高山被孢霉Δ5去飽和酶基因的克隆及異源表達研究
2013-08-07 08:51:00劉建民栗茂騰
食品科學 2013年15期
劉建民,栗茂騰
(1.華中科技大學生命科學與技術學院,湖北 武漢 430074;2.河南理工大學資源環境學院,河南 焦作 454000)
生物體內的脂肪酸分為飽和脂肪酸、單不飽和脂肪酸和多不飽和脂肪酸。多不飽和脂肪酸(polyunsaturated fatty acids,PUFAs)是對人體有重要生理活性的物質,廣泛應用于保健品和醫藥等行業。花生四烯酸(20:4Δ5,8,11,14,arachidonic acid,AA)是研究、應用較多的多不飽和脂肪酸之一,其廣泛分布于動物的中性脂肪中[1]。在腦和神經組織中,AA的含量通常占總PUFAs的40%~50%,在神經末梢中更是高達70%,它在促進大腦和神經發育方面有著重要的影響[2]。前人的研究結果表明,AA在細胞內具有第二信使的作用,參與調控細胞的生命活動[3];同時,在改善記憶力和視力、神經功能調節和炎癥反應中都有重要的作用[4]。
AA有重要的生理功能,但人體不能自身合成,必須從外界攝取獲得。被孢霉(Mortierella)的發酵生產是目前AA最主要的來源,但通過被孢霉發酵獲得的AA有較濃的產生菌氣味。在PUFAs的合成過程中,以雙高γ-亞麻酸(20:3Δ8,11,14,dihomo-γ-linolenic acid,DHGLA)為底物,Δ5去飽和酶催化脂肪酸鏈第5、6位碳原子間脫氫形成AA。Δ5去飽和酶是AA合成途徑中的關鍵酶,因此,克隆Δ5去飽和酶基因并對其功能進行驗證,可以對AA的發酵生產提供理論支持。
本研究以實驗室篩選的高山被孢霉W15為材料,通過RT-PCR的方法克隆到Δ5去飽和酶基因,將該基因轉化畢赤酵母GS115菌株后,得到了表達產物,這為AA新來源的拓展提供參考。
1 材料與方法
1.1 菌株、試劑與培養基
菌株 高山被孢霉W15為實驗室篩選和保種的菌株。
連接酶、限制性內切酶 加拿大Fermentas公司;雙高γ-亞麻酸 美國Sigma公司;pMD18-T Simple Vector、Escherichia coli DH5α 寶生物(大連)有限公司;pPIC3.5K載體、Pichia pastoris GS115 美國Invitrogen公司;G418(遺傳霉素) Genveiw公司;A反應液 北京天根生化科技有限公司;其他試劑為進口試劑或國產分析純。
