不同植被恢復措施下剖面根系與SOC的分布特征
2013-08-17 03:50:10王彥麗
水土保持研究 2013年6期
關鍵詞:措施
王彥麗
(內蒙古鄂爾多斯市環境保護中心監測站,內蒙古 鄂爾多斯017000)
水土流失伴隨著SOC的流失[1],特別是在地形破碎、溝壑縱橫的黃土高原地區,嚴重的土壤侵蝕導致該地區的SOC密度和儲量均處于全國最低水平,甚至SOC含量與母質沒有差異[2]。水土流失治理可顯著提高SOC含量,據估計,全球水土流失治理的固碳潛力為1.47~3.04PgC/a[3]。植被恢復是治理水土流失的根本措施,一方面,植物根系通過對土壤的穿插、纏繞、網絡以及固結作用,增加了土壤的抗蝕性、抗沖性以及固定作用[4-5];另一方面,植物根系主要通過影響有機物向土壤中的歸還量[6-7]及其本身性質的差異[8]最終導致SOC的差異。根系作為聯系生態系統地上過程和地下過程的樞紐[9],影響著土壤中養分向植物體的輸送以及養分在土壤中的運移[10]。根系分布影響土壤剖面中碳的位置,而植被的地上地下部分的分配影響碳在剖面內的相對數量[11]。目前,大量的研究關注植被恢復措施(根系)在水土流失中的作用,但水土流失治理(植被恢復措施)后,根系生物量與SOC的剖面分布及二者的關系研究相對較少。
近年來,通過植被重建,治理水土流失、提高區域碳匯能力進而改善生態環境成為黃土高原地區生態環境建設的主要目標,但剖面內根系與SOC的分布特征以及二者的關系研究很少。因此,本研究基于黃土丘陵溝壑區典型小流域,在同一地形上(梁峁坡),選取不同的植被恢復措施,研究剖面內SOC與細根生物量的分布特征,以期為揭示根系對SOC影響的機理提供數據支持。
1 材料與方法
1.1 流域概況
燕溝流域(36°28′—36°32′N,109°20′—109°35′E)位于陜西省延安市寶塔區,主溝長8.6km,呈東南—西北流向,總面積約48km2,海拔986~1 425m,以梁峁地貌為主,溝壑密度為4.8km/km2,屬于典型的黃土丘陵溝壑區。研究區為暖溫帶半濕潤向半干旱過渡區,年平均氣溫9.8℃,多年平均降雨量約558 mm。土壤為半熟化狀態的黃綿土(約占90%以上),肥力較低。治理前(1997年以前)流域水土流失面積42.55km2,占總面積的88.65%,土壤侵蝕模數為9 000t/(km2·a),屬于強度水土流失地區[12]。
該區植被類型多樣,屬暖溫帶闊葉林向溫帶草原過渡地帶。人工草被主要為紫花苜蓿(MedicagosativaL),分布較少;人工灌木林主要為檸條(Caraganakorshinskii)、沙棘(Hippophaerhamnoides)、杠柳(PeriplocasepiumBge);人工喬木主要為刺槐(Robiniapseudoacacia)、山楊(Pobulusdavidiana)、油松(Pinustabulaeformis)、仁用杏(Prnuusarmeniacsibirica)、側柏(Biotaorientalis)。天然草被主要為鐵桿 蒿 (Artemisiagmelinii)、長 芒 草 (Stipa bungeana)、白羊草(Bothriochloaischaemum)、狗尾草(SetariaviridisL.)