連續剪梢對金葉女貞不同葉位呈色和光合特性的影響
2013-08-02 00:52:10彭麗麗姜衛兵王明玉張斌斌
江蘇農業學報 2013年2期
彭麗麗, 姜衛兵, 韓 健, 王明玉, 張斌斌
(1.南京農業大學園藝學院,江蘇 南京 210095;2.江蘇省農業科學院園藝研究所,江蘇 南京 210014)
彩葉植物因其絢麗的葉色近年來在園林綠化中備受重視,應用越來越廣泛[1]。金葉女貞(Ligustrum vicaryi L.)作為一種常見的彩葉植物,原分布于中國長江以南地區,現已引種到中國北方城市,在城市和園林綠化中發揮著重要作用[2]。金葉女貞單葉片的葉色是隨著葉齡變化的,即幼葉呈金黃色(淺黃色),成熟葉呈黃綠色,老葉呈綠色。由于在同一枝條、同一植株上同時存在不同葉齡的葉片,所以同時呈現出金黃、黃綠、綠色等不同葉色的葉片,且自然生長情況下著生葉位具有規律性,即枝條上部(陽光直射處)葉片呈金黃色,枝條中部(樹冠中部)葉片呈黃綠色,枝條下部(樹冠內堂和基部)葉片呈綠色。
在城市和園林綠化管理中,整形修剪是一些彩葉樹木栽培過程中重要養護措施之一,通過整形修剪可以滿足觀賞要求;整形修剪可促進衰老樹、弱枝、彎曲枝更新復壯、加強樹勢,也可削弱過強枝長勢以平衡枝條均衡分布;此外,整形修剪可改善樹冠內通風透光條件[3-4]。目前關于修剪對彩葉樹木光合生理的影響尚無報道,因此,本研究以2年生的盆栽金葉女貞為試驗材料,研究剪梢條件下金葉女貞光合生理特性和色素含量的變化規律,以期為進一步完善金葉女貞的人工管理措施提供參考依據。
1 材料與方法
1.1 試驗材料及處理
試驗于2011年5月至9月在南京農業大學校園內進行。選用生長健壯,長勢相近的2年生盆栽金葉女貞30株[種植于內徑30 cm、深25 cm的花盆中,栽培基質為園土∶基質∶有機肥=1∶1∶1(體積比),每盆1株]。不剪梢(對照)和剪梢處理各15盆,采用常規管理,2011年5月摘除花序。
2011年5月中旬對剪梢處理組進行第1次剪梢處理(去除枝條上部金或淺黃葉),7月上旬和9月上旬分別對新枝條進行同樣的處理。
1.2 方法
分別于2011年5月下旬、7月中下旬和9月中下旬的晴朗天氣,選擇不剪梢和剪梢金葉女貞南向的枝梢,測定中部功能葉、下部老葉光合色素含量、光合作用、葉綠素熒光參數,3次重復。