MD培養基:酵母氮堿基13.4g、生物素4×10-4g、葡萄糖10g,用蒸餾水定容至1L;YPD培養基:每升含有酵母提取物10g、蛋白胨20g、葡萄糖20g;BMGY培養基:酵母提取物10g、蛋白胨20g、1mol/L磷酸鉀緩沖液100mL、酵母氮堿基13.4g、甘油10mL,定容至1L;BMMY:酵母提取物10g、蛋白胨20g、1mol/L磷酸鉀緩沖液100mL、酵母氮堿基13.4g、甲醇10mL,定容至1L。
1.2 方法
1.2.1 高山被孢霉Δ5去飽和酶基因的克隆
將高山被孢霉接種到土豆培養基中,25℃振蕩培養5d,真空抽慮收集菌體,按照Invitrogen公司的Trizol RNA提取試劑盒的說明提取高山被孢霉的總RNA,用First Strand cDNA Synthesis Kit (Fermentas)合成第一鏈cDNA。
Δ5去飽和酶基因引物參考GenBank已提交序列(AF054824),根據Michaelson等[5]進行設計,在上游引物引入Kozak序列[6],引物擴增的片段為Δ5去飽和酶基因的完整序列,引物兩端中引入EcoR Ⅰ酶切位點。Δ5L1:5’-GCGA ATTCGCCACCATGGGTACGGACCAAGGAAA-3’;Δ5R1:5’-CGGAATTCCTACTCTTCCTTGGGACGGAG-3’。
以合成的第一鏈cDNA為模板,用ToYoBo公司的KOD plus高保真酶進行擴增,反應體系為50μL。根據引物設計軟件Oligo 6.0推薦的退火溫度,PCR反應程序為:95℃預變性2min,然后進行30個循環的擴增反應,每個循環包括94℃變性20s,57℃退火30s,68℃延伸1min 30s。
PCR產物用0.8%的瓊脂糖凝膠進行電泳分離,回收純化PCR產物。由于用高保真酶擴增的PCR產物為平末端,因此,需對回收的PCR產物進行加A反應,便于與T載體的連接,取15μL回收PCR產物,加入4μL 5×加A反應液和1μL 2.5U/μL Taq DNA聚合酶,混勻后,用PCR儀72℃保溫40min完成加A反應。加A反應的產物與pMD18-T Simple Vector進行連接和轉化大腸桿菌DH5α感受態細胞,用菌液PCR的方法篩選陽性克隆,送北京三博生物技術有限公司測序。
