耐鹽柳樹BADH基因克隆及表達分析
2013-08-02 00:52:10王奎山李玉娟
江蘇農業學報 2013年3期
李 敏, 張 健, 王奎山, 李玉娟, 談 峰, 王 瑩
(江蘇沿江地區農業科學研究所,江蘇 如皋 226541)
大多數高等植物在受到鹽、干旱等脅迫時會在細胞內積累大量甘氨酸甜菜堿(簡稱甜菜堿)來提高自身抵抗逆境環境因子脅迫的能力[1]。甜菜堿作為季胺類水溶性生物堿,具有滲透調節功能,能夠保護質膜的完整性,有助于植物適應非生物逆境[2-3]。
耐鹽柳樹,具有較強的耐鹽能力,能在含鹽0.4%、pH 10.4的土壤中正常生長,是中國沿海灘涂主要的造林樹種之一。耐鹽柳樹適應性強,易繁殖,造林成活率高,生長迅速,無論在營造工業用材林,還是在防風固沙、水土保持、鹽堿地改造等方面都有廣闊的應用前景[4]。目前,國內外學者已從川芎(Liqusticum churning Franch Hort)、枸杞(Lycium barbarum)、刺五加(Eleutherococcus senticosus)、甘菊(Dendranthema lavandulifolium)、胡楊(Populus euphratica)、遼寧堿蓬 (Suaeda liaotungensis)、梭 梭(Haloxy lonammodendron)、鹽爪爪(Kalidium foliatum)、大麥(Hordeum vulgare L.)、鹽穗木 (Halostachys caspica)等[5-14]植物中相繼克隆得到甜菜堿醛脫氫酶(BADH)基因,但有關耐鹽柳樹BADH基因的研究還未見報道。本研究擬從耐鹽柳樹中克隆BADH基因,探討其與其他品種或物種同源基因的相似性,構建系統進化樹,并對其進行序列分析和在鹽脅迫下的表達分析,旨在為耐鹽柳樹種質改良研究奠定基礎。
1 材料與方法
1.1 植物材料
供試植物為江蘇沿江地區農業科學研究所選育的耐鹽柳樹L0911,取新鮮葉片,用液氮速凍存于-70℃冰箱待用。
1.2 試劑
引物 Taq DNA聚合酶、dNTP(10 mmol/L)、DNA marker和 PCR產物純化回收試劑盒(生工SK1141)、質粒提取試劑盒(生工 SK1191 UNIQ-10)、熒光定量PCR用試劑盒(SK8661)、常規化學試劑等均購自上海生工生物工程有限公司,pMD18-T載體購自寶生物工程(大連)有限公司。
1.3 RNA提取和總cDNA的合成
采用Trizol試劑(Invitrogen公司)參照試劑說明手冊提取總RNA。總cDNA的合成用AMV逆轉錄酶(BBI公司)按照反轉錄試劑盒說明書進行。
1.4 BADH 基因克隆
根據毛果楊(Populus trichocarpa)的PtBADH設計引物BADH F1:5'-AGCTCTCAACTCACTAATCGAGT-3';BADH R1:5'-TTATAGCTTGGCGGGAGACTG-3'。以L0911葉片總cNDA為模板,PCR產物經1%瓊脂糖電泳后,切膠回收,并進行T/A克隆(pMD18-T)。陽性克隆送上海生工生物工程有限公司測序。所得的克隆序列為2次獨立PCR和3個以上質粒的測序結果。
1.5 BADH基因的分子進化分析
在GenBank查找BADH基因的核苷酸和氨基酸相關序列,使用 Clustal W2進行多序列比對,MEGA5.0軟件中 Neighbor-Joining(NJ)法進行建樹,Bootstrap值設為1 000。
1.6 鹽處理
