日本血吸蟲TOM34基因的克隆和表達分析
2013-08-21 06:51:56李學珍傅志強石耀軍楊健美朱傳剛苑純秀李祥瑞林矯矯
中國人獸共患病學報 2013年2期
洪 煬,李學珍,,傅志強,石耀軍,陸 珂,楊健美,朱傳剛,苑純秀,李 浩,李祥瑞,林矯矯
2.南京農(nóng)業(yè)大學動物醫(yī)學院,南京 210095
血吸蟲病(schistosomiasis)是由血吸蟲感染人或其他哺乳動物而導致的一種分布廣泛、危害嚴重的人獸共患寄生蟲病(zoonosis)。該病流行于全球的76個國家和地區(qū),有約6億人受到威脅,2億人感染[1]。根據(jù)我國衛(wèi)生部統(tǒng)計:2010年血吸蟲病仍在我國3 520個鄉(xiāng)鎮(zhèn)流行,患病人數(shù)達32.58萬人[2]。目前,血吸蟲病的防治主要依賴于化學藥物吡喹酮的使用,但是這種方法在血吸蟲病未消滅地區(qū)只能暫時降低發(fā)病率,不能徹底阻斷該病的傳播。同時,藥物治療對已造成損傷的組織無法修復,并且不能防止病人、病畜再次感染。近年來,在曼氏血吸蟲和日本血吸蟲中都已經(jīng)出現(xiàn)了吡喹酮的耐藥蟲株[3-4]。吡喹酮是目前唯一大規(guī)模使用的血吸蟲病治療藥物,耐藥蟲株的出現(xiàn)引起了人們的高度關注。因而加強抗血吸蟲疫苗和新治療藥物的研發(fā)格外重要。
研究表明,日本血吸蟲在不同適宜性宿主體內(nèi)的生存狀態(tài)是不同的,這可能是因為不同的宿主為日本血吸蟲提供了不同的生存環(huán)境,從而影響到蟲體的生存和發(fā)育。本課題組近期利用熒光差異凝膠雙向電泳對來源于日本血吸蟲易感宿主小鼠、非易感宿主大鼠和抗性宿主東方田鼠10 d童蟲的差異表達蛋白進行了比較分析,結(jié)果顯示不同適宜性宿主來源童蟲的蛋白表達存在差異,其中一些差異表達的蛋白可能是蟲體生存發(fā)育所需要的關鍵分子[5]。本文選擇其中一個差異表達蛋白—線粒體外膜轉(zhuǎn)運酶34(SjTOM34)進行了初步的研究。
線粒體是真核細胞中能量代謝的重要場所,它除了產(chǎn)生ATP外,還可以代謝氨基酸、脂類和鐵硫簇等其他重要的化合物。此外,線粒體還參與諸如細胞分化、細胞信息傳遞和細胞凋亡等過程,并擁有調(diào)控細胞生長和細胞周期的能力。線粒體中含有的蛋白種類大約占整個細胞的25%,在哺乳動物細胞中大約為1 500種,這些蛋白中大約有99%是在細胞質(zhì)中合成之后被轉(zhuǎn)運至線粒體內(nèi)的[6]。在這個過程中,線粒體的外膜轉(zhuǎn)運酶(translocase of the outer mitochondrial membrane,TOM)發(fā)揮了重要的作用,它們通過主動運輸?shù)姆绞綄⑦@些線粒體前體蛋白運輸?shù)骄€粒體內(nèi)[7]。近幾年,國外許多學者對TOM34進行了相關研究,認為該蛋白在胞質(zhì)中大量分布,在線粒體膜上也有部分分布,并且認為它在線粒體前體蛋白進入線粒體的過程中發(fā)揮著重要的作用[8-9]。盡管 TOM34在哺乳動物細胞中已有一些相關研究,但有關血吸蟲TOM34蛋白國內(nèi)外尚未見報道。
