產褐藻膠裂解酶菌株的分離與鑒定
2013-08-07 09:23:18黃李淑馨曹海龍譚海東趙小明杜昱光
食品工業科技 2013年23期
黃李淑馨,劉 鋼,岳 敏,曹海龍,譚海東,趙小明,尹 恒,* ,杜昱光,*
(1.中國科學院大連化學物理研究所,遼寧大連116023;2.中國科學院大學,北京100049;3.大連海洋大學食品科學與工程學院,遼寧大連116023)
褐藻膠是由β-D-甘露糖醛酸(mannuronate,M)和α-L-古洛糖醛酸(guluronate,G)組成的直鏈多糖。兩種糖醛酸單體通過α/β-1,4 糖苷鍵隨機排列成褐藻膠分子中的聚甘露糖醛酸片段(polyMblock),聚古洛糖醛酸片段(polyG-block)以及MG 交替嵌段(polyMG-block)[1]。褐藻膠結構復雜,物化性質多樣,因而它被廣泛應用于食品、醫藥、化妝品等領域[2]。此外,褐藻膠寡糖因具有免疫調節[3]和神經保護[4]等生物活性,所以它可被開發成獨特療效藥物或功能性食品。近年來,隨著對褐藻膠及其寡糖的功能評價和開發研究的不斷深入,“量身定制”(tailor-made)的新理念被提出,即指設計具有確定結構和聚合度的褐藻膠及其寡糖,使其物化性質或生物活性滿足特定用途的需要[5]。褐藻膠裂解酶是一類通過β-消去反應斷裂褐藻膠的裂解酶,因其具有專一、高效及反應溫和等優點,被視為改造褐藻膠的有力工具。褐藻膠裂解酶按底物專一性可分為聚甘露糖醛酸片段裂解酶(EC 4.2.2.3)和聚古洛糖醛酸片段裂解酶(EC 4.2.2.11),也可按反應模式分為內切或外切酶[6]。褐藻膠裂解酶還可用于降解菌膜[7]及生產生物燃料[8]。受底物專一性和反應模式的影響,單一類型的酶能斷裂的褐藻膠位點有限。因此,篩選褐藻膠代謝新菌株,尋找不同類型的酶對開發高值化褐藻膠及其寡糖產品具有重要意義。本研究從海南采集的多類樣品中篩到高產褐藻膠裂解酶的4株菌,然后研究了其產酶規律,并通過形態學和系統發育分析對其進行鑒定,旨在獲得新酶源微生物資源,為酶的克隆表達及酶法改造褐藻膠奠定基礎。

表1 產褐藻膠裂解酶菌株的來源Table 1 Sources of 10 alginate lyase-producing strains
1 材料與方法
1.1 材料與儀器
供分離的37 個樣品(包括土壤、淤泥、淡水和海水) 分別采自海南省的風爐嶺、紅樹林、玉帶灘、七仙嶺、五指山、天涯海角、黎安鎮漁港及海口市周邊水產海藻工廠等地區;無機營養鹽溶液(1000mL):NaCl 5g,KH2PO41g,MgSO4·7H2O 0.5g,CaCl20.2g,FeSO4·7H2O 