1.2.1 光合色素含量的測定 葉綠素(Chlorophyll,Chl.)和類胡蘿卜素(Carotinoid,Car.)含量的測定采用張憲政[5]的方法。于8∶00,分別采摘植株中、下部葉位葉片,立即帶回實驗室,去掉主脈,剪碎混勻,稱取0.2 g,用20 ml提取液(丙酮∶無水乙醇=1∶1,體積比)浸提24 h,并用保鮮膜封口防止提取液揮發,浸泡至組織變白后,用紫外/可見光分光光度計測定440 nm、645 nm、663 nm的OD值,葉綠素和類胡蘿卜素含量的計算公式如下:葉綠素a(Ca,mg/L)=12.72A663- 2.59A645,葉綠素 b(Cb,mg/L)=22.88A645- 4.67A663,葉綠素(Ct,mg/L)=Ca+Cb,類胡蘿卜素 (Cc,mg/L)=4.695A440-0.268(Ca+Cb),色素含量=(色素濃度×提取液體積×103)/葉片鮮重,單位為mg/g,重復3次,所有色素含量均以鮮重表示。
1.2.2 光合作用日變化的測定 選擇晴朗無風天氣,自7∶00~17∶00每隔2 h在完全模擬自然環境的條件下,選取完好的功能葉、老葉為測定對象,利用英國PP-System公司生產的CIRAS-1型光合測定系統測定葉片凈光合速率(Pn)、蒸騰速率(Tr)等光合參數,每次重復3次,取平均值。水分利用效率WUE=Pn/Tr(取日平均值),利用AutoCAD軟件計算日變化曲線圍成的面積,得到Pn日積分值(Diurnal integral value of Pn,DIV of Pn)[6]。
1.2.3 葉綠素熒光參數的測定 采用FMS-2便攜脈沖調制式熒光儀(英國Hansatech公司提供),與光合作用日變化同時進行測定葉片葉綠素熒光參數的日變化,測定的指標包括初始熒光(F0)、光系統Ⅱ(PSⅡ)最大光化學效率(Fv/Fm)、光系統Ⅱ(PSⅡ)實際光化學效率(Yield)。葉綠素熒光的光化學淬滅(qp)和非光化學淬滅(NPQ)方面的光能分配參照Quick等[7]提出的方法計算,電子傳遞速率ETR=0.50 ×0.85 × ΦPSⅡ × PAR[8]。各指標每次重復3次,均取日均值。
1.3 數據分析
利用Excel和Systatv7.0統計軟件進行數據處理與分析。采用HSD法(P<0.05)進行差異顯著性分析。
2 結果
2.1 連續剪梢對金葉女貞不同葉位葉片光合色素含量的影響
不剪梢(對照)與剪梢處理金葉女貞不同葉位光合色素含量(表1)顯示:不同時期不剪梢處理(對照)金葉女貞中部葉位葉片葉綠素含量由大到小依次為:2011年9月、2011年7月、2011年5月,不同時期剪梢處理中部葉位葉片葉綠素含量由大到小依次為:2011年7月、2011年9月、2011年5月,不同時期不剪梢(對照)和剪梢處理下部葉位葉片葉綠素含量均由大到小依次為:2011年7月、2011年9月、2011年5月,差異均達到顯著水平(P<0.05)。2011年7月剪梢處理中、下部葉位葉片葉綠素含量均顯著大于不剪梢處理(對照)(P<0.05);2011年9月剪梢下部葉位葉片葉綠素含量顯著大于不剪梢處理(對照)(P<0.05)。