、黃蒿(Artemisiascoparia);天然灌木林主要為黃刺玫(Rosaxanthina)、狼牙刺(S.viciifdia)、虎榛子 (OstryopsisDavidianaDcne)、灰栒子(CotoneasteracutifoliusTurcz)、北京丁香(SyringapekinensisRupr);天然喬木林主要為遼東 櫟 (Queccusliaotungensis)、白 樺 (Betula platyphylla)、天然小喬木鼠李(Rhamnusdavurica)。其中,灌木林分布最廣。
1997年以來,在流域南部、中部、北部配置了不同的水土流失治理措施,南部主要以涵養水源的天然次生林為主,中部以人工水土保持植被為主,北部以農田和經濟林為主。
1.2 試驗設計與樣品采集
于2009年6月植被生長旺盛期進行野外樣品的采集,采樣深度定為1m,共采集了10個植被群落(表1),包括草本、灌木和喬木,分析其剖面內根系生物量(細根)和SOC的差異。為排除其他立地因子的干擾,所選樣地都在峁坡上,并保證其他環境因子基本一致。采樣時利用GPS進行定位,并記錄每個樣點的海拔、經緯度、地形地貌、植被特征、坡向、采樣點重復次數、采樣深度等信息。
每個樣地的土壤樣品各采3次重復,用直徑為3 cm的土鉆(根樣采5次重復,用直徑為9cm的根鉆)從地表豎直向下分別采集0—10,10—20,20—40,40—60,60—80,80—100cm土層的樣品。土壤樣品用自封袋收集,根樣用塑料袋收集,所有樣品帶回室內進行分析。

表1 燕溝流域樣地信息
1.3 室內分析
新鮮土樣混合均勻后置于陰涼處自然風干,風干樣磨細過0.25mm篩,測定SOC(H2SO4—K2Cr2O7外加熱法)[13]含量。挑出土壤中的所有根系,按處理和重復編號后放于塑封袋中,用水清洗干凈后,按直徑Φ≤2mm(細根)和Φ>2mm(粗根)分為兩級。將直徑小于2mm的根系用小紙袋裝好置于鼓風干燥箱中70~80℃烘干至恒重,再分別稱重和記錄。
1.4 數據處理與統計分析
利用Excel軟件進行數據的預處理;使用統計軟件SAS進行方差分析(Proc GLM程序包),當F檢驗顯著時再進行各樣地均值間的比較(Duncan法),用軟件Sigmaplot 10.0作圖。
2 結果與分析
2.1 不同治理措施下SOC和根系的分布
表2顯示了不同植被治理措施下SOC和細根密度的變化狀況。在0—100cm剖面內,不同治理措施下的SOC呈現出灌木>喬木>灌草>草本的趨勢(p<0.05)。以草本治理措施為對照,在0—10cm土層,灌 木 (12.13g/kg)、喬 木 (14.01g/kg)、灌 草(10.54g/kg)治理措施的SOC分別是草本治理措施(9.08g/kg)的1.34,1.56,1.16倍;在10—20cm土層,灌木(9.50g/kg)、喬木(8.30g/kg)、灌草(8.53 g/kg)治理措施的SOC分別是草本治理措施(7.57 g/kg)的1.26,1.10,1.13倍;在20—100cm土層,灌木(4.01~6.78g/kg)、喬木(3.76~6.43g/kg)、灌草(3.13~6.92g/kg)治理措施的SOC分別是草本治理措施(2.15~2.97g/kg)的1.56~2.28,1.47~2.16,1.45~2.33倍。
