豆渣RNA 及其酶解物對小鼠免疫功能的影響
2013-08-07 09:13:06邵美麗陳慧燕
食品工業科技 2013年23期
邵美麗,陳慧燕,李 娜
(東北農業大學食品學院,黑龍江哈爾濱150030)
豆渣是大豆制品產業的主要副產物,目前有關豆渣的深加工利用非常局限,主要集中在膳食纖維、異黃酮和維生素E 等活性物質的開發方面[1-3]。隨著大豆產量的不斷增加和大豆加工行業的迅速發展,豆渣的綜合利用和深度開發成為急需解決的問題。核糖核酸(ribonucleic acid,RNA)是細胞內的重要成分,研究表明核糖核酸及核糖核苷酸對免疫系統正常功能、腸道生長發育、肝組織功能及脂代謝都有重要影響[4-8]。我國可提供的核酸資源較單一,工業上主要以廢啤酒酵母作為原料提取RNA,但由于獲取原料困難,不能滿足市場需要[9]。而豆渣中的核酸含量也不亞于酵母核酸,且豆渣價格低廉,來源豐富,是天然優質的核酸資源[10]。故本實驗室開展了有關豆渣RNA 制備、酶解及其功能方面的研究。本文主要利用實驗室前期制備好的豆渣RNA[11]及其酶解物,研究二者對小鼠的細胞免疫、體液免疫及非特異性免疫功能的影響,并比較二者免疫調節作用的差異,以期為豆渣資源的深度開發提供科學依據。
1 材料與方法
1.1 材料與儀器
Balb/c 小白鼠(清潔級,8 周齡,體重(18 ±2)g,雌雄各半) 哈爾濱獸醫研究所;豆渣RNA 及其酶解物 實驗室自制;刀豆蛋白A 天津市恒興化學試劑制造有限公司;四甲基偶氮唑鹽 天津博迪化工股份有限公司;RPMI1640 細胞培養液 天津市致遠化學試劑有限公司;小牛血清 杭州四季青生物工程公司;SRBC 哈爾濱獸醫研究所;豚鼠補體 實驗室自制;印度墨汁 北京市西中化工廠。
BIO RAD 680 酶聯免疫檢測儀 美國Labconco公司;TE2000 倒置顯微鏡 OLYMPUS 公司;MR23i低溫冷凍離心機 法國Jouan 公司。
1.2 實驗方法
1.2.1 動物分組及給藥 小白鼠隨機分為7 組,分別為正常對照組(生理鹽水)、豆渣RNA 高劑量組(10.0mg/mL)、豆渣RNA 中劑量組(1.0mg/mL)、豆渣RNA 低劑量組(0.1mg/mL)、豆渣RNA 酶解物高劑量組(10.0mg/mL)、豆渣RNA 酶解物中劑量組(1.0mg/mL )、豆渣 RNA 酶解物低劑量組(0.1mg/mL)。每組12 只,雌雄各半。每只每日灌胃0.5mL 受試物,連續30d,自由采食和飲水。
1.2.2 臟器指數的測定 參照文獻[12],實驗結束,處死小鼠,取胸腺和脾臟,用濾紙吸干表面血污,分別稱重,并按公式計算胸腺指數、脾臟指數。計算公式如下:

1.2.3 脾淋巴細胞增殖反應(MTT 法) 參照文獻[13],實驗結束,無菌取小鼠脾臟,置于平皿內200目的不銹鋼網上。用注射器針芯研磨,加入RPMI1640 培養液沖洗。將上述細胞懸液轉入2 倍體積的淋巴細胞分離液,1000r/min 離心10min,吸取中間的白膜層(單個核細胞層),用RPMI1640 培養液洗滌2 次后,計數,并調整濃度至1 ×106/mL。
將上述制備的脾細胞懸液分兩孔加入24 孔培養板中,每孔1mL,其中一孔加ConA(5.0μg/mL),另一孔作對照,置5% CO2,37℃CO2孵箱中培養72h。培養結束前4h,每孔吸取上清液0.7mL,加入0.7mL不含小牛血清的RPMI1640 培養液,同時加入MTT(5mg/mL)50μL/孔,繼續培養4h。培養結束后,每孔加入1mL 酸性異丙醇,吹打混勻,使紫色結晶完全溶解。然后分裝到96 孔培養板中,每組設3 個平行孔,測OD570值,用加ConA 孔的光密度值減去不加ConA 孔的光密度值代表淋巴細胞的增殖能力。
1.2.4 小鼠遲發型超敏反應(DTH)檢測 參照文獻[14],實驗結束,用20%(v/v)SRBC 懸液注射在小鼠右側足墊皮下,每鼠50μL,同時左側注射50μL 生理鹽水。24h 后,用游標卡尺測定小鼠左右足墊厚度,以左右足墊厚度之差為腫脹程度,同一部位測量3 次,取平均值。
1.2.5 小鼠脾抗體生成細胞檢測(定量溶血分光光度法) 參照文獻[15],于實驗第26d,向小鼠腹腔注射2%綿羊紅細胞(SRBC)0.2mL。第30d 處死小鼠,取脾臟并按照1.2.3 方法制備脾細胞懸液。然后取脾細胞懸液1mL,依次加入補體和0.2% SRBC 各1mL,于37℃溫浴1h,3000r/min 離心10min,取上清,測OD413值。
1.2.6 單核-巨噬細胞功能的測定(碳廓清實驗)參照文獻[16],實驗結束,小鼠尾靜脈注射稀釋的印度墨汁0.05mL。并分別于2、10min 后內毗靜脈叢采血2μL,并將血樣迅速吹到2mL 的0.1%碳酸鈉溶液中,然后測OD619值。按照下列公式計算碳粒廓清指數K 及吞噬指數α。
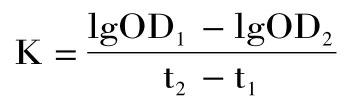
OD1:2min 時的吸光值;OD2:10min 時的吸光值;t1:2min;t2:10min。

2 結果與分析
2.1 豆渣RNA 及其酶解物對小鼠免疫器官重量的影響
胸腺和脾臟是機體內重要的免疫器官,與機體的細胞、體液免疫關系密切。當對機體使用免疫增強劑時會增加胸腺和脾臟的重量,故在一定程度上,胸腺和脾臟的重量可反映機體免疫功能的強弱,因此測量其重量可作為判斷機體免疫功能的輔助指標[17]。豆渣RNA 及其酶解物對小鼠免疫器官的影響如圖1 所示。