將水培生長健壯的柳樹L0911幼苗用200 mmol/L NaCl溶液分別處理0 h、3 h、6 h、9 h、12 h、24 h、48 h、72 h后,取新鮮嫩葉片。將 L0911幼苗分別用 0 mmol/L、100 mmol/L、200 mmol/L 、300 mmol/L NaCl溶液處理12 h后,取新鮮嫩葉片。所有樣品用液氮速凍存于-70℃冰箱待用。
1.7 實時熒光定量PCR分析
根據測序結果,設計引物BADH F2(5'-GTGCCAAGTATTTGCGTGCTA-3')和BADH R2(5'-AAACAACCTGCGACATCATCC-3'),對L0911 BADH基因鹽脅迫處理后的響應進行分析。用ubiquitin-like(UBQ-L)[15]為內參基因,引物為 UBQ-L-F(5'-TGAGGCTTAGGGGAGGAACT-3')和UBQ-L-R(5'-T GTAGTCGCGAGCTGTCTTG-3')。將經過鹽處理的柳樹L0911幼苗嫩葉進行總RNA提取以及cDNA合成。按柱式植物總RNA抽提純化試劑盒(SK8661)配制PCR體系,于ABI Stepone plus型熒光定量PCR儀中進行定量PCR檢測,測定其Ct值,用 F=2-△△Ct公式[16]計算樣品間目的基因的表達量差異。
2 結果
2.1 柳樹L0911 BADH基因的克隆
從圖1可以看出,引物BADH F1和BADH R1在柳樹L0911的cDNA模板中成功擴增到目的片段,長度大小符合預期目標。測序結果表明,L0911的BADH基因cDNA全長1 539 bp,具有起始密碼子、完整的開放閱讀框(1 512 bp)、終止密碼子、5'非編碼區(27 bp),共編碼512個氨基酸。獲得的氨基酸序列與毛果楊PtBADH2、胡楊PeBADH2編碼序列以及擬南芥、水稻的氨基酸序列相似性達到95.83%、95.03%、82.31%、71.68%。說明本試驗獲得的基因與PtBADH2和PeBADH2是同源的。
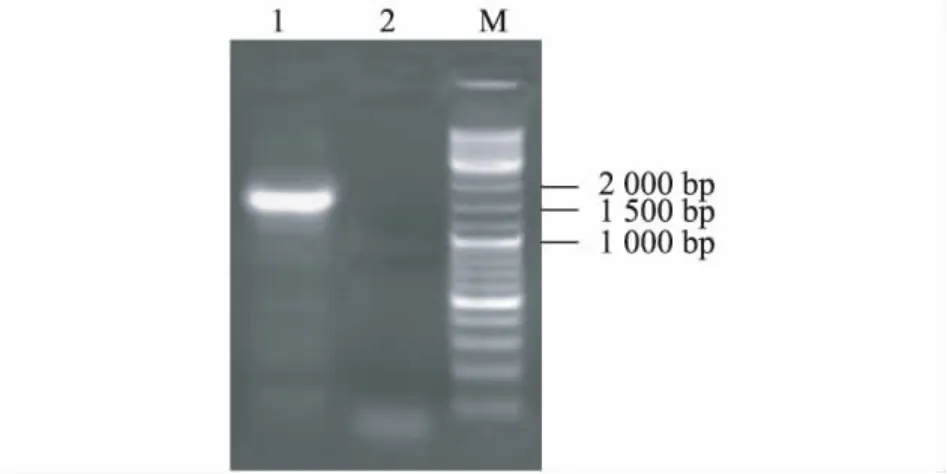
圖1 引物BADH-F1/R1擴增柳樹L0911 cDNA的瓊脂糖凝膠電泳檢測結果Fig.1 Amplification of Salix cultivar L0911 cDNA by PCR using primers BADH-F1/R1
2.2 柳樹L0911氨基酸序列分析和系統進化樹構建
用MEGA 5.0對BADH基因編碼的氨基酸序列進行分子進化分析,結果與用基因組序列(www.phytozome.net)分析結果一致。柳樹 L0911的BADH基因與楊樹位于相同的分支(圖2)。在胡楊[9]、白骨壤[17]、麻瘋樹[18]等植物中,BADH 基因的功能已有報道,本研究獲得的BADH可能具有相應的生物學功能。

圖2 植物BADH基因編碼氨基酸的分子進化分析圖Fig.2 Phylogentic tree of BADH gene of various plants based on amino acids sequence
2.3 鹽脅迫下柳樹L0911 BADH基因的表達分析
2.3.1 不同濃度 NaCl溶液脅迫下柳樹L0911BADH基因的表達特性 以未進行NaCl脅迫處理的葉片中BADH基因的表達量為對照,對實時熒光定量中得到的目的基因BADH和內參基因UBQ-L的平均Ct值,計算作圖,得到圖3。可以看出柳樹L0911葉片中BADH基因的表達情況隨著鹽濃度的升高呈先升高后降低的趨勢,在200 mmol/L NaCl脅迫下表達量最高,是對照的1.90倍。說明一定濃度的 NaCl脅迫可以誘導柳樹 L0911的BADH基因表達量增加,但鹽濃度過高BADH基因表達量又會下降。