1.2.2 畢赤酵母表達載體的構建
用EcoR I酶切重組的T載體,得到Δ5去飽和酶基因片段,將該片段連接到經EcoR I消化和CIAP去磷酸化的pPIC3.5K載體上,轉化E. coli DH5α并用50mg/mL的卡那霉素進行篩選。為了鑒定基因在酵母表達載體上的插入方向,合成一條酵母表達載體通用引物,該引物序列位于pPIC3.5K的5’AOX1啟動子的下游,序列如下:
5’AOX1:5’-GACTGGTTCCAATTGACAAGC-3’。
PCR擴增時,用5’AOX1引物和擴增基因時使用的下游引物對目的基因進行PCR擴增;取1μL菌液做模板,預變性時間為6min,其他按照擴增基因的條件進行PCR反應。如擴增產物大小與目的片段相同,則可初步確定該克隆為陽性克隆且基因為正向連接,該重組質粒命名為pPIC3.5K-D5;否則是假的陽性克隆或者基因在載體中插入的方向相反。
1.2.3 重組酵母載體對畢赤酵母GS115的轉化
StuⅠ線性化重組質粒pPIC3.5K-D5,回收后采用電擊轉化法轉化感受態畢赤酵母GS115,MD選擇性培養基平板進行初篩。對平板上長出的菌落分別涂布在G418質量濃度為0.5、0.75、1、2、3、4mg/mL的YPD平板進行復篩,選擇耐受高質量濃度G418的轉化菌株用于表達研究,同時,以轉化空質粒pPIC3.5K的酵母作為空白對照。
1.2.4 酵母轉化子的鑒定
取200μL轉化酵母菌液,25℃、3000r/min離心2min。倒去上清,加入100μL 0.2% SDS,重新懸浮酵母菌體,把懸浮的菌液煮沸5min后,用離心機短暫離心10s。取處理后的上清1μL做模板,以Δ5L1/Δ5R1為引物,PCR鑒定陽性轉化酵母。
1.2.5 重組畢赤酵母的誘導表達
重組酵母接種到裝有10mL BMGY培養基的50mL三角瓶中,30℃、200r/min搖床培養16~18h。以1%接種量轉接到裝有50mL BMMY培養基的250mL三角瓶中,30℃、200r/min搖床培養。每24h向培養基中添加甲醇,使其終濃度為0.6%。持續培養5d,離心收集菌體,用去離子水洗滌3~4次。