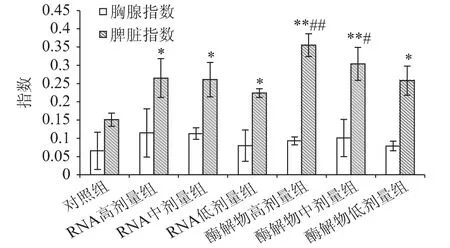
圖1 豆渣RNA 及其酶解物對小鼠免疫器官重量的影響Fig.1 Effect of okara RNA and hydrolysates on viscera index in mice
由圖1 可知,豆渣RNA 及其酶解物組各劑量組的胸腺指數雖然較對照組有所增加,但無顯著性差異(p >0.05),表明豆渣RNA 及其酶解物對胸腺無明顯影響。而豆渣RNA 及其酶解物組各劑量組的脾臟指數均顯著高于對照組(p <0.05),其中酶解物中、高劑量組極顯著高于對照組(p <0.01)。同時,酶解物高劑量還極顯著高于RNA 各劑量組(p <0.01),表明豆渣RNA 及其酶解物均可明顯促進小鼠脾臟生長,并在劑量范圍內呈現明顯的劑量效應關系,其中酶解物高劑量組效果最優。
2.2 豆渣RNA 及其酶解物對小鼠脾淋巴細胞增殖反應的影響
脾臟是各類免疫細胞居住的場所,也是對抗原物質產生免疫及產生免疫效應物質的重要基地。伴刀球蛋白(con A)可以選擇性地刺激T 淋巴細胞使之增殖,因此通過測定伴刀球蛋白誘導的脾淋巴細胞增殖能力可以反映機體的細胞免疫功能[18]。豆渣RNA 及其酶解物對小鼠脾淋巴細胞增殖反應的影響如圖2 所示。
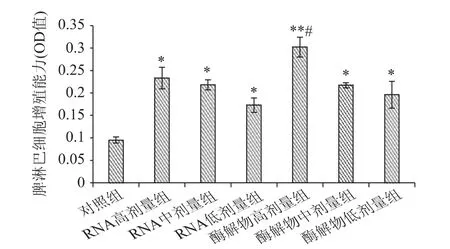
圖2 豆渣RNA 及其酶解物對小鼠脾淋巴細胞增殖反應的影響Fig.2 Effect of okara RNA and hydrolysates on lymphocyte transformation in mice
由圖2 可以看出,與對照組相比,豆渣RNA 及其酶解物各劑量組均能顯著提高小鼠脾淋巴細胞增殖能力(p <0.05),而且二者均呈現明顯的劑量效應關系。其中,酶解物高劑量組極顯著高于對照組(p <0.01),同時它還顯著高于RNA 各劑量組(p <0.05),表明豆渣RNA 及其酶解物均能明顯提高小鼠的脾淋巴細胞增殖能力,增強小鼠細胞免疫功能,其中酶解物高劑量組的效果最顯著。
2.3 豆渣RNA 及其酶解物對小鼠遲發型變態反應的影響
DTH(遲發型變態反應)是檢測T 細胞功能的體內實驗方法之一,通過測定受測物對機體遲發型超敏反應的影響,可反映機體的T 細胞功能或受試物的作用[19]。豆渣RNA 及其酶解物對小鼠遲發型變態反應的影響如圖3 所示。
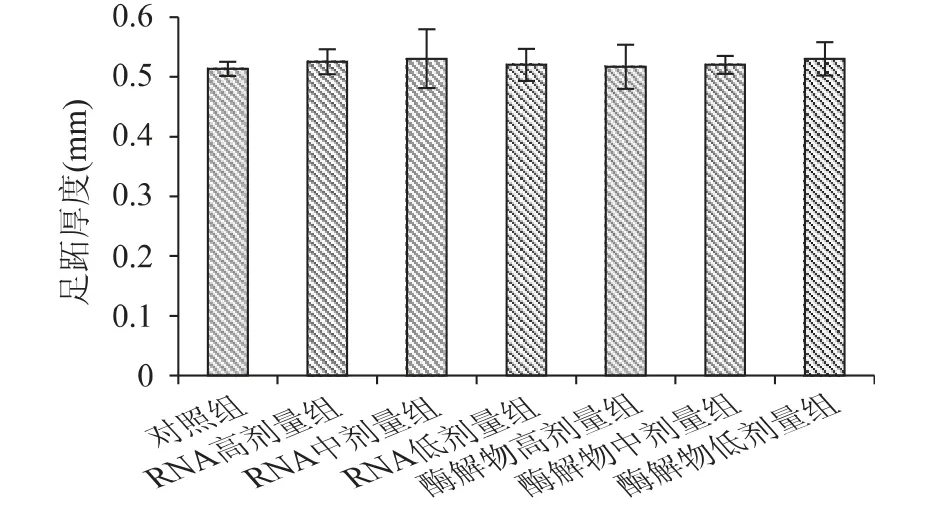