圖3 不同NaCl濃度脅迫12 h后柳樹L0911 BADH基因的表達Fig.3 Expression of BADH gene of Salix cultivar L0911 under NaCl stress for 12 h at different concentrations
2.3.2 NaCl不同脅迫時長下 L0911BADH基因的表達特性 從圖4可以看出,隨NaCl脅迫時間的延長,葉片中BADH基因表達量呈現先升高后降低的趨勢。脅迫初期(3 h),L0911 BADH基因表達量低于對照;隨著時間的延長,表達量逐漸增大,脅迫12 h時達到最高,是對照組的1.88倍,說明鹽脅迫能誘導該基因表達量的增加;隨著時間進一步延長,表達量逐漸降低,脅迫72 h時表達量低于對照水平。
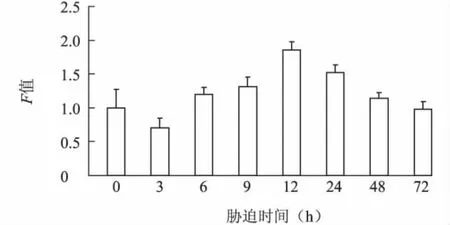
圖4 200 mmol/LNaCl脅迫不同時間柳樹L0911 BADH基因的表達Fig.4 Expression of BADH gene of Salix cultivar L0911 under 200 mmol/L NaCl stress for different times duration
3 討論
植物在干旱、鹽堿等逆境條件下可積累大量甜菜堿,而植物體內甜菜堿的合成途徑相對簡單,主要經兩步催化合成,即乙酰膽堿→甘氨酸甜菜堿醛→甘氨酸[19]。甜菜堿合成后不能進一步代謝,積累在細胞中作為永久性或半永久性滲透調節劑[20]。因此,甜菜堿合成途徑中的甜菜堿醛脫氫酶(BADH)被認為是最有希望的脅迫抗性基因之一,BADH的分子生物學與生物化學研究引起了廣泛關注。本研究從柳樹L0911克隆到BADH基因,并對該基因的結構、系統進化、表達模式進行了比較詳細的研究。
本研究采用同源克隆法從柳樹L0911cDNA中擴增并克隆了BADH基因。測序結果表明,該基因含有1個完整的開放閱讀框架,核苷酸序列全長為1 539 bp,推測的氨基酸序列全長為512個氨基酸殘基。氨基酸序列同源性分析表明L0911的BADH基因與毛果楊、胡楊等同源性最近,為95%,與水稻的同源性為71%。L0911的BADH氨基酸序列同源性分析也表明BADH在高等植物中是相對保守的。
植物在逆境條件下會積累甜菜堿,BADH基因的表達活性又直接影響甜菜堿的合成。本試驗用熒光定量的方法檢測柳樹L0911在鹽脅迫下BADH mRNA的表達情況。結果表明,BADH基因是受鹽誘導的,當鹽濃度升高至200 mmol/L時,表達活性最大;在同一鹽濃度下,隨著鹽脅迫時間的延長,在一定范圍內,表達活性也增大。Arakawa等[21]研究結果表明大麥(Hordeum vulgare)BADH酶活性隨著鹽濃度的增加而增大;Reda等[22]研究結果也表明鹽角草(Salicornia europaea)和堿蓬(Suaeda maritima)的BADH mRNA在鹽脅迫下的表達水平增加,本研究結果與他們基本一致。這為進一步揭示L0911的耐鹽機制及利用該基因進行遺傳工程創造耐鹽新種質奠定了基礎。
[1]CHEN T H H,MURATA N.Glycinebetaine:an effective protectant against abiotic stress in plants[J].Trends Plant Sci,2008,13(9):499-505.
[2]朱先燦,胡鳶雷,王曉麗,等.兩種濱藜甜菜堿醛脫氫酶基因的克隆及序列分析[J].生物技術通報,2007(6):89-91.