2011年5月不剪梢(對照)與剪梢處理金葉女貞中部葉位葉片類胡蘿卜素含量均顯著大于2011年7月、2011年9月(P<0.05);2011年7月不剪梢處理(對照)金葉女貞下部葉位葉片類胡蘿卜素含量均顯著大于2011年7月、2011年9月(P<0.05)。2011年5月剪梢處理下部葉位葉片類胡蘿卜素含量顯著大于不剪梢處理(對照)(P<0.05);2011年7月剪梢處理下部葉位葉片類胡蘿卜素含量小于不剪梢處理(對照)(P>0.05),中部葉位葉片則相反。2011年5月不剪梢(對照)與剪梢處理中、下部葉位葉片葉綠素a/葉綠素b值均顯著高于2011年7月和2011年9月(P<0.05);2011年5月和2011年7月不剪梢處理(對照)中、下部葉位葉片葉綠素a/葉綠素b值均高于2011年9月。2011年7月不剪梢(對照)與剪梢處理金葉女貞下部葉位葉片葉綠素/類胡蘿卜素值均顯著高于2011年5月和2011年9月;2011年7月剪梢處理下部葉位葉片葉綠素/類胡蘿卜素值顯著大于不剪梢處理(對照)(P<0.05)。
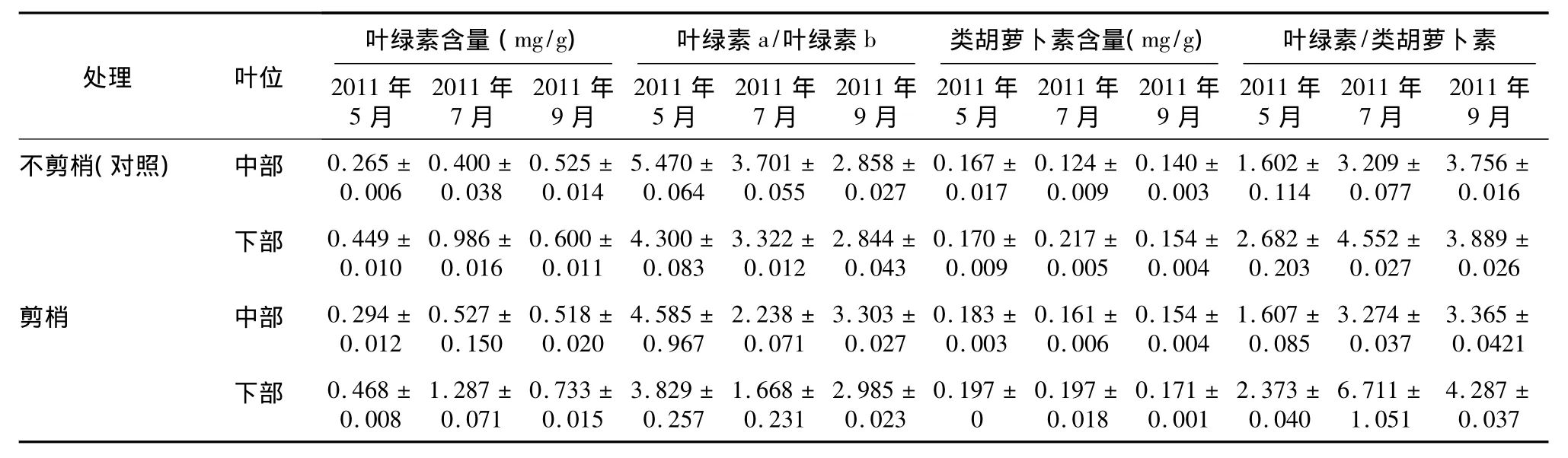
表1 剪梢對金葉女貞不同葉位葉片光合色素含量的影響Table 1 Effects of tip pruning on pigment content in middle and lower leaves of Ligustrum vicaryi
2.2 比較連續剪梢對不同葉位光合特性的影響
2.2.1 凈光合速率(Pn)日變化與凈光合速率日積分值(DIV of Pn) 圖1表明,不同時期不剪梢(對照)與剪梢處理中部葉位葉片凈光合速率日變化均呈雙峰曲線,均在11∶00~13∶00間出現光合午休現象;不同時期不剪梢(對照)與剪梢處理下部葉位葉片凈光合速率日變化均為單峰曲線。2011年7月剪梢處理金葉女貞中、下部葉片凈光合速率均高于不剪梢處理(對照)。
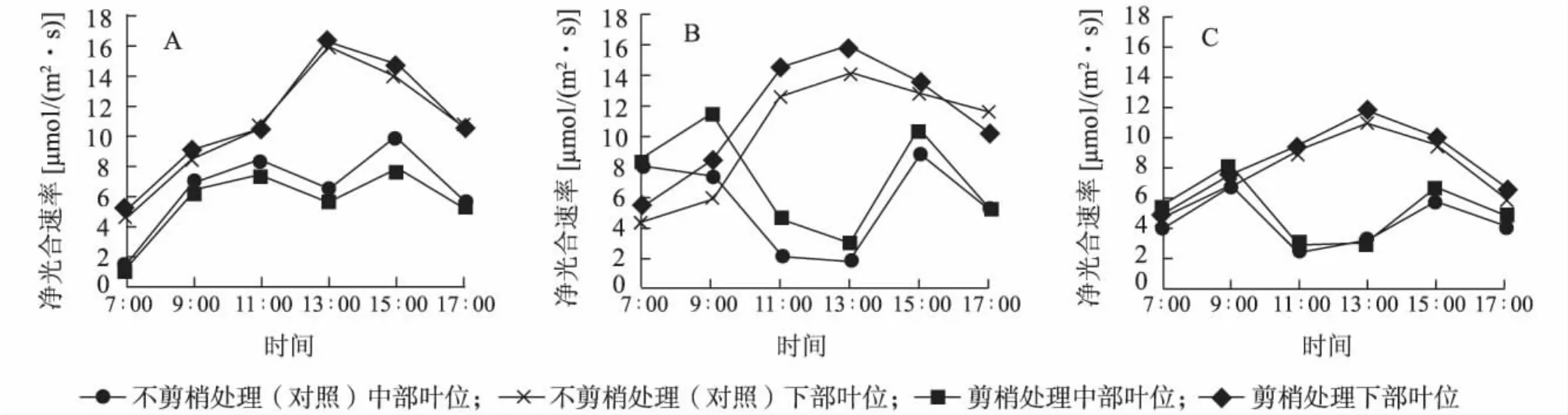
圖1 凈光合速率(Pn)日變化Fig.1 Diurnal variation of Pnin middle and lower leaves with or without pruning
凈光合速率日積分值(DIV of Pn)可以反映一定時間內植物實際的光合凈積累量。如圖2所示,不剪梢處理(對照)金葉女貞中、下部葉位葉片凈光合積累量隨著時間的推進而下降。2011年7月剪梢處理金葉女貞中、下部葉位葉片凈光合積累量大于2011年8月和2011年9月。

圖2 不同葉位葉片凈光合速率日積分值Fig.2 DIV of Pnand WUE of middle and lower leaves with or without pruning