圖3 豆渣RNA 及其酶解物對小鼠的遲發型變態反應影響Fig.3 Effect of okara RNA and hydrolysates on delayedtype hypersensitivity in mice
由圖3 可知,經口給予小鼠不同劑量的豆渣RNA 及其酶解物30d 后,盡管豆渣RNA 組及其酶解物各劑量組的足趾增厚值比對照組有所增加,但未達到顯著性水平(p >0.05),說明豆渣RNA 及其酶解物對T 細胞介導的遲發型超敏反應的影響不明顯。
2.4 豆渣RNA 及其酶解物對小鼠脾臟抗體生成細胞的影響
脾臟的抗體生成細胞水平是反映機體體液免疫功能的重要指標之一[10]。豆渣RNA 及其酶解物對小鼠脾臟抗體生成細胞的影響如圖4 所示。
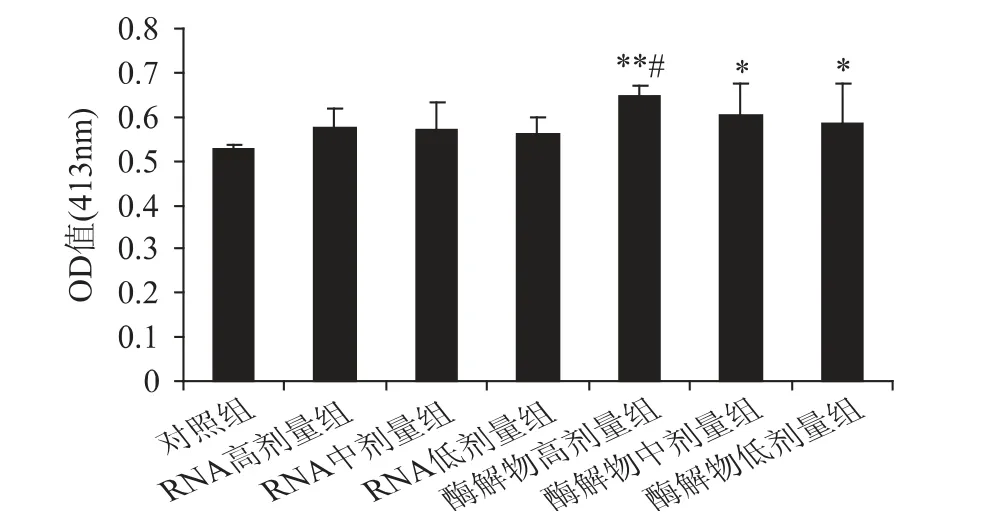
圖4 豆渣RNA 及其酶解物對小鼠脾臟抗體形成細胞的影響Fig.4 Effect of okara RNA and hydrolysates on the antibody forming cell in mice
由圖4 可以看出,豆渣RNA 組及其酶解物各劑量組小鼠的脾臟抗體生成細胞水平均高于對照組。其中,豆渣RNA 各劑量組與對照組相比,差異不顯著(p >0.05),而豆渣RNA 酶解物各劑量組則顯著高于對照組(p <0.05),且酶解物高劑量組極顯著高于對照組(p <0.01),同時顯著高于RNA 各劑量組(p <0.05),并呈現明顯的劑量效應關系。表明豆渣RNA酶解物可明顯增加脾臟的抗體生成細胞水平,增強小鼠的體液免疫功能。其中酶解物高劑量組的效果最好。
2.5 豆渣RNA 及其酶解物對小鼠單核-巨噬細胞功能的影響
巨噬細胞是機體天然免疫防御系統中重要的免疫效應細胞,其吞噬功能可反映機體非特異性免疫功能的強弱[21]。豆渣RNA 及其酶解物對小鼠單核-巨噬細胞吞噬功能的影響如圖5 所示。
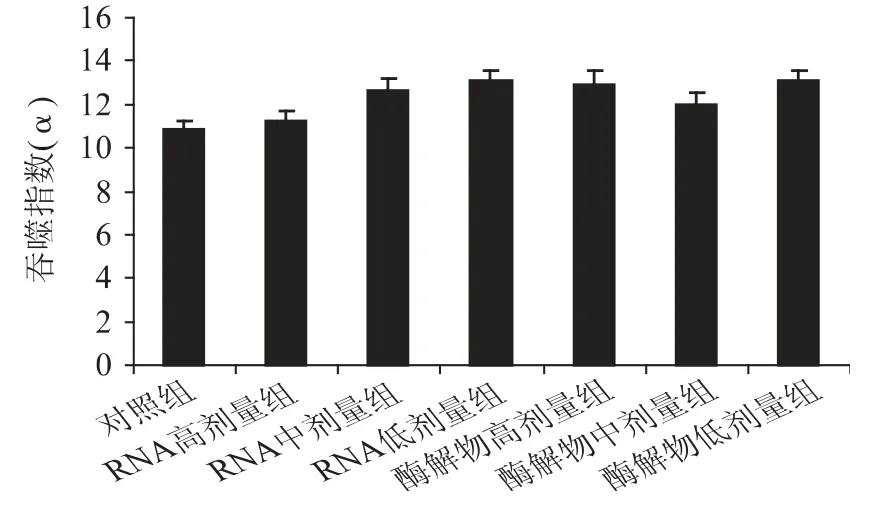
圖5 豆渣RNA 及其酶解物對小鼠巨噬細胞吞噬功能的影響Fig.5 Effect of okara RNA and hydrolysates on ability of carbon clearance in mice
由圖5 可知,與對照組相比,豆渣RNA 及其酶解物各劑量組吞噬指數雖有所提高,但差異不顯著(p >0.05),表明豆渣RNA 及其酶解物不能明顯促進小鼠巨噬細胞的吞噬功能,提示豆渣RNA 及其酶解物對機體的非特異性免疫調節作用不明顯。
3 討論