0.02g;分離培養基(1000mL):無機營養鹽溶液中加入褐藻酸鈉7g,(NH4)2SO45g,瓊脂20g,pH7.2;產酶發酵培養基(1000mL):無機營養鹽溶液中加入褐藻酸鈉10g,蛋白胨5g,酵母粉1g,pH7.2;種子培養基同發酵培養基;褐藻酸鈉 美國Sigma 公司;其他試劑均為國產分析純;細菌基因組DNA 提取試劑盒及PCR 反應所用的相關試劑 均購自寶生物工程(大連)有限公司;凝膠回收試劑盒 北京博邁德科技發展有限公司。
LRH 系列生活培養箱、HWS24 型電熱恒溫水浴鍋 上海一恒科學儀器有限公司;全溫振蕩培養箱 太倉市華美生化儀器廠;Veriti 型PCR 儀 美國ABI 公司;GelDoc-It 型凝膠成像儀 美國UVP 公司;Thermo Multiskan Ascent 酶標儀 芬蘭Labsystems 公司;IX73型顯微鏡 日本Olympus 公司。
1.2 實驗方法
1.2.1 菌株的分離 稱取土壤或淤泥10g,放入盛有90mL 無菌水及玻璃珠的三角瓶中,220r/min 振蕩15min 使樣品充分分散,梯度稀釋后涂布于含有褐藻酸鈉的分離培養基平板。30℃倒置培養2~3d,挑取生長良好、形態不一的菌落進行多次劃線純化,并結合顯微鏡觀察得到純菌株。海水和淡水樣品直接進行梯度稀釋后涂布分離培養基平板。
1.2.2 產褐藻膠裂解酶菌株的初篩 將分離得到的菌株挑取一環,接種于50mL 產酶發酵培養基,30℃、150r/min 條件下培養,觀察培養基粘度變化。培養4d 后取發酵上清液檢測是否有酶活。
1.2.3 產褐藻膠裂解酶菌株的復篩 將初篩獲得的產酶菌株挑取一環,接種于25mL 種子培養基,30℃、150r/min 搖床培養24h。種子液以2%的接種量轉接于50mL 產酶發酵培養基,30℃、150r/min 培養。分別于1 ~6d 取樣,將發酵液12000r/min 離心10min,取上清液作為粗酶液測酶活。
1.2.4 褐藻膠裂解酶活力的測定 酶活力測定采用3,5-二硝基水楊酸法(DNS)[9],以0.5%(w/v)褐藻酸鈉為底物,測定發酵上清液中的褐藻膠裂解酶活力。酶活力單位定義為:每分鐘釋放1μmol 還原糖(以葡萄糖計)所需的酶量為一個酶活力單位(U)。發酵液酶活力單位定義為每毫升發酵液含酶活單位(U/mL)。
1.2.5 16S rDNA 序列測序與系統發育樹的構建 將目的菌株接種于種子培養基中,30℃、150r/min 搖床培養20h,12000r/min 離心10min 收集菌體,按試劑盒操作步驟提取菌株的基因組DNA。采用通用引物27f(5'-AGAGTTTGATCM[C:A]TGGCTCAG-3')和1518r(5'-AAGGAGGTGATCCAN[A:C:G:T]CCR[A:G]CA-3')[10],以菌株基因組DNA 為模板進行16S rDNA 的PCR 擴增。PCR 反應條件為:95℃5min;94℃1min,55℃1min,72℃1.5min,30 個循環;72℃10min。PCR 擴增產物回收純化后由寶生物工程(大連)有限公司測序。