本研究以SjTOM34為研究對象,對該基因進行了克隆、表達及相關的生物信息學分析,制備了針對該基因的多克隆抗體,分析了該基因在不同時期蟲體內(nèi)mRNA水平和蛋白水平上的表達情況,對該基因在蟲體內(nèi)的分布情況進行了免疫定位,為進一步研究該基因的生物學功能及作用機制奠定了基礎。
1 材料與方法
1.1 材料
1.1.1 寄生蟲、質(zhì)粒、菌種及實驗動物 日本血吸蟲中國大陸株尾蚴由中國農(nóng)業(yè)科學院上海獸醫(yī)研究所釘螺室提供;大腸埃希氏桿菌(E.coli)DH5α、BL21和質(zhì)粒p ET28a(+)均為本實驗室保存;6 w齡的雌性BALB/c(20~25 g)小鼠購自上海斯萊克實驗動物中心,雄性新西蘭大白兔(2.5 kg~3.0 kg)購自上海羅涇飛達實驗動物養(yǎng)殖場。
1.1.2 相關試劑 TRIzol購自Invitrogen公司;RNA提取試劑盒、SYBR GreenⅠ、EASY Dilution、p MD19-T載體、T4 DNA連接酶、限制性內(nèi)切酶、DNA marker(DL2000,DL15000)購自 Ta KaRa生物工程(大連)有限公司;瓊脂糖凝膠電泳DNA純化回收試劑盒、質(zhì)粒快速提取試劑盒購自Axygen公司;Ni-NTA HisBind Resin購自Novagen公司;沉淀型單組分TMB底物溶液、可溶性單組分TMB底物溶液購自天根生物科技(北京)有限公司;硝酸纖維素膜(NC膜)購自Whatman公司;辣根過氧化物酶標記的山羊抗小鼠IgG二抗、Cy3標記的山羊抗小鼠IgG二抗、DAPI溶液購自碧云天生物技術有限公司。
1.2 方法
1.2.1 蟲體收集 新西蘭大白兔腹部貼片感染3 000~20 000條日本血吸蟲尾蚴,于感染后7 d、14 d、21 d、28 d、35 d和42 d分別剖殺,以灌注法從肝門靜脈收集蟲體,用滅菌的PBS充分沖洗蟲體。用吸管將部分42 d合抱的成蟲反復吹打,使合抱的雌雄蟲分開,分別收集雌蟲和雄蟲,將各時期的蟲體保存于液氮中備用。
1.2.2 總RNA及蟲體蛋白的提取 取液氮中保存的7 d、14 d、21 d、28 d、35 d和42 d日本血吸蟲,按TRIzol試劑盒說明書分別提取總RNA,將42 d的RNA反轉(zhuǎn)錄為c DNA,-80℃保存?zhèn)溆谩?/p>
取液氮中保存的7 d、14 d、21 d、28 d、35 d和42 d日本血吸蟲,各稱取相同重量20 mg,加入250μL RIPA裂解液,用研磨棒對蟲體進行充分研磨。隨后加入苯甲基磺酰氟化物(PMSF),使其最終濃度為1 mmol/L。冰上超聲5次,每次超聲5 s,停15 s。之后在4℃下,12 000 r/min離心20 min,吸取上清放于-80℃?zhèn)溆谩?/p>