研究表明RNA 是維持機體正常免疫功能的必需營養成分。補充外源性RNA,不僅可以增強機體的免疫功能,還能恢復由蛋白質缺乏或其他原因引起的免疫功能喪失[22]。豆渣RNA 作為一種新的RNA 資源,其免疫活性如何,尚未見報道。本研究依照《保健食品檢驗與評價技術規范》的要求,分別從免疫器官重量、細胞免疫功能、體液免疫功能和單核-巨噬細胞功能4 個方面探討了豆渣RNA 及其酶解物在不同劑量水平下對小鼠免疫功能的影響,并比較了二者免疫調節作用的差異。
本實驗表明豆渣RNA 及其酶解物均能促進小鼠脾臟生長,增強脾淋巴細胞增殖能力,提高小鼠脾臟細胞的抗體生成水平,二者均呈現劑量依賴性。這說明豆渣RNA 及其酶解物可通過提高小鼠的細胞免疫和體液免疫增強機體的整體免疫能力。此結果與王蘭芳等研究日糧中添加核苷酸能極顯著提高免疫抑制小鼠的抗體形成細胞的數目和淋巴細胞轉化率的結果相似[23]。
豆渣RNA 與豆渣RNA 酶解物相比,在增強脾臟指數,淋巴細胞增殖能力和抗體生成量方面,RNA 酶解物的高劑量組顯著優于RNA 其他各劑量組,RNA酶解物的中、低劑量組較相對應的RNA 中、低劑量組雖然也有提高,但差異不顯著。這說明豆渣RNA酶解中小分子的低聚寡核苷酸可能更加易于小腸吸收,所以酶解物組的免疫效果相比非酶解物組好,但RNA 酶解物高劑量組(10.0mg/mL)的效果最明顯。
此外,本實驗表明豆渣RNA 及其酶解物不能明顯促進小鼠巨噬細胞的吞噬功能。且豆渣RNA 及其酶解物各劑量組吞噬指數未呈現劑量依賴性。豆渣RNA 組出現隨劑量增加,吞噬指數逐漸降低的趨勢,推測可能因劑量過大造成機體短期刺激過大,影響其炭粒廓清能力,導致吞噬指數降低。豆渣RNA酶解物組則出現隨劑量增加,吞噬指數先降低后增加的趨勢,此原因有待于下一步在本實驗劑量(0.1~10.0mg/mL)范圍內,減小劑量設計倍數,增加實驗組數,進一步實驗后分析探討。
4 結論
豆渣RNA 及其酶解物均能促進小鼠脾臟生長,增強脾淋巴細胞增殖能力,提高抗體生成細胞水平,但對巨噬細胞吞噬功能影響不明顯。這表明豆渣RNA 及其酶解物主要通過促進機體的細胞免疫和體液免疫而發揮免疫調節作用。其中,豆渣RNA 酶解物高劑量組(10.0mg/mL)的效果明顯優于豆渣RNA 組。
[1]陳霞,楊香久,徐永華,等.豆渣膳食纖維制備及其在食品中的應用[J].中國糧油學報,2002,17(2):1-3.
[2]高金燕,徐江林.從大豆豆渣中提取大豆異黃酮的初步研究[J].中國食品添加劑,2003(5):16-18.
[3]戴小陽,羅澤民.豆渣的有效利用[J].湖南農業科學,1996(1):28-29.
[4]徐慧,劉建軍,趙祥穎.啤酒廢酵母提取核糖核酸工藝的研究[J].中國釀造,2008,35(6):35-37.