1.2.6 酵母油脂的提取及脂肪酸成分分析
收獲菌體于50℃烘干后,研碎,用3mL石油醚(30~60℃沸程)抽提2次,在旋轉蒸發儀上快速蒸干溶劑;加入0.4mol/L KOH甲醇溶液1mL,80℃水浴中皂化3~5min,冷卻后加入14%三氟化硼乙醚甲醇溶液2mL,于80℃水浴中振蕩處理1min,待冷卻至室溫,加入正己烷1mL,加入飽和氯化鈉溶液1mL,反復顛倒幾次,靜置3~5min,上清即為甲酯化的脂肪酸正己烷溶液。
甲酯化樣品的分析在華中科技大學分析測試中心完成,使用Agilent 7890A-5975C氣質聯用儀,儀器設置參數和分析條件如下:進樣口溫度為250℃,50:1分流比,1μL進樣;色譜柱用強極性柱HP-INNOWax(30m×0.25mm,0.25μm);載氣為He,流速為1mL/min;升溫程序為:80~200℃,25℃/min;200~230℃,3℃/min;230℃保留10min。
2 結果與分析
2.1 Δ5去飽和酶基因的克隆與序列分析
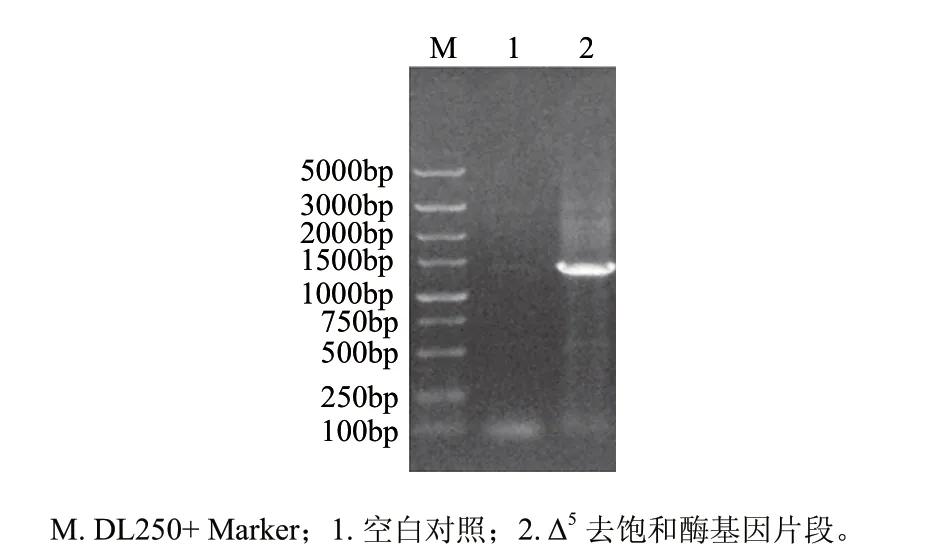
圖 1 高山被孢霉Δ5去飽和酶基因RT-PCR產物的瓊脂糖凝膠電泳Fig.1 Agarose gel electrophoresis of the RT-PCR product of Mortierella alpina Δ5 desaturase gene

圖 2 高山被孢霉Δ5去飽和酶基因推導氨基酸序列比對Fig.2 Alignment of a deduced amino acid sequence of the Δ5 desaturase gene from Mortierella alpina
提取高山被孢霉W15的總RNA,以合成的cDNA為模板,擴增得到約1400bp的片段(圖1),與T載體連接后進行測序,結果表明,所克隆的Δ5去飽和酶基因全長1341bp(GenBank登錄號:FJ625826),與參考Δ5去飽和酶基因核苷酸序列同源性為90%,但其推導氨基酸序列與參考基因編碼氨基酸序列的同源性達到了95%,這說明Δ5去飽和酶基因在不同高山被孢霉菌株中的核苷酸序列變化較大,但它們編碼的氨基酸序列相對較為保守。