[3]劉 萍,丁義峰,于 娜,等.維生素C與甘氨酸甜菜堿對芍藥花瓣生理生化的影響[J].江蘇農業科學,2011,39(6):267-269.
[4]解孝滿,李景濤,劉建軍,等.柳樹無性系苗期試驗分析[J].山東林業科技,2008(3):44-45.
[5]周嘉裕,廖 海.川芎甜菜堿醛脫氫酶基因的克隆及序列分析[J].西北農業學報,2011,20(3):45-51.
[6]田躍勝,許潔婷,唐克軒,等.枸杞甜菜堿醛脫氫酶基因全長cDNA的克隆與表達分析[J].揚州大學學報,2010,31(2):48-52.
[7]邢朝斌,吳 鵬,陳 龍,等.刺五加GAPDH基因的克隆及序列分析[J].中草藥,2012,43(1):155-158.
[8]劉振林,曹華雯,夏新莉,等.甘菊BADH基因cDNA的克隆及在鹽脅迫下的表達[J].武漢植物學研究,2009,27(1):1-7.
[9]劉佳琪,楊 雪,李 迪,等.胡楊甜菜堿醛脫氫酶基因的功能分化[J].生物工程學報,2012,28(3):329-339.
[10]李秋莉,張 毅,尹 輝,等.遼寧堿蓬甜菜堿醛脫氫酶基因(BADH)啟動子分離及序列分析[J].生物工程學報,2006,22(1):77-81.
[11]石 磊,甘曉燕,陳虞超,等.梭梭甜菜堿醛脫氫酶基因克隆及序列分析[J].西北植物學報,2010,30(2):223-228.
[12]曾幼玲,幸 婷,蔡忠貞,等.鹽生植物鹽爪爪甜菜堿醛脫氫酶基因的克隆及在鹽脅迫下的BADH基因的表達[J].云南植物研究,2007,29(1):79-84.
[13]ZHAO Y W,HAO J G,BU H Y,et al.Cloning of HvBADH gene from hulless barley and its transformation to tobacco[J].Acta Agronomica Sinica,2008,34(7):1153-1159.
[14]胡有貞,張富春,曾幼玲,等.鹽穗木甜菜堿醛脫氫酶基因(BADH)的克隆及其在鹽脅迫下的表達分析[J].植物生理學通訊,2010,46(1):47-50.
[15]關亞麗.柳樹AP3同源基因的克隆與表達分析[D].南京:南京林業大學,2006.
[16]LIVAK K J,SCHMITTGEN T D.Analysis of relative gene expression data using real-time quantitative PCR and the 2-△△Ctmethod[J].Method,2001,25:402-408.
[17]周涵韜,李 芳,張賽群,等.紅樹植物白骨壤甜菜堿醛脫氫酶基因的克隆與功能研究[J].廈門大學學報,2008,47(增刊2):11-15.
[18]ZHANG F L,NIU B,WANG Y C,et al.A novel betaine aldehyde dehydrogenase gene from Jatropha curcas,encoding an enzyme implicated in adaptation to environmental stress[J].Plant Science,2008,174:510-518.
[19]TAKABE T,RAI V,HIBINO T.Metabolic engineeringof glycine betaine[M]//RAI A K,TAKABE T.Abiotic stress tolerance in plants.Berlin:Springer,2006:137-151.
[20]KUMAR S,DHINGRA A,DANIELL H.Plastid-expressed betaine aldehyde dehydrogenase gene in carrot cultured cells,roots,and leaves confers enhanced salt tolerance[J].Plant Physiol,2004,136(1):2843-2854.
[21]ARAKAWA K,KATAYAMA M.Levels of betaine and betaine aldehyde dehydrogenase activity in the green leaves,and etiolated leaves and roots of barley[J].Plant Cell Physiol,1990,31:797-803.
[22]MOGHAIEB R E A,SANEOKA S,FUJITA K.Effect of salinity on osmotic adjustment,glycinebet aine accumulation and the betaine aldehyde dehydrogenase gene expression in two halophytic plants,Salicornia europaea and Suaeda maritime[J].Plant Sci,2004,166:1345-1349.
猜你喜歡
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
山東工業技術(2016年15期)2016-12-01 05:31:22
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