將16S rDNA 測序結果在NCBI 核苷酸數據庫中進行BLAST 檢索(http://blast.ncbi.nlm.nih.gov/),選取同源性較高菌株的16S rDNA 序列,采用MEGA5.1軟件選擇模型,然后用BEAST v1.7.5 軟件構建系統發育樹[11-12]。
2 結果與分析
2.1 產酶菌株篩選
采用褐藻酸鈉為唯一碳源的分離平板從海南樣品中分離出菌株,然后將菌株接入發酵培養基進行初篩。初篩基于的原理是褐藻酸鈉溶于水能形成粘稠狀溶液,若菌株在含有褐藻酸鈉的粘稠培養基中生長并使其粘度下降,說明菌株可以分解褐藻酸鈉[13];同時若該菌株的培養基上清液用DNS 法檢測到酶活,則進一步確認該菌株產褐藻膠裂解酶。采用雙重指標進行初篩,能有效避免假陽性結果,該篩選模型的建立為今后的實驗者提供了良好的參考依據。
通過初篩,獲得了具有產褐藻膠裂解酶能力的10 個菌株,它們的來源見表1。從表1 可以看出,產褐藻膠裂解酶的菌群分布范圍較寬,它們不僅存在于海水、腐爛海藻中,還存在于火山砂土、淡水河流泥沙中。
將初篩得到的10 株菌經種子培養后,轉接于發酵培養基中,分別于第24、48、72、96、120、144h 檢測發酵上清液中酶活。通過復篩,獲得4 株具有較好產酶能力的菌株,編號分別為W14、S20、E03 和W13,它們的發酵液酶活隨培養時間的變化如圖1 所示。由圖1 可以看出,4 株菌中菌株E03 的發酵液酶活最高(0.58U/mL),并且在培養48~72h 內其酶活呈現陡增態勢,72h 后發酵液中酶活逐步下降。菌株S20 和W14 的最高發酵液酶活相當(0.23~0.27U/mL),但其產酶高峰期不同。菌株S20 在培養前期(24~48h)迅速達到產酶高峰,而此階段W14 的發酵液檢測不到酶活,直至培養中期(48~96h)W14 產酶顯著增加至高峰。菌株W13 的發酵液酶活稍低(0.13U/mL),但其在培養72h 內產酶平穩。
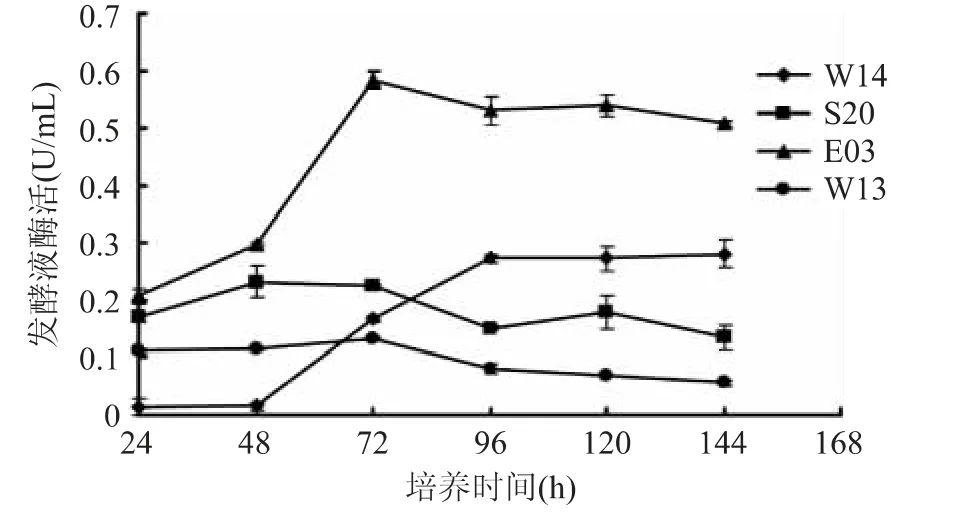
圖1 褐藻膠裂解酶活性變化曲線Fig.1 Activition curve of alginate lyases
2.2 菌落及菌株形態觀察