2.2.2 水分利用效率(WUE)日變化 如圖3所示,2011年7月、2011年9月白天上午隨著溫度光照強度的增加,不剪梢處理(對照)葉片水分利用效率降幅大于剪梢處理,自午間開始,二者變化趨勢相近。
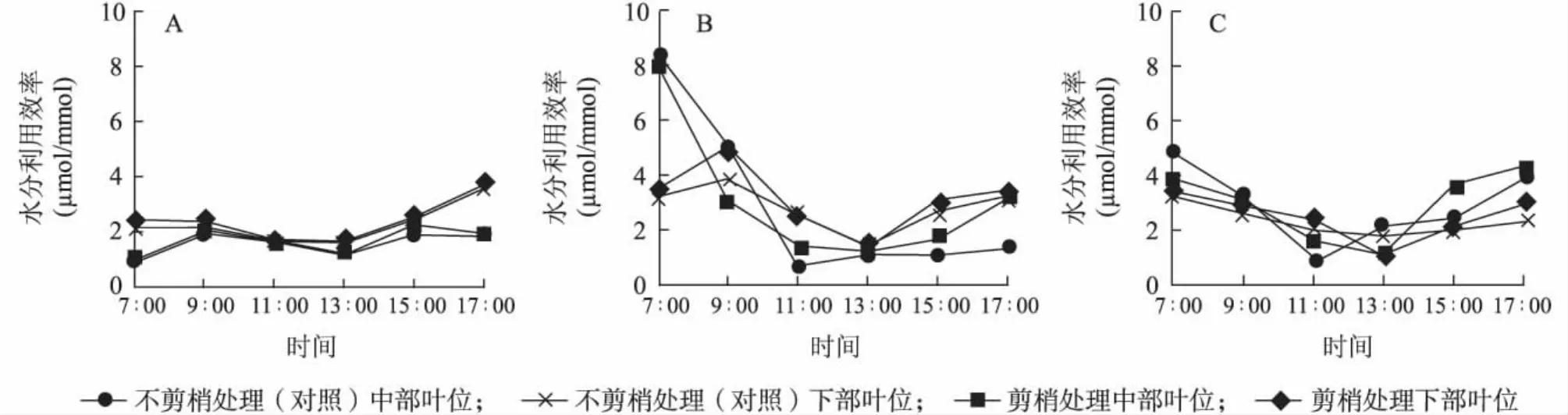
圖3 水分利用效率(WUE)日變化Fig.3 Diurnal variation of WUE in middle and lower leaves with or without pruning
2.2.3 葉綠素熒光參數的比較 由圖4可知,隨著時間的推進,不剪梢處理(對照)中、下部葉位葉片的初始熒光(FO)隨之增加。2011年5月、2011年7月剪梢處理中、下部葉位葉片F0均高于不剪梢處理(對照),2011年9月剪梢處理中、下部葉位葉片F0均低于不剪梢處理(對照)。不剪梢處理(對照)中、下部葉位葉片光系統Ⅱ(PSⅡ)最大光化學效率(Fv/Fm)變化趨勢與F0一致。2011年5月、2011年7月剪梢處理中部葉位葉片Fv/Fm大于不剪梢處理(對照)(P<0.05)。2011年7月、2011年9月剪梢處理中部葉位葉片PSⅡ實際光化學效率(Yield)顯著小于不剪梢處理(對照)(P<0.05),2011年9月剪梢處理下部葉位葉片Yield顯著小于不剪梢處理(對照)(P<0.05)。不同時期剪梢處理下部葉位葉片qp均小于不剪梢處理(對照)。2011年7月、2011年9月剪梢處理中、下部葉位葉片非光化學猝滅(NPQ)均顯著高于不剪梢處理(對照)(P<0.05)。不同時期剪梢處理中、下部葉位葉片電子傳遞速率(ETR)均低于不剪梢處理(對照)。
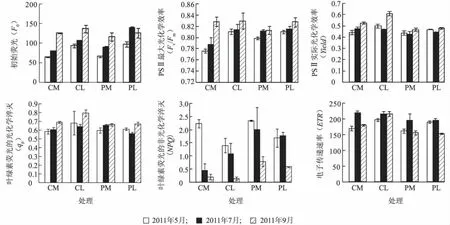
圖4 不同葉位初始熒光(Fo)、PSⅡ最大光化學效率(Fv/Fm)、PSⅡ實際光化學效率(Yield)、電子傳遞速率(ETR)、葉綠素熒光的光化學淬滅(qp)和葉綠素熒光的非光化學淬滅(NPQ)Fig.4 Fo,Fv/Fm,Yield,ETR,qpand NPQ of middle and lower leaves with or without pruning
3 討論
3.1 連續剪梢對金葉女貞光合特性的影響