從行走機構(gòu)和翻轉(zhuǎn)機構(gòu)的模態(tài)振型圖可以看出,各個頻率的振型差別較大。各階模態(tài)振型可分為整體模態(tài)和局部模態(tài)。其中,第一階、第三階和第四階為局部模態(tài),第二階和第五階為整體模態(tài),第六階為整體扭曲變形模態(tài)。各階模態(tài)振型說明如表2所示。
1.2.3SjTOM34序列的擴增及生物信息學分析
本實驗室之前對來源于小鼠、大鼠、東方田鼠的日本血吸蟲10 d童蟲差異表達蛋白的研究中,獲得了一個EST序列(登錄號:FN315604.1),根據(jù)該EST序列設計上下游引物,進行PCR擴增。上游引物:5′-ATGAAACCTGAACAGATTTCCATGCTC-3′,下 游 引 物:5′-CTAGTCTAAGTCTGGACATGATTGGTTAG-3′。以日本血吸蟲35 d成蟲cDNA作為模板進行PCR擴增,PCR反應條件為94℃預變性5 min;然后94℃30 s,55℃30 s,72℃90 s,共30個循環(huán);循環(huán)結(jié)束后72℃延伸10 min,將PCR產(chǎn)物純化后送上海英俊生物技術有限公司進行測序。
將測序得到的c DNA序列利用DNAStar軟件分析該序列的開放閱讀框,氨基酸殘基數(shù)目、組成,理論相對分子質(zhì)量及等電點,同時在美國國家生物技術信息中心(NCBI)利用 BLAST 軟件(http://www.ncbi.nlm.nih.gov/BLAST)進行序列的相似性和同源性分析,通過Clustal X軟件將蛋白序列進行多重比對,繪制出系統(tǒng)進化樹。利用SignalP軟件(http://www.cbs.dtu.dk/services/SignalP/)對蛋白的信號肽進行預測;利用Ex PASy服務器上的ProtScale 軟 件 (http://us.expasy.org/cgi-bin/protscale.pl)對蛋白序列進行疏水性分析;利用Wo LF PSORT 軟件(http://wolfpsort.org/)對該蛋白的亞細胞定位進行分析。
1.2.4 熒光實時定量PCR 選擇血吸蟲看家基因tubulin作為內(nèi)參。將提取的7 d、14 d、21 d、28 d、35 d、42 d蟲體總RNA,去除基因組DNA后反轉(zhuǎn)錄為cDNA,采用熒光染料法進行實時定量PCR檢測。利用Beacon Designer軟件設計Sj TOM34和tubulin基因的實時定量PCR引物,其中SjTOM34實時定量PCR的上游引物為5′-GCCGTCTCATGCCATAGC-3′,下 游 引 物 為 5′-AGCATCGTCTGTGTAGTCTAAC-3′,擴增片段為163 bp;tubulin實時定量PCR的上游引物為5′-CTGATTTTCCATTCGTTTG-3′,下游引物為5′-GTTGTCTACCATGAAGGCA-3′,擴增片段為213 bp。