對推導的氨基酸序列進行分析顯示,其N末端存在去飽和酶家族廣泛存在的細胞色素b5結構域及保守的HPGG氨基酸序列,同時還存在3個保守的組氨酸富集區HDASH、HMLGHH和QAVHH(圖2中虛線表示),這與前人的研究結果一致[5,7]。
2.2 pPIC3.5K-D5酵母表達載體的構建及對畢赤酵母的轉化
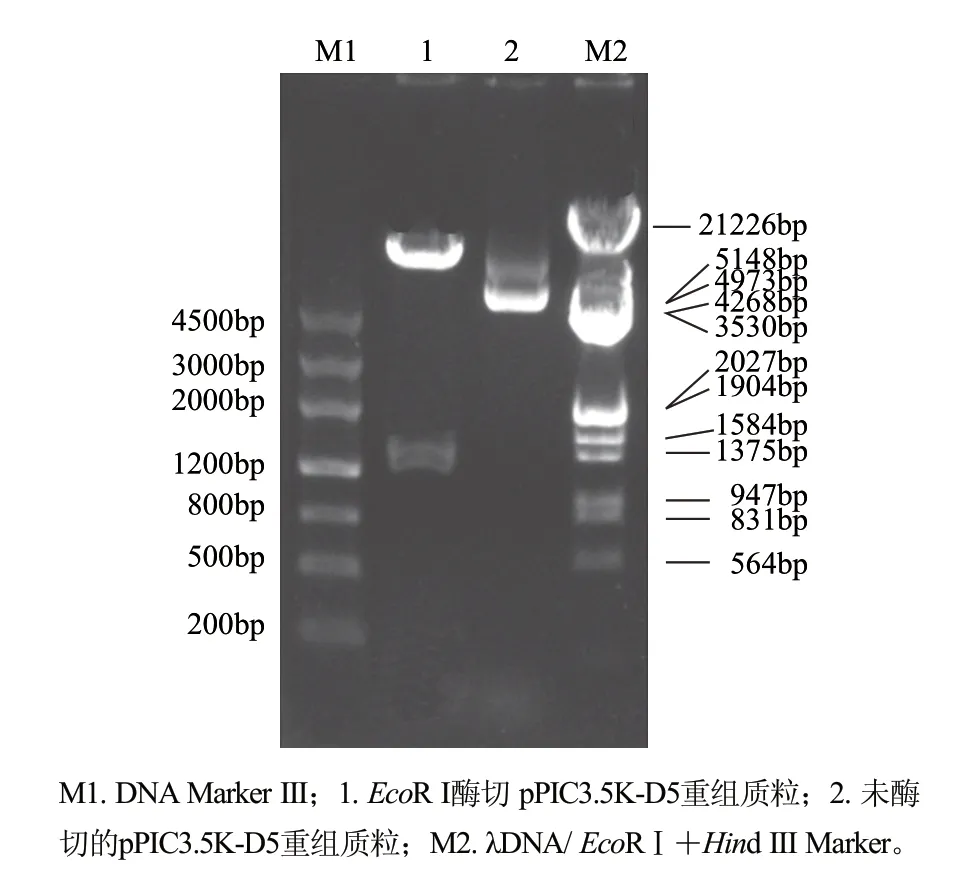
圖 3 重組質粒pPIC3.5K-D5酶切鑒定Fig.3 Identification of pPIC3.5K-D5 with EcoR Ⅰ digestion
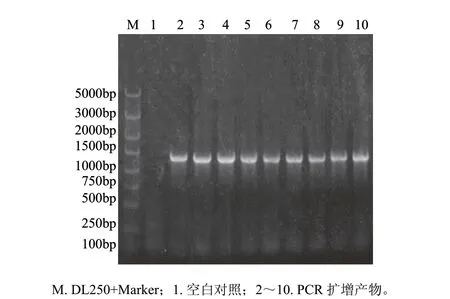
圖 4 轉化畢赤酵母菌液擴增Δ5去飽和酶基因Fig.4 Δ5 desaturase gene amplification of recombinant P. pastoris GS115
為了驗證Δ5去飽和酶基因的功能,將連接在T載體上的Δ5去飽和酶基因用EcoR I切下,亞克隆到經EcoR I消化和CIAP處理的pPIC3.5K表達載體上,轉化E. coli DH5α后,用AOX1引物和Δ5R1引物驗證基因插入方向。提取基因正向插入的重組質粒,用EcoR I進行酶切驗證,結果見圖3。酶切后,在1400bp和9000bp處有預期條帶出現,證明該基因已成功亞克隆到pPIC3.5K表達載體中。
用Stu I線性化重組質粒pPIC3.5K-D5,以電擊法轉化畢赤酵母GS115感受態細胞。通過MD平板和含不同質量濃度G418的YPD平板篩選,在4mg/mL G418的平板上得到較多的高抗性重組子。隨機挑取9個單菌落,用Δ5L1和Δ5R1引物進行PCR鑒定,見圖4,所挑取的菌落均能擴增出與目標條帶大小一致的片段,這個結果表明Δ5去飽和酶基因已整合到酵母基因組中。
2.3 表達產物的GC-MS分析
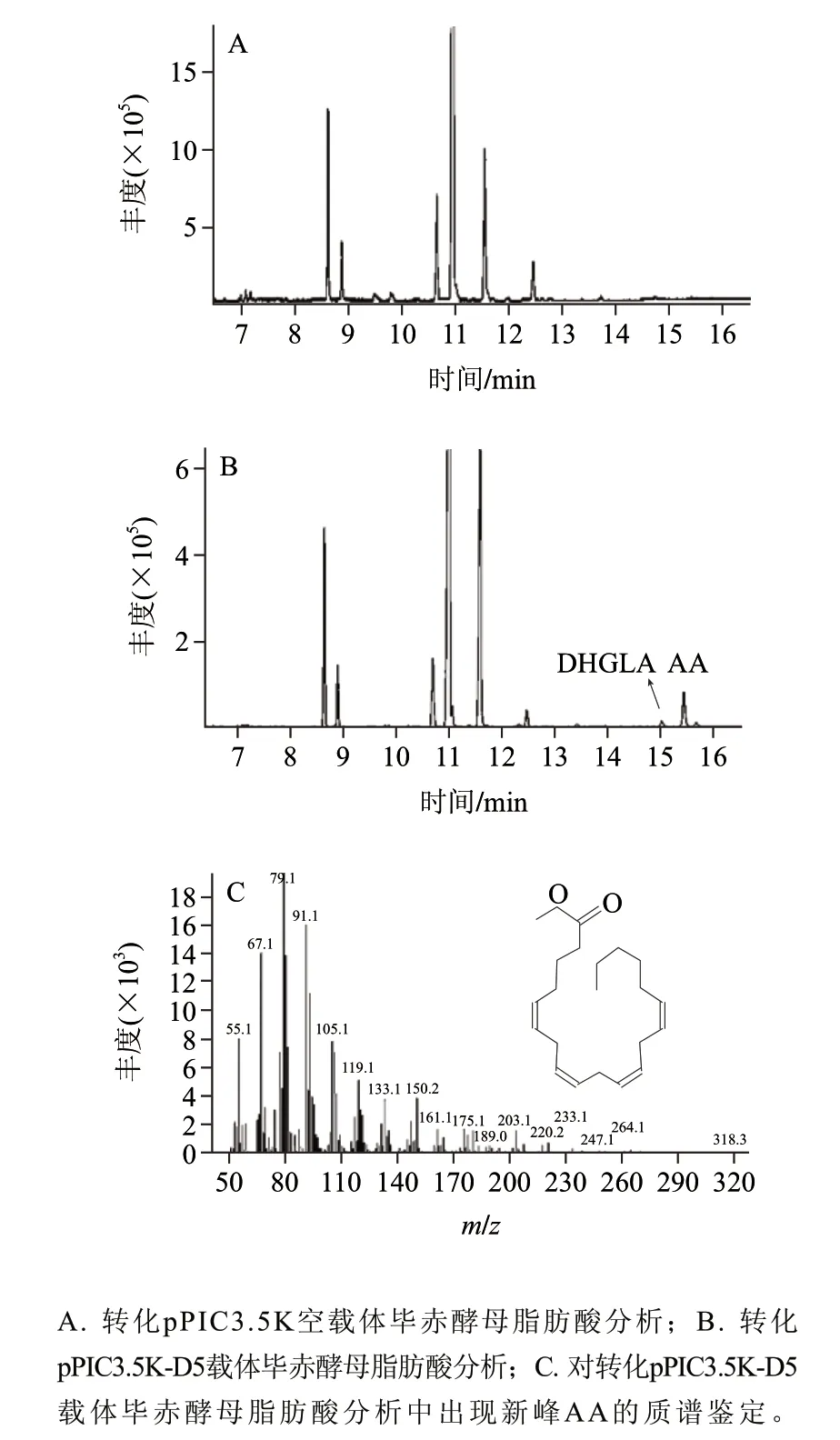
