal of Food Microbiology,1998,40:65-75.
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