PCR 反應體系為:2×SYBR Premix Ex TaqTM10μL,10 μmol/L上游引物0.4μL,10μmol/L下游引物0.4 μL,dd H2O 8.2μL,cDNA模板1μL,共20μL。反應參數(shù)為:50℃2 min,95℃10 s,(95℃5 s;60℃34 s)40個循環(huán),其中60℃34 s結(jié)束時間為熒光信號檢測點,每個反應均進行3個重復。采用Mastercyclerep Realplex軟件進行分析,分別計算出7 d、14 d、21 d、28 d、35 d、42 d雌蟲和42 d的雄蟲中SjTOM34相對于tubulin的含量。
1.2.5 重組表達質(zhì)粒的構(gòu)建 根據(jù)GenBank中的EST序列信息,設計該基因的擴增引物進行cDNA片段的擴增。上游引物:5′-CGCGTCGACATGAAACCTGAACAGATTT-3′,下劃線處為SacI位點;下 游 引物:5′-TCCTCGAGCTAGTCTAAGTCTGGACATG-3′,下劃線處為XhoI位點。以反轉(zhuǎn)錄得到的35 d蟲體的cDNA為模板進行PCR擴增,得到引入SacI和XhoI酶切位點的SjTOM34基因ORF序列。將測序正確的序列定向克隆至原核表達載體p ET28a(+),構(gòu)建重組表達質(zhì)粒p ET28a(+)-SjTOM34,并轉(zhuǎn)化至大腸桿菌BL21(DE3)感受態(tài)細胞中。
1.2.6 重組蛋白的表達與純化 將測序正確的p ET28a(+)-SjTOM34/BL21接種于含有卡那霉素的液體LB培養(yǎng)基中,37℃振蕩培養(yǎng),至OD600為0.6時,加入終濃度為1 mmol/L的IPTG進行誘導表達。應用SDS-PAGE電泳分析最佳誘導時間及表達形式后,通過Ni-NTA His Bind Resin對重組蛋白進行純化及復性。
1.2.7 抗體的制備 選擇6 w BALB/c進行皮下注射免疫,每次每只注射206佐劑和20μg純化的重組SjTOM34蛋白的乳化液,其中重組蛋白SjTOM34與206佐劑以體積比46∶54的比例進行混合乳化。免疫間隔時間為2周,共免疫3次,第3次免疫后2周剖殺小鼠,收集血清,-20℃保存?zhèn)溆谩?/p>
2 結(jié) 果
2.1SjTOM34的克隆及生物信息學的分析 在本實驗室10 d童蟲差異性表達蛋白質(zhì)組學研究基礎上,選擇了一個EST序列進行進一步的研究。應用RT-PCR的方法擴增得到了一個與預期大小相符的特異基因片段,測序結(jié)果顯示,NCBI上公布的序列在797位多了一個核苷酸“A”,從而使ORF的大小發(fā)生了改變。DNAStar分析表明該基因的ORF大小為1 083 bp,編碼360個氨基酸,理論分子量為40.8 k D,理論等電點為4.94。該氨基酸序列的N端無信號肽,疏水區(qū)不明顯。