圖 5 轉化Δ5去飽和酶基因酵母重組子脂肪酸GC-MS分析Fig.5 GC-MS analysis of fatty acid composition in total lipids from P. pastoris GS115 transformants
用甲醇誘導畢赤酵母轉化子,以驗證Δ5去飽和酶基因是否具有功能。在培養基中添加Δ5去飽和酶的作用底物雙高γ-亞麻酸,培養結束后分離酵母細胞并提取細胞內中總脂肪酸,甲酯化后進行GC-MS分析,結果如圖5B,添加外源雙高γ-亞麻酸作為Δ5去飽和酶底物時,重組畢赤酵母能利用該底物產生AA。對重組畢赤酵母的脂肪酸進行分析表明,雙高γ-亞麻酸占總脂肪酸的0.45%,AA占酵母總脂肪酸的2.85%,說明被酵母利用的雙高γ-亞麻酸在Δ5去飽和酶的催化下,大部分轉化為AA。朱敏等[7]把高山被孢霉M6的Δ5去飽和酶基因構建在畢赤酵母pPIC9K載體上,并在畢赤酵母中做了功能驗證,發現該酶能催化合成占總脂肪酸5.8%的AA生成,AA的產量是本實驗結果的2倍。出現這一結果可能存在如下兩個原因:1)不同的表達載體對基因的表達量和酶活性有一定的影響,不同菌株來源的相同基因在同樣的表達載體中表達具有更好的對比性;2)這兩個基因均編碼446個氨基酸,其比對結果中僅有11個氨基酸存在差異,這揭示了不同種屬來源的相同基因,在進化過程中,基因序列的變化導致氨基酸序列的改變,最終使這些基因在外源宿主中的表達水平可能會有較大的差別。因此,比較不同來源的相同基因的DNA序列和編碼的氨基酸序列,分析發生突變的氨基酸位點和影響酶活的關鍵位點,可為進一步定向改造基因的結構提供有力的幫助。由表1可知,重組畢赤酵母中的亞油酸(linoleic acid,LA)與含空載體酵母中LA相比,含量從9.61%提高到23.58%,說明在畢赤酵母培養過程中,添加一定量的外源雙高γ-亞麻酸,可能會提高LA的產量。
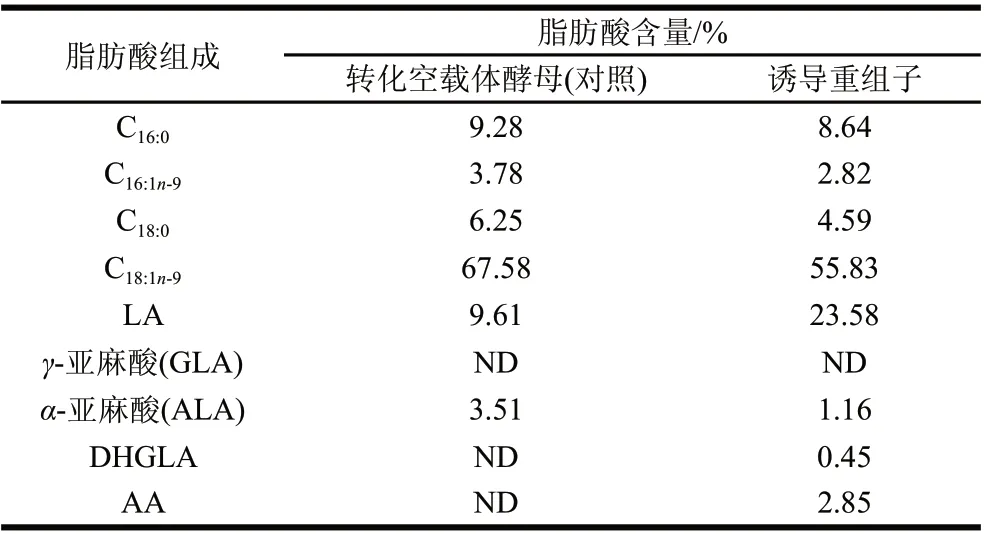
表 1 轉化Δ5去飽和酶基因的酵母的脂肪酸成分分析(n=3)Table 1 Fatty acid composition of recombinant P. pastoris GS115 (n=3)
3 討 論