菌株E03 單個菌落呈淡粉色半透明,圓形,隆起,表面光滑,邊緣整齊,菌苔粘稠,容易形成融合狀態菌落(圖2A),革蘭氏染色陰性,菌體呈球桿狀,可見單個散生或形成兩個聚集(圖2E)。菌株S20 單個菌落呈橙黃色不透明,圓形,隆起,表面光滑,邊緣整齊(圖2B),革蘭氏染色陰性,菌體為桿狀,可見單個散生(圖2F)。菌株W13 單個菌落呈白色半透明,圓形,隆起,表面光滑,邊緣整齊(圖2C),革蘭氏染色陰性,菌體為桿狀,兩端鈍園,可見單個散生或形成多個聚集(圖2G)。菌株W14 單個菌落呈白色不透明,圓形,隆起,表面光滑,邊緣整齊,菌苔粘稠,容易形成融合狀態菌落(圖2D),革蘭氏染色陰性,菌體為桿狀,可見單個散生或形成2~3 個聚集(圖2H)。
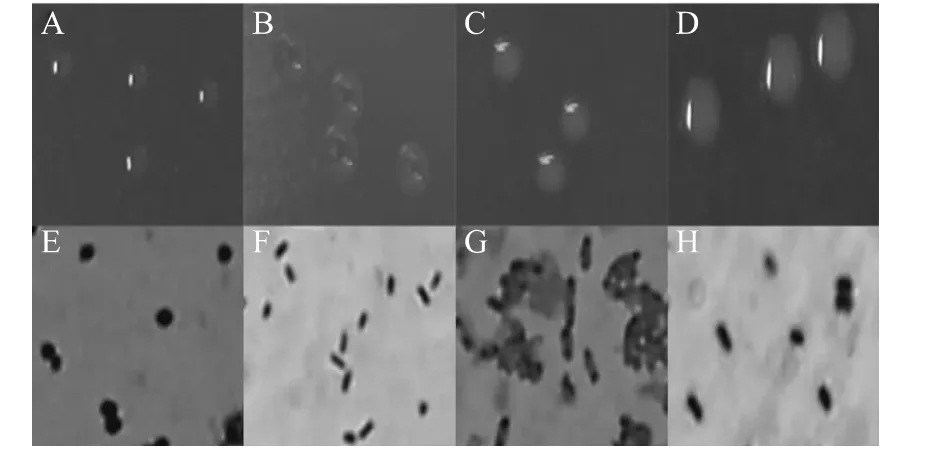
圖2 菌株E03、S20、W13 和W14 的菌落形態和顯微形態(×1000)Fig.2 The colonial morphology and micrograph of four bacteria strains
2.3 16S rDNA 序列分析與系統發育樹的構建
菌株E03、S20、W13 和W14 的16S rDNA 的PCR擴增產物大小均為1460bp 左右(圖3),經測序后提交至 GenBank (登錄號:KC777292、JQ886462、KC777293 和KC777294)。將4 個菌株的16S rDNA序列在NCBI 數據庫進行BLAST 比對,然后分別選取同源性較高菌株的16S rDNA 序列,采用BEAST 軟件構建了4 個菌株的系統發育樹(圖4)。從圖4 上可以看出,發育樹分為兩大聚類分支,它們分別從屬于擬 桿 菌 門 (Bacteroidetes ) 和 變 形 菌 門(Proteobacteria)。黃桿菌屬(Flavobacterium)屬于擬桿菌門,假單胞菌屬(Pseudomonas)和弧菌屬(Vibrio)均屬于變形菌門。據推測,約25 億年前變形菌門與其它門從共同祖先發生分歧[14],因此,在圖4 的發育樹上,假單胞菌屬和弧菌屬聚類的大分支在進化的較早時期就與黃桿菌屬分開。假單胞菌屬和弧菌分屬于γ-變形菌綱(Gammaproteobacteria)中不同的目,因此,在圖4 的發育樹上,假單胞菌屬分支和弧菌屬分支于進化的較晚時期才分開。此外,圖4 還顯示了菌株S20 與黃桿菌屬菌株聚類于同一分支,而菌株E03 落在假單胞菌屬菌株聚類分支中,菌株W13和W14 則落在弧菌屬菌株聚類分支中。雖然菌株W13 和W14 在此系統發育樹上聚于同一小分支,但它們的產酶規律和菌落形態有明顯差異,提示它們可能屬于弧菌屬內不同的種。由菌落形態特征結合16S rDNA 系統發育分析,初步確定菌株S20 屬于黃桿菌屬(命名為Flavobacterium sp.S20),菌株E03 屬于假單胞菌屬(命名為Pseudomonas sp.E03),菌株W13 和W14 屬于弧菌屬(分別命名為Vibrio sp.W13,Vibrio sp.W14)。