將來自曼氏血吸蟲(GenBank登錄號ref|XP_002577521.1)、華支睪吸蟲(GenBank登錄號:dbj|GAA28250.1)、蜥蜴(GenBank登錄號:ref|NP_003221034.1)、原雞 (GenBank 登錄 號:ref|XP_001025928.1)、穴 兔 (GenBank 登 錄 號:XP_002710718.1)、黑 猩 猩 (GenBank 登 錄 號:XP_003311967.1)和 人 (GenBank 登 錄 號:gb|AAB38382.1)7個物種的TOM34蛋白序列進行多重比對,結(jié)果顯示該基因編碼的蛋白序列與曼氏血吸蟲TOM34蛋白的氨基酸序列的相似性為78%,與華支睪吸蟲的相似性為57%,與蜥蜴、原雞、穴兔、黑猩猩、人的相似性分別為44%、44%、42%、40%、42%(見圖1、圖2)。
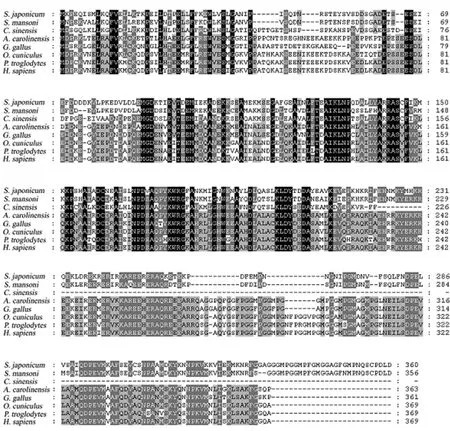
圖1 不同物種TOM34蛋白序列的多重比對Fig.1 Comparison of the protein sequence of Sj TOM34 with those of the other species
2.2SjTOM34基因的期別、性別差異表達分析Real-time RT-PCR結(jié)果表明,SjTOM34基因在所檢測的童蟲及成蟲各階段蟲體中均有表達,其中在35 d蟲體的表達量最高,在其他時期蟲體中的表達量較為接近,在42 d雌蟲和雄蟲中的表達無顯著差別(圖3)。
2.3 重組原核表達質(zhì)粒p ET28a(+)-SjTOM34的構(gòu)建 經(jīng)雙酶切鑒定(圖4)和測序鑒定后,證實了p ET28a(+)-SjTOM34重組表達質(zhì)粒的構(gòu)建成功。
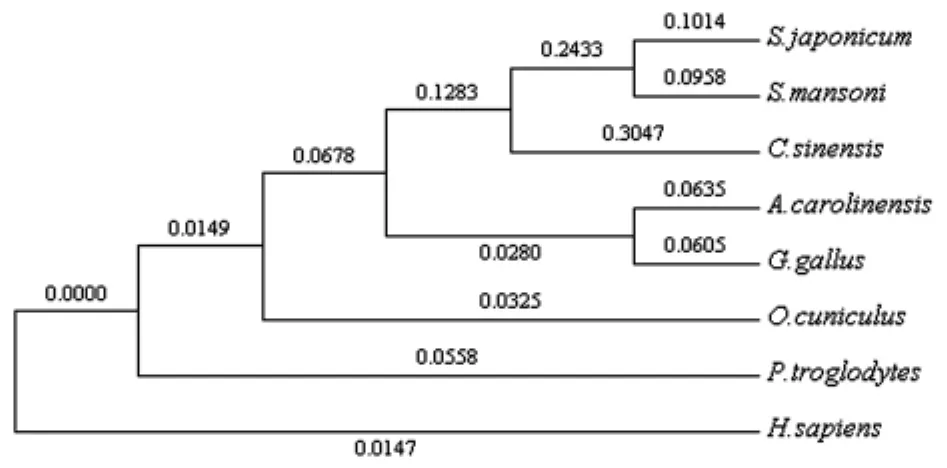
圖2 不同物種TOM34蛋白的進化樹分析Fig.2 Phylogenetic tree analysis of Sj TOM34 with its homologues
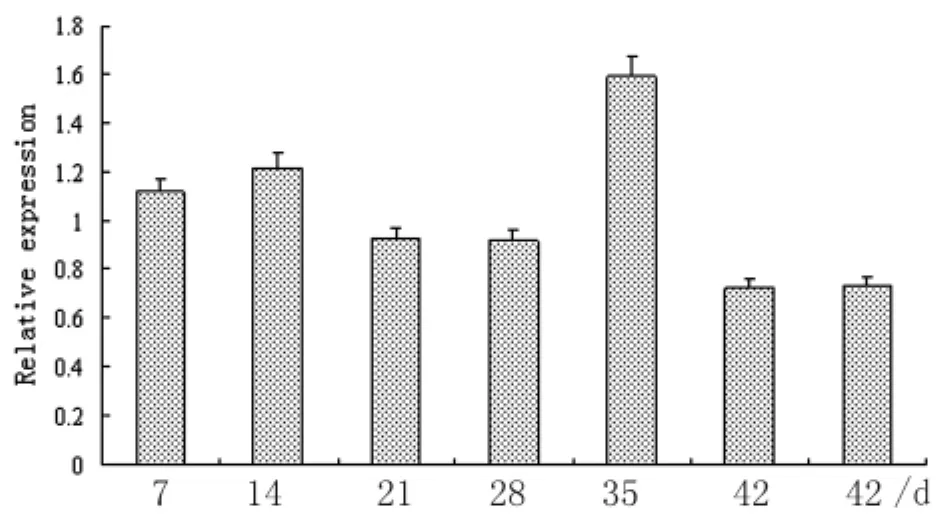
圖3 實時定量PCR檢測Sj TOM34基因在日本血吸蟲不同期別、性別蟲體中的表達情況Fig.3 Stage and gender differential expression of Sj TOM34 in S.japonicum by real-time RT-PCR7 d,14 d,21 d,28 d,and 35 d represent worms at 7 days,14 days,21 days,28 days,and 35 days,respectively.42 d(f):female adult worms at 42 days;42 d(m):male adult worms at 42 days.