花生四烯酸(AA)是一種人體必需的多不飽和脂肪酸,對嬰兒的大腦發育尤其重要[8]。微生物發酵是生產AA最重要的方式,高產菌株的篩選和發酵工藝的改進可以提高微生物發酵生產AA的產量[9-10],利用構建工程菌株可以增加AA生產菌株的來源。本研究以實驗室篩選的高山被孢霉W15菌株為出發材料,通過RT-PCR的方法克隆了與AA合成相關的Δ5去飽和酶基因,BLAST結果表明,該基因與參考序列的同源性為90%,與GenBank所有提交序列(AF067654)的同源性最高為91%,推導的氨基酸序列與報道的氨基酸序列的同源性達到了95%,說明同一物種在進化過程中,Δ5去飽和酶基因的核苷酸序列變化較大,但它們編碼的氨基酸序列相對較為保守,這在一定程度上體現了物種的變異是一個長期的、緩慢的過程。對Δ5去飽和酶基因推導的氨基酸序列進行分析顯示,其N末端存在去飽和酶家族廣泛存在的細胞色素b5結構域及保守的HPGG氨基酸序列,同時還存在3個保守的組氨酸富集區,這一結果在其他的去飽和酶中也有發現[11-12]。
畢赤酵母表達系統是目前應用較多的一種有效的外源基因表達系統。根據載體的不同,可進行胞內表達或胞外分泌表達,其生長過程中合成多種飽和和不飽和的脂肪酸,因此,是進行研究多不飽和脂肪酸表達的系統之一[13-15]。本研究將克隆的Δ5去飽和酶基因連接到載體上后轉入畢赤酵母,在添加特異性底物雙高γ-亞麻酸的前提下,生成了占總脂肪酸2.85%的AA,證明該基因具有催化的活性。但AA的產量很低,根據對基因序列和推導的氨基酸序列的分析,通過改變基因中部分堿基,從而改變其氨基酸組成及酶分子的結構,可能會達到提高AA產量的目的。另外,構建產AA的基因工程菌,必須要考慮基因表達載體及工程菌產物對人、動物和環境的安全。一般認為,安全的微生物基因作為載體,修飾后不留下任何抗生素基因才是安全的[16]。本實驗中所用到的畢赤酵母表達載體pPIC3.5K具有氨芐青霉素和卡那霉素兩個抗性基因,在進行外源基因表達時還需要添加甲醇做誘導,因此,使用畢赤酵母表達系統生產供人類利用的產物時,需要對表達載體進行改造,消除抗生素基因,改變與甲醇誘導相關的基因序列和選擇其他安全的誘導物,保證產物對人類的安全。
[1] 姚昕, 秦文, 齊春梅, 等. 花生四烯酸的生理活性及其應用[J]. 糧油加工與食品機械, 2004(5): 57-59.