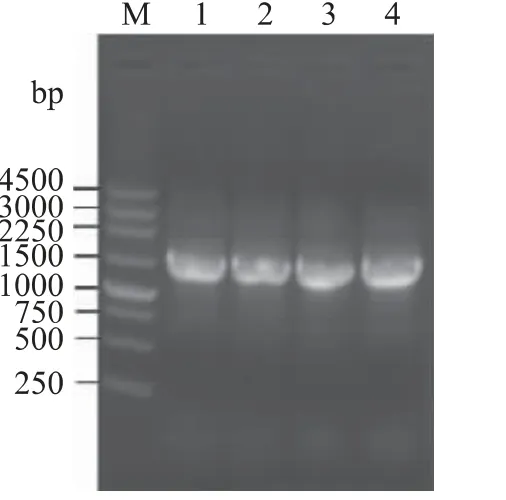
圖3 菌株E03、S20、W13 和W14 的16S rDNA 電泳圖譜Fig.3 Electrophoresis pattern of the PCR products of 16S rDNA of strains
微生物是褐藻膠裂解酶的重要來源,目前,在海洋細菌、土壤細菌、真菌中均發現有產褐藻膠裂解酶的菌種,其中假單胞菌屬、弧菌屬的產酶菌株報道較多[6],而黃桿菌屬產酶菌株報道較少[9]。假單胞菌是典型的聚甘露糖醛酸片段裂解酶生產源,某些假單胞菌菌株還產其他類型的褐藻膠裂解酶,如Pseudomonas aeruginosa PAO1 產偏好降解MG 交替嵌段的裂解酶[15]。海洋弧菌也是很好的獲取聚甘露糖醛酸片段裂解酶和聚古洛糖醛酸片段裂解酶的菌源[16-17]。黃桿菌菌株S20 產聚古洛糖醛酸片段裂解酶,并且研究發現該菌株還產其他類型的褐藻膠裂解酶[18]。本研究獲得的假單胞菌新菌株E03、弧菌新菌株W13 和W14、黃桿菌新菌株S20 可作為獲得不同類型褐藻膠裂解酶的菌種資源,為下一步酶的克隆表達,酶學性質研究以及酶法開發高價值化褐藻膠及其寡糖產品奠定基礎。
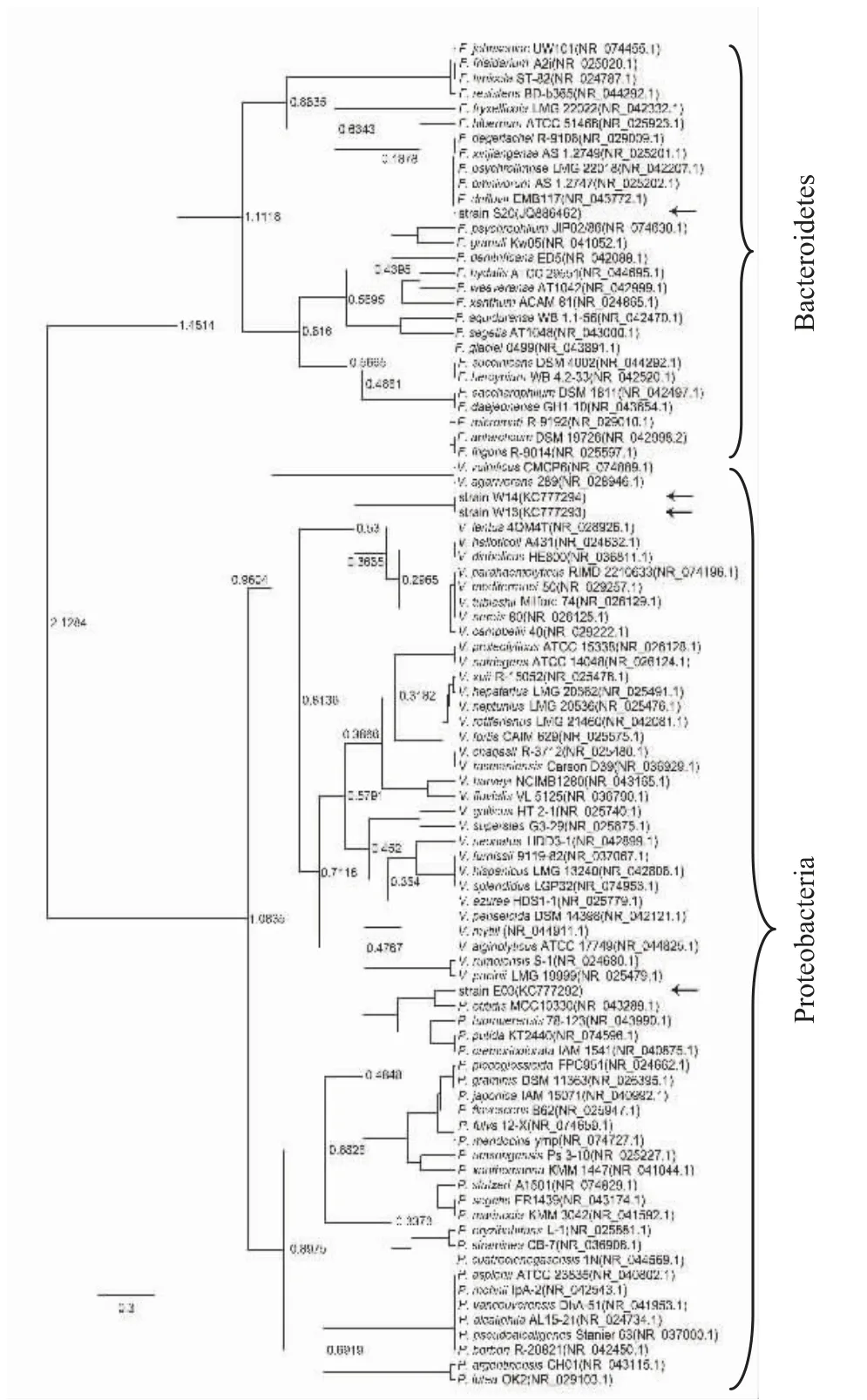
圖4 菌株E03、S20、W13 和W14 的16S rDNA 的序列系統發育樹Fig.4 The phylogenetic tree based on 16S rDNA sequence of four bacteria strains
3 結論
本研究采用褐藻酸鈉為唯一碳源的培養基,對采自海南的多類樣品進行菌株分離,然后選用觀察培養基粘度變化和酶活力檢測的雙重指標對菌株初篩,獲得具有產褐藻膠裂解酶能力的10 個菌株。通過測定發酵液酶活隨培養時間的變化,最終獲得高產褐藻膠裂解酶且產酶規律顯著差異的4 株菌。通過形態學結合16S rDNA 系統發育同源分析,初步鑒定菌株E03 屬于假單胞菌屬,菌株W13 和W14 屬于弧菌屬,而菌株S20 屬于產褐藻膠裂解酶報道較少的黃桿菌屬。
[1]Haug A,Larsen B,Smidsr?d O.Studies on the sequence of uronic acid residues in alginic acid[J].Acta Chem Scand,1967,21:691-704.