[5]胡剛,孫軍勇,蔡國林,等.濃鹽法提取啤酒廢酵母核糖核酸的研究[J].中國釀造,2009(7):112-114.
[6]Marina C E,Victor L,María F.Extraction of RNA from fermented milk products for in situ gene expression analysis[J].Analytical Biochemistry,2010,10(2):307-309.
[7]Ha S,Li K,Lori A K,et al.Evaluation of microbial RNA extractions from Streptococcus pneumonia [J]. Journal of Microbiological Methods,2007,68:342-348.
[8]Luliana S,Keith S,Bhoomi M. Emerging role of small ribonucleic acid in gastrointestinal tumors:Critical Reviews in oncology[J].Hematology,2010,76(3):173-185.
[9]孫榮丹,劉瑩,張洪林,等.濃鹽法與稀堿法在啤酒廢酵母中提取RNA 的研究[J].氨基酸和生物資源,2006,28(3):76-78.
[10]畢韜韜,吳廣輝,高愿軍.豆渣深加工研究進展[J].食品研究與開發,2011,32(6):149-151.
[11]李娜,柴菜建,邵美麗,等.工業化方法提取及純化豆渣RNA 的工藝研究[J].食品工業,2013,34(1):4-7.
[12]李靜,劉力源,任捷,等.地錦粗提物對脾虛小鼠臟器指數和抗氧化能力的影響[J].中國微生態學雜志,2011(12):1060-1063.
[13]Avantika V,Kashi N P,Aloukick K S.Evaluation of the MTT lymphocyte proliferation assay for the diagnosis of neurocysticer cosis[J].Microbiological Methods,2010,81(2):175-178.
[14]張秀美.新編獸醫實用手冊[M].濟南:山東科學技術出版社,2006:8-10.
[15]Christopher C G,Carola G V,Katrina L R.Control systems and decision making for antibody production [J]. Nature Immunology,2010,11:681-688.
[16]Dongsan P,Dae-Kwon B,Jeong H J.Immunopotentiation and antitumor effects of a ginsenoside Rg3- fortified red ginseng preparation in mice bearing H460 lung cancer cells[J].Environmental Toxicology and Pharmacology,2011,31 (3):397-405.
[17]Kulkarni A D,Rudolph F B,Van-Buren C T.The role of dietary sources of nucleotides in immune function:a review[J].Nutrure,1994,124(8):1442-1446.
[18]楊小軍,左偉勇,陳偉華.面筋蛋白的胃蛋白酶酶解物對大鼠免疫功能的影響[J].南京農業大學學報,2004,27(4):69-72.
[19]霍乃蕊,岳文斌,劉玉花.羊骨木瓜蛋白酶水解物對小鼠免疫功能的影響[J].中國實驗動物學報,2010,5(18):418-420.
[20]Clement K A,Andrew A A,Keiku Y.Dietary nucleosidenucleotides mixture aggravates nasal allergic responses induced by toluene diisocyanate in mice[J].Nutrition Research,2010,17(10):1597-1605.
[21]Huang Q,Li Z,Yang XF.Flow cytometry in evaluating phagoeytosis of mouse peritoneal Macrophages[J]. Chin J Pharmacol Toxicol,2007,21(2):140-146.
[22]李蓉,佟曉永,潘宏志.飲食核酸對不同齡小鼠免疫功能的影響[J].營養學報,2000,22(1):36-39.
[23]王蘭芳,樂國偉,施用暉.外源核苷酸對小鼠免疫功能的影響[J].營養學報,2003,25(3):298-301.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
課堂內外·初中版(科學少年)(2023年10期)2023-12-10 00:43:06
中華詩詞(2022年6期)2022-12-31 06:41:24
全科護理(2022年10期)2022-12-26 21:19:15
國際放射醫學核醫學雜志(2021年10期)2021-02-28 08:41:58
學苑創造·A版(2020年9期)2020-10-13 09:41:02
中國科技論壇(2017年7期)2017-07-25 08:49:53
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
中國合理用藥探索(2014年11期)2014-03-11 20:30:20
中國中醫藥現代遠程教育(2014年22期)2014-03-01 04:32:55