在0—100cm剖面內,不同治理措施下的細根密度呈現出喬木>灌木>灌草>草本的趨勢(p<0.05)。以草本治理措施為對照,在0—10cm土層,灌木 (1.78mg/cm3)、喬 木 (1.92mg/cm3)、灌 草(1.03mg/cm3)治理措施的細根密度分別是草本治理措施(1.30mg/cm3)的1.37,1.48,0.79倍;在10—20 cm土層,灌木(1.57mg/cm3)、喬木(1.95mg/cm3)、灌草(1.06mg/cm3)治理措施的細根密度分別是草本治理措施(0.33mg/cm3)的4.76,5.91,3.21倍;在20—100cm土層,灌木(0.18~1.08mg/cm3)、喬木(0.33~0.89mg/cm3)、灌草(0.16~0.57mg/cm3)治理措施的細根密度分別是草本治理措施(0.04~0.17mg/cm3)的1.96~7.09,4.00~8.13,1.45~5.00倍。
2.2 不同治理措施下SOC和根系的垂直分異
表2給出了不同治理措施下植被群落的SOC和細根密度在1m土層范圍內的垂直分布情況。SOC和細根生物量都隨土層深度的增加呈對數遞減趨勢(圖1),但同一土層SOC和細根生物量的分布不一致。
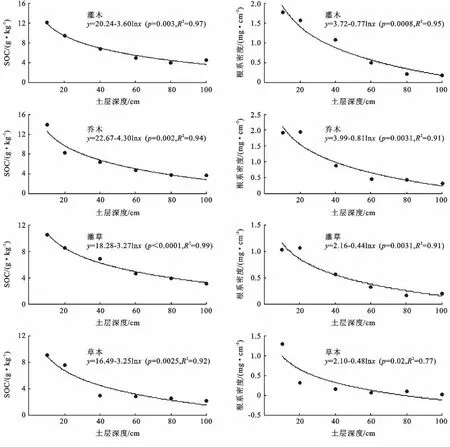
圖1 SOC和根系密度與土層深度的關系
在剖面內,以40—100cm土層的SOC和細根生物量為對照,在0—10cm土層,灌木、喬木、灌草和草本治理措施的SOC和細根生物量分別為底層(40—100cm)的2.69倍和2.00倍、3.40倍和1.55倍、2.72倍和1.50倍以及3.59倍和5.91倍;在10—20 cm土層,灌木、喬木、灌草和草本治理措施的SOC和細根生物量分別為底層(40—100cm)的2.11倍和1.76倍、2.02倍和1.58倍、2.20倍和1.55倍以及2.99倍和1.50倍;在20—40cm土層,灌木、喬木、灌草和草本治理措施的SOC和細根生物量分別為底層(40—100cm)的1.50倍和1.21倍、1.56倍和0.72倍、1.78倍和0.82倍以及1.17倍和0.77倍。
在0—10cm土層,灌木、喬木、灌草和草本治理措施的SOC和細根生物量分別占整個剖面的29%和33%,34%和32%,28%和31%以及38%和64%;在10—20cm土層,灌木、喬木、灌草和草本治理措施的SOC和細根生物量分別占整個剖面的23%和30%,20%和33%,23%和32%以及28%和16%;在20—40cm土層,灌木、喬木、灌草和草本治理措施的SOC和細根生物量分別占整個剖面的16%和20%,16%和15%,18%和17%以及11%和8%;但在40—100 cm土層,灌木、喬木、灌草和草本治理措施的SOC和細根生物量分別占整個剖面的32%和17%,30%和21%,31%和21%以及28%和11%。
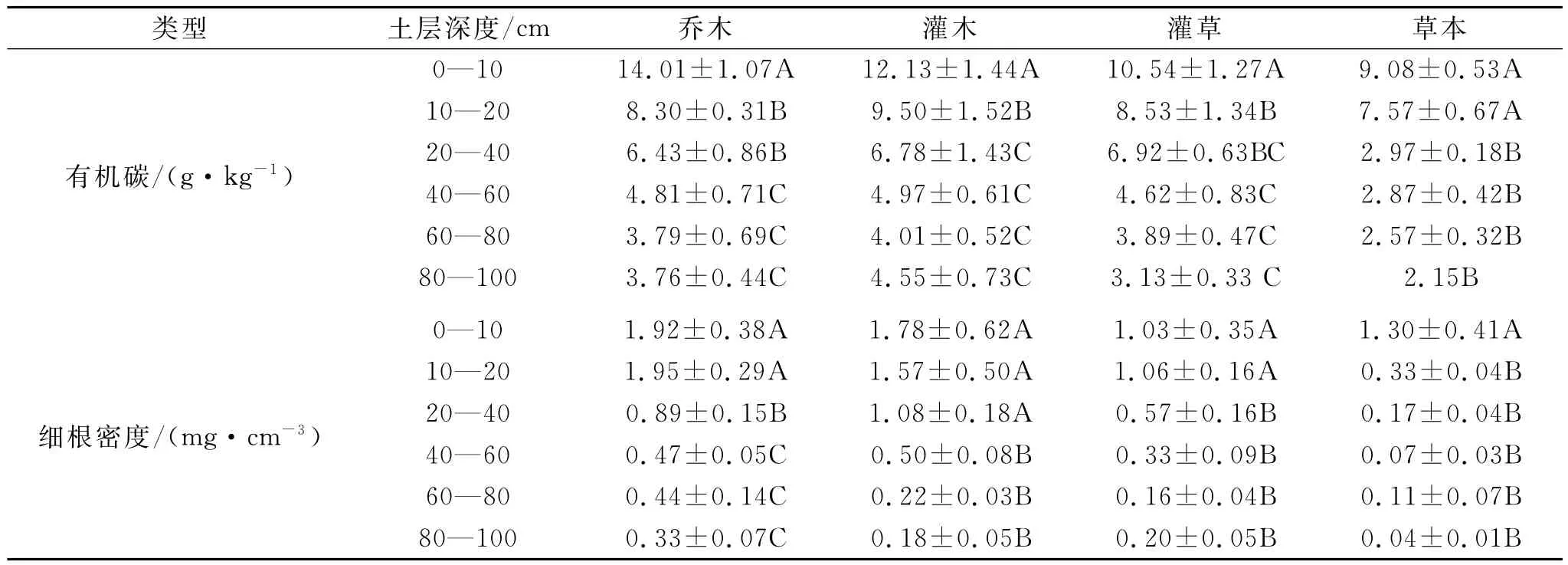
表2 不同土層有機碳和根系的差異
3 討 論
3.1 根系生物量對土壤有機碳的影響