[2]郭遼樸,李洪軍.褐藻膠生物活性及在食品中應用的研究進展[J].四川食品與發酵,2007,43(6):9-12.
[3]Yamamoto Y,Kurachi M,Yamaguchi K,et al.Stimulation of multiple cytokine production in mice by alginate oligosaccharides following intraperitoneal administration[J].Carbohydr Res,2007,342(8):1133-1137.
[4]Tusi SK,Khalaj L,Ashabi G,et al.Alginate oligosaccharide protects against endoplasmic reticulum and mitochondrial mediated apoptotic cell death and oxidative stress [J].Biomaterials,2011,32(23):5438-5458.
[5]Kim HS,Lee CG,Lee EY.Alginate lyase:structure,property,and application[J].Biotechnol Bioprocess Eng,2011,16(5):843-851.
[6]Wong TY,Preston LA,Schiller NL.Alginate lyase:Review of major sources and enzyme characteristics,structure - function analysis,biological roles and applications [J].Annu Rev Microbiol,2000,54:289-340.
[7]Alkawash MA,Soothill JS,Schiller NL. Alginate lyase enhances antibiotic killing of mucoid Pseudomonas aeruginosa in biofilms[J].APMIS,2006,114(2):131-138.
[8]Wargacki AJ,Leonard E,Win MN,et al. An engineered microbial platform for direct biofuel production from brown macroalgae[J].Science,2012,335(6066):308-313.
[9]An QD,Zhang GL,Wu HT,et al.Properties of an alginatedegrading Flavobacterium sp. strain LXA isolated from rotting algae from coastal China[J].Can J Microbiol,2008,54(4):314-320.
[10]Kim DE,Lee EY,Kim HS,et al.Cloning and characterization of alginate lyase from a marine bacterium Streptomyces sp.ALG-5[J].Mar Biotechnol,2009,11(1):10-16.
[11]Tamura K,Peterson D,Peterson N,et al.MEGA5:molecular evolutionary genetics analysis using maximum likelihood,evolutionary distance,and maximum parsimony methods[J].Mol Biol Evol,2011,28(10):2731-2739.
[12]Drummond AJ,Suchard MA,Xie D,et al. Bayesian phylogenetics with BEAUti and the BEAST 1.7[J].Mol Biol Evol,2012,29(8):1969-1973.
[13]An QD,Zhang GL,Wu HT,et al.Production and partial properties of alginase from newly isolated Flavobacterium sp.LXA[J].Process Biochem,2008,43(8):842-847.
[14]Miller SR,Augustine S,Le Olson T,et al.Discovery of a free-living chlorophyII d-producing cyanobacterium with a hybrid proteobacterial/cyanobacterial small -subunit rRNA gene[J].PNAS,2005,102(3):850-855.
[15]Yamasaki M,Moriwaki S,Miyake O,et al.Structure and function of a hypothetical Pseudomonas aeruginosa protein PA1167 classified into family PL-7[J].J Biol Chem,2004,279(30):31863-31872.
[16]李京寶,于文功,韓峰,等.從海洋中分離的弧菌QY102褐藻膠裂解酶的純化和性質研究[J].微生物學報,2003,43(6):753-757.
[17]Wang Y,Guo EW,Yu WG,et al. Purification and characterization of a new alginate lyase from a marine bacterium Vibrio sp.[J].Biotechnol Lett,2013,35(5):703-708.
[18]Huang LS,Zhou JG,Li X,et al.Characterization of a new alginate lyase from newly isolated Flavobacterium sp.S20[J].J Ind Microbiol Biotechnol,2013,40(1):113-122.