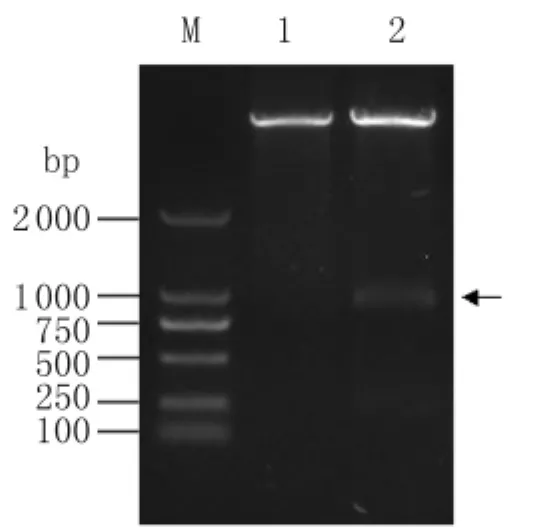
圖4 重組表達質(zhì)粒pET28a(+)-Sj TOM34的雙酶切鑒定Fig.4 The recombinant plasmid identified by enzyme digestionM:DL2000 marker;1:p ET28a(+)digested with Sal I and Xho I restriction enzyme;2:p ET28a(+)-Sj TOM34 digested with Sal I and Xho I restriction enzyme
2.4 pET28a(+)-SjTOM34的原核表達和重組蛋白的純化 SDS-PAGE結(jié)果顯示,重組原核表達質(zhì)粒p ET28a(+)-SjTOM34在大腸桿菌BL21中成功表達,重組蛋白的分子量約為45 k D,與預期結(jié)果相符。該重組蛋白在超聲上清和沉淀中都有存在,經(jīng)Ni-NTA樹脂純化后,可獲得較純的重組蛋白(圖5)。
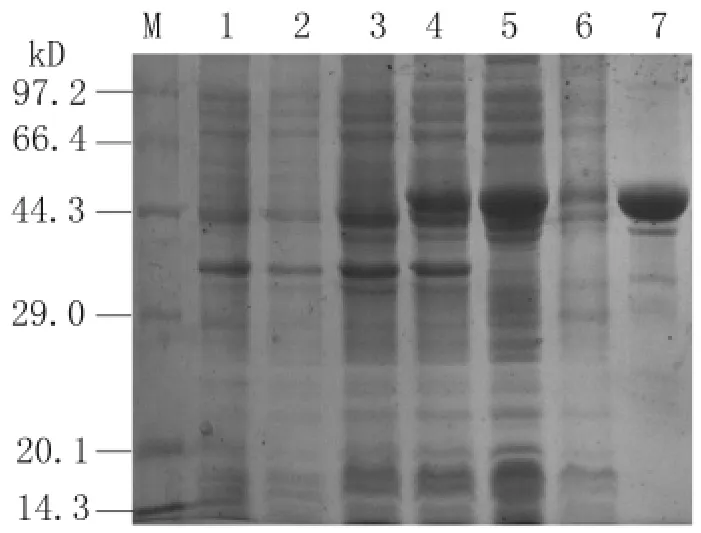
圖5 SDS-PAGE分析pET28a(+)-Sj TOM34/BL21的蛋白表達情況Fig.5 SDS-PAGE analysis of the expression of pET28a(+)-Sj TOM34/BL21 in E.coli.M:Protein marker;Lane 1 and 2:Total extract of p ET28a(+)before and after induction with 1 m M IPTG;Lane 3 and 4:Expression of recombinant plasmid p ET28a(+)-Sj TOM34 before and after induction with 1 m M IPTG;Lane 5 and 6:Pellet and supernatant of expression product after lysis,respectively;Lane 7:Sj TOM34 purified through Ni2+-charged column chromatography and after dialysis.