植被根系一方面能減少水土流失[14-15],另一方面顯著地影響土壤中有機碳的累積,根系對SOC的影響主要表現在:不同植被治理措施歸還到土壤中的根系生物量不同,在0—100cm剖面內,灌木、喬木以及灌草恢復措施的根系生物量分別是草本恢復措施根系生物量的4.3倍、5.2倍以及3.1倍(表2),這與已有的研究結果類似[6-7];不同根系的生物化學性狀不同,從而使得有機物的分解速率不同[16-17]。例如,林草地根系的降解性要難于農田,因為凋落物分解的難易程度與自身的C∶N呈反相關[16];地下根系的生化特征會因植被類型的改變而發生顯著的變化,例如,玉米根系中的纖維素和木質數含量分別為49%和12%,黑麥草中則分別為30%和9%[17]。根系分泌物和細根的周轉是SOC的主要來源[16,18],但不是唯一的來源,同時土壤中的有機碳的累積還受其他環境因素和生物因素的制約,這可以從根系生物量與SOC的對數關系得到驗證(圖2)[19],即隨著根系密度的增加土壤有機碳呈對數增加趨勢,但不同植被恢復措施的增加程度不同,且呈現出喬木措施(單位根系密度的轉化系數為4.38)>灌草措施(單位根系密度的轉化系數3.91)>灌木措施(單位根系密度的轉化系數為2.87)>草本措施(單位根系密度的轉化系數2.33)的趨勢。隨著根系輸入到土壤中的有機物質增加,SOC不斷積累,越利于植被的生長,從而能更有效地減少水土流失和改善生態環境,特別在水土流失嚴重和生態環境惡化的黃土高原地區顯得尤為重要。
3.2 土壤有機碳和根系的垂直分異
根系作為SOC的主要來源[18],其在剖面的分布及數量決定著SOC的分布以及數量,但根系生物量不是決定SOC分布以及數量的唯一因素,因而研究發現SOC和根系在剖面的分布并不一致(表1),例如,Weaver等[20]發現,在0—15cm的土層中有機質含量占整個剖面(0—100cm)的40%,但相應土層中的根系生物量卻占整個剖面生物量的70%;同時Gill[21]研究也發現,在0—100cm 的剖面中,根系生物量的75%處于0—15cm處,而相應的有機質卻只占23%;盡管根系與SOC在剖面的分布并不一致,但王一冬[19]在我國內蒙古地區研究得出,在0—300cm的土壤剖面中,根系生物量密度與SOC含量呈正的線性相關(R2=0.62),但與SIC呈負的指數相關;但也有學者研究得出土壤有機碳在剖面內的分布不僅決定著根系的分布[20],植被類型的自身性質或者降水也可以決定其分布[19]。已有的研究表明植被的根系分布比土壤有機碳的分布淺[13]。
無論是SOC還是根系生物量,均呈現出上層(0—20cm)大于下層(20—100cm)的趨勢,這與許多研究者得出的結論一致[22],這可能與剖面內微生物的分布、數量、結構有關,還有可能與剖面內土壤中的養分狀況以及水分可利用性密切相關,但具體原因有待進一步研究。

圖2 SOC和根系密度的關系
根系生物量和SOC在不同植被治理措施下呈現出明顯差異,無論是根系生物量還是SOC均呈現出表層大于深層的趨勢,且均隨土層深度的增加呈對數遞減趨勢;SOC和根系在剖面的分布不一致,并且根系比SOC的分布淺,但二者呈對數增加趨勢。
[1] 畢銀麗,王百群.黃土丘陵區壩地系統土壤養分特征及其及侵蝕環境的關系:Ⅰ.壩地土壤的理化性狀及其數值分析[J].土壤侵蝕與水土保持學報,1997,3(3):1-9.
[2] 楊文治,余存祖.黃土高原區域治理與評價[J].北京:科學出版社,1992.