前人研究認為,植物葉片在強光下難以全部消耗過剩光能,這些光能將產生光抑制,不利于植物葉綠素的生成,還會使其分解[9-10]。本研究結果顯示,2011年9月剪梢處理中部葉片葉綠素含量小于不剪梢處理(對照),但2011年5月、2011年7月剪梢處理中、下部葉位葉片葉綠素含量均大于不剪梢處理(對照),這可能是因為剪梢改善中、下部葉位葉片的光照條件,同時也消除植株頂端優勢,從而使物質分配發生轉移,為葉綠素的合成提供了更多的底物。葉綠素含量的增加利于光合作用的進行[11-12]。本研究結果2011年7月剪梢處理金葉女貞中、下部葉片凈光合速率均高于不剪梢處理(對照)。此外,2011年5月剪梢處理的中部葉位葉片光合速率降低,可能是中部葉位光合物質用于修復損傷[13]。
夏季植物接收太陽光能進行光合作用的同時也遭受強太陽輻射的脅迫,葉片光合機構吸收過剩光能后會引起開放PSII反應中心最大光化學效率(Fv/Fm)的降低,即產生光抑制現象,嚴重的光抑制可以導致反應中心的不可逆破壞[14]。2011年7月剪梢處理中部葉位葉片Fv/Fm和光化學猝滅系數(qp)均高于不剪梢處理(對照),這表明PSII反應中心的能量捕捉效率和轉換效率增加,電子傳遞活性增大。2011年7月剪梢處理中部葉位葉片PSⅡ實際光化學效率(Yield)和電子傳遞速率(ETR)小于不剪梢處理(對照)。2011年7月剪梢和不剪梢(對照)處理凈光合速率及葉綠素熒光參數(Fv/Fm、qp)結果表明,午間剪梢處理金葉女貞中部葉位葉片電子傳遞受抑制相對嚴重,加劇PSII反應中心的光抑制程度。
高等植物在系統發育過程中形成了對光合機構起重要保護作用的生理生化機制,這種機制依賴葉黃素循環和PSII反應中心可逆失活與熱能耗散[15-16]。非光化學猝滅(NPQ)反映植物通過熱耗散消耗過剩激能實現自身光保護的能力。本研究結果表明,連續剪梢處理在改善光合作用的同時激發了葉片光合機構的高速運轉,更過多的過剩光能以熱耗散形式散發,從而減輕傷害,這與前人的研究結果一致[17-19]。
3.2 連續剪梢對金葉女貞葉片光合色素含量的影響
金葉植物葉片在強光下葉色金黃,弱光下由于葉綠素含量增加,葉色轉變為黃綠[20]。本研究結果表明,連續剪梢處理有利于金葉女貞中、下部葉位葉片類胡蘿卜素的合成。2011年5月、2011年7月剪梢處理金葉女貞中、下部葉位葉片葉綠素/類胡蘿卜素值高于不剪梢處理。表明完全剪除上部金黃葉或淺黃葉不利于金葉女貞葉片色彩的表達。
綜上,2011年5月、2011年7月連續剪梢處理可促進金葉女貞葉片葉綠素的合成,有利于葉片光合作用,但2011年7月剪梢處理金葉女貞中部葉位葉片PSII反應中心午間的電子傳遞受阻更嚴重,光抑制程度加重。此外,完全去除上部金黃葉或淺黃葉不利于金葉女貞觀賞價值的實現。因此,在園林綠化栽培修剪管理中,應該采取輕度修剪,保留部分金黃葉或淺黃葉以平衡其生長發育與觀賞特性。
[1]姜衛兵,莊 猛,花國平,等.彩葉植物呈色機理及光合特性研究進展[J].園藝學報,2005,32(2):352-358.