[2] 丁兆坤, 劉亮, 許友卿. 二十碳四烯酸研究[J]. 水產科學, 2007,26(12): 684-688.
[3] PIOMELLI D, VOLTERRA A, DALE N, et al. Lipoxygenase metabolites of arachidonic acid as second messengers for presynaptic inhibition of aplysia sensory cells[J]. Nature, 1987, 328: 38-43.
[4] 林樹新, 於峻. 花生四烯酸代謝物對呼吸道感受器的作用[J]. 生理學報, 2007, 59(2): 141-149.
[5] MICHAELSON L V, LAZARUS C M, GRIFFITHS G, et al. Isolation of a delta5-fatty acid desaturase gene from Mortierella alpina[J].Journal of Biological Chemistry, 1998, 273(30): 19055-19059.
[6] KOZAK M. Compilation and analysis of sequences upstream from the translational start site in eukaryotic mRNAs[J]. Nucleic Acids Research, 1984, 12(2): 857-872.
[7] 朱敏, 劉智, 余龍江, 等. 高山被孢霉Δ5去飽和酶基因的分離與驗證[J]. 遺傳學報, 2005, 32(9): 986-992.
[8] 詹現璞, 呂銀德, 趙俊芳. 花生四烯酸在乳制品中的應用[J]. 中國食品添加劑, 2009(4): 155-159.
[9] 朱路英, 張學成, 宋曉金. n-3多不飽和脂肪酸DHA、EPA研究進展[J]. 海洋科學, 1997, 31(11): 78-85.
[10] 張燕鵬, 黃鳳洪, 楊湄, 等. 發酵法生產多不飽和脂肪酸[J]. 中國生物工程雜志, 2007, 27(4): 139-145.
[11] SAKURADANI E, KOBAYASHI M, SHIMIZU S. Delta 6-fatty acid desaturase from an arachidonic acid-producing Mortierella fungus-gene cloning and its heterologous expression in a fungus,Aspergillus[J]. Gene. 1999, 238(2): 445-453.
[12] WAN Xia, ZHANG Yinbo, WANG Ping, et al. Production of gammalinolenic acid in Pichia pastoris by expression of a delta-6 desaturase gene from Cunninghamella echinulata[J]. Journal of Microbiology and Biotechnology, 2009, 19(10): 1098-1102.
[13] 李明春, 孫穎, 張琦, 等. 高山被孢霉Δ6脂肪酸脫氫酶基因在畢赤酵母中的胞內表達[J]. 生物工程學報, 2004, 20(1): 34-38.
[14] LIU Jianmin, LI Dianrong, YIN Yongtai, et al. Δ6desaturase from Mortierella alpina: cDNA cloning, expression, and phylogenetic analysis[J]. Biotechnology Letters, 2011, 33(10): 1985-1991.
[15] LI Yuntao, LI Maoteng, FU Chuhua, et al. Improvement of arachidonic acid and eicosapentaenoic acid production by increasing the copy number of the genes encoding fatty acid desaturase and elongase[J].Biotechnology Letters, 2009, 31(7): 1011-1017.
[16] 張光一, 李樹立, 劉增然. 食品級乳酸工程菌的研究進展[J]. 食品與發酵工業, 2005, 31(6): 73-75.