[3] Lal R.Global soil erosion by water and carbon dynamics[M]∥Lal R,Klmbkle J,Levine E.Soil and Global Change.Boca Raton F L:Lewis Publishers,1995.
[4] 毛瑢,孟廣濤,周躍.植物根系對土壤侵蝕控制機理的研究[J].水土保持研究,2006,13(2):241-243.
[5] 李鵬,李占斌,鄭良勇.植被保持水土有效性研究進展[J].水土保持研究,2002,9(1):76-80.
[6] French N R,Steinhorst R K,Swift D M.Grassland biomass trophic pyramids[J].Perspectives in Grassland Ecology,Ecol.,Studies,1979,32:59-87.
[7] 沈善敏.無機氮對土壤氮礦化與固定的影響:兼論土壤氮的“激發效應”[J].土壤學報,1986,23(1):10-16.
[8] Rovira P,Vallejo V R.Labile and recalcitrant pools of carbon and nitrogen in organic matter decomposing at different depths in soil:An acid hydrolysis approach[J].Geoderma,2002,107(1):109-141.
[9] Trumbore S E,Gaudinski J B.The secret lives of roots[J].Science,2003,302(5649):1344-1345.
[10] 韓鳳朋,鄭紀勇,張興昌.黃土退耕坡地植物根系分布特征及其對土壤養分的影響[J].農業工程學報,2009,25(2):50-55.
[11] Davidson E A,Savage K,Bostad P,et al.Belowground carbon allocation in forests estimated from litterfall and IRGA-based soil respiration measurements[J].Agricultural and Forest Meteorology,2002,113:39-51.
[12] Schenk H J,Jackson R B.Rooting depths,lateral root spreads and below-ground/above-ground allometries of plants in water-limited ecosystems[J].Journal of Ecology,2002,90(3):480-494.
[13] Jobbagy E G,Jackson R B.The vertical distribution of soil organic carbon and its relation to climate and vegetation[J].Ecological Applications,2000,10(2):423-436.
[14] 胡海波,魏勇,仇才樓.蘇北沿海防護林土壤可蝕性的研究[J].水土保持研究,2001,8(1):150-154.
[15] 楊永紅,劉淑珍,王成華,等.含根量與土壤抗剪強度增加值關系的試驗研究[J].水土保持研究,2007,14(3):287-289.
[16] Szanser M,Ilieva-Makulec K,Kajak A,et al.Impact of litter species diversity on decomposition processes and communities of soil organisms[J].Soil Biology &Biochemistry,2011,43(1):9-19.
[17] Fernandez I,Mahieu N,Cadisch G.Carbon isotopic fractionation during decomposition of plant materials of different quality[J].Global Biogeochemical Cycles,2003,17(3):1-9.
[18] Matamala R,Gonzàlez-Meler M A,Jastrow J D,et al.Impacts of fine root turnover on forest NPP and soil C sequestration potential[J].Science,2003,302(5649):1385-1387.
[19] Wang Y G,Li Y,Ye X H,et al.Profile storage of organic/inorganic carbon in soil:From forest to desert[J].Science of the Total Environment,2010,408(8):1925-1931.
[20] Weaver J E,Hougen V H,Weldon M D.Relation of root distribution to organic matter in prairie soil[J].Botanical Gazette,1934,96(3):389-420.
[21] Gill R,Burke I C,Milchunas D G,et al.Relationship between root biomass and soil organic matter pools in the shortgrass steppe of eastern Colorado[J].Ecosystems,1999,2(3):226-236.
[22] 趙忠,李鵬.渭北黃土高原主要造林樹種根系分布特征及抗旱性研究[J].水土保持學報,2002,16(1):96-99.
猜你喜歡
今日畜牧獸醫(2022年10期)2022-12-23 06:23:28
今日畜牧獸醫(2022年10期)2022-12-23 06:19:54
今日農業(2021年9期)2021-11-26 07:41:24
建材發展導向(2021年15期)2021-11-05 08:22:20
今日農業(2020年13期)2020-12-15 09:08:51
湖北農機化(2020年4期)2020-07-24 09:07:10
甘肅教育(2020年8期)2020-06-11 06:09:48
中學生數理化(高中版.高考理化)(2020年3期)2020-05-30 12:26:24
中國外匯(2019年21期)2019-05-21 03:04:06
豬業科學(2018年4期)2018-05-19 02:04:38