[2]廖 容,崔 潔,卓春麗,等.成都市32種立體綠化植物降溫增濕效應比較研究[J].江蘇農業科學,2012,40(6):178-182.
[3]高文海,薛純紅.樹木整形修剪的原因和季節[J].黑龍江科技信息,2009(7):269.
[4]黃冬賢,黎可華,黎明星,等.紫薇的整形修剪及園林應用[J].廣東園林,2010(4):67-69.
[5]張憲政.作物生理研究法[M].北京:中國農業出版社,1986.
[6]莊 猛,姜衛兵,李志國.金邊黃楊和大葉黃楊光合特性的比較[J].植物生理學通訊,2006,42(1):39-42.
[7]QUICK W P,STITT M.An examination of factors contributing to non-photochemical quenching of chlorophyll fluorescence in barley leaves[J].Biochimica and Biophysica Acta,1989,977:287-296.
[8]MAXWELLA K.JOHNSON G N.Chlorophyll fluorescence-a practical guide[J].Journal of Experimental Botany,2000,51(345):659-668.
[9]陸定志,傅家瑞,宋松泉.植物衰老及其調控[M].北京:中國農業出版社.1997.
[10]YANG X H,ZOU Q,WANG W.Photoinhibition in shaded cotton leaves after exposing to light and the time course of its restoration[J].Acta Botanica Sinica,2001,43:1255-1259.
[11]崔培強,姜衛兵,翁忙玲,等.遮蔭對紫葉李幼苗葉片色素含量及光合速率的影響[J].西北植物學報,2010,30(11):2286-2292.
[12]姜 武,姜衛兵,李志國.園藝作物光合性狀種質差異及遺傳表現研究進展[J].經濟林研究,2007,25(4):102-108.
[13]高桐梅,衛雙玲,張海洋,等.打頂對芝麻不同葉位光合特性的影響[J].中國油料作物學報,2009,31(4):492-498.
[14]尤 鑫,龔吉蕊,段慶偉,等.兩種雜交楊品系光合系統II葉綠素熒光特征[J].生態學報,2008,28(11):5641-5648.
[15]謝智華,姜衛兵,韓 鍵,等.葉片花色素苷對植物光合作用影響的研究進展[J].植物生理學報,2011,47(6):545-550.
[16]DEMMIG B,WINTER K,KROGER A,et al.Photoinhibit ion and zeaxanthin formation in int act leaves:A possible role of the xantho2 phyll cycle in the dissipat ion of excess light energy[J].Pl ant Physiol,1987,84:218-224.
[17]楊興洪,鄒 琦,趙世杰.遮蔭和全光下生長的棉花光合作用和葉綠素熒光特征[J].植物生態學報,2005,29(1):8-15.
[18]YANG X,YI F,PRASAD P.Response of photosynthesis and chlorophyll fluorescence quenching to leaf dichotocarpism in Ligustrum vicaryi[J].Photosynthetica,2009,47(1):137-140.
[19]YANG X H,ZOU Q,WANG W.Photoinhibition in shaded cotton leaves after exposing to light and the time course of its restoration[J].Acta Botanica Sinica,2001(43):1255-1259.
[20]胡海姿,張 睿,尚愛芹,等.金葉植物色素含量對光強的響應[J].園藝學報,2007,34(3):717-722.