2.5 重組蛋白的抗原性分析 通過Western blotting檢測,在45 k D呈現(xiàn)一明顯的識別條帶(圖6),證明該重組蛋白具有良好的抗原性。對SjTOM34基因在不同期別蟲體內(nèi)蛋白表達水平的Western blotting檢測顯示,該蛋白在不同時期蟲體內(nèi)都存在(圖7)。
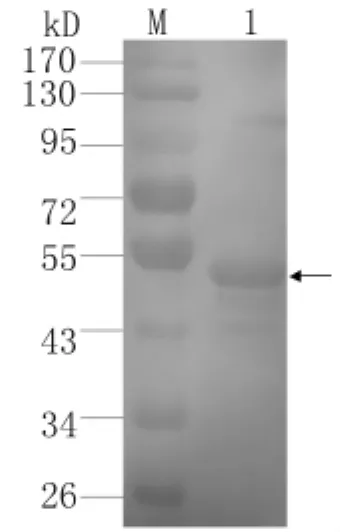
圖6 重組蛋白Sj TOM34的Western blotting分析Fig.6 Western blotting analysis of pET28a(+)-Sj TOM34M:Prestained protein marker;Lane 1:Purified recombinant Sj TOM34 protein was recognized with mouse serum against Sj TOM34.
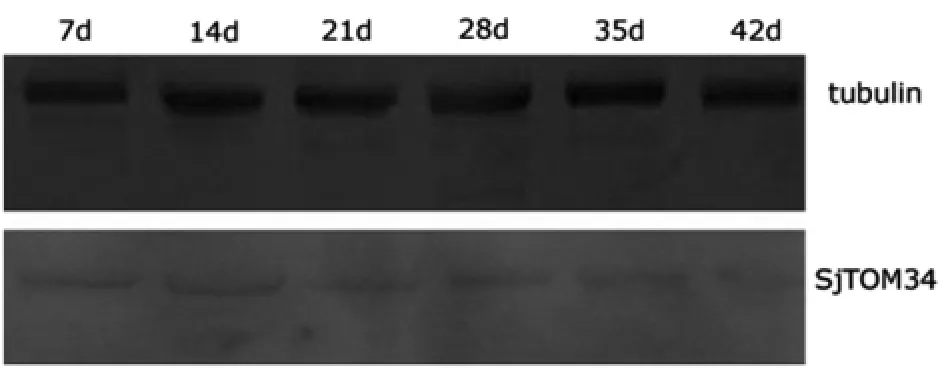
圖7 不同時期蟲體中Sj TOM34蛋白的Western blotting分析Fig.7 Western blotting analysis of the protein extracts from S.japonicum at different stages7 d,14 d,21 d,28 d,35 d and 42 d represent worms at 7 days,14 days,21 days,28 days,35 days and 42 days,respectively.
2.6SjTOM34蛋白在蟲體內(nèi)的免疫組化分析在熒光顯微鏡下可以觀察到Cy3標記的山羊抗小鼠IgG二抗發(fā)出的紅色熒光以及DAPI復染核酸后發(fā)出的藍色熒光,實驗結(jié)果表明,該蛋白廣泛分布在蟲體內(nèi)部的實質(zhì)組織中(圖8)。
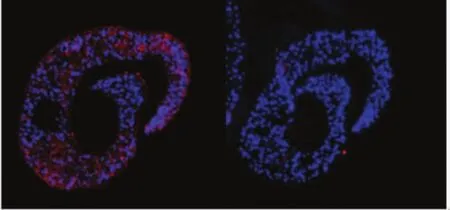
圖8 Sj TOM34蛋白在21 d日本血吸蟲蟲體內(nèi)的定位分析(10×40)Fig.8 The localization analysis of Sj TOM34 by immunofluorescence in 21-day-old worms(10×40)
3 討 論
在我國,日本血吸蟲可以感染40多種哺乳動物,山羊、黃牛、綿羊、兔子和小鼠等都是其易感宿主,另外一些哺乳動物如大鼠等是日本血吸蟲的不易感宿主。在不易感宿主體內(nèi),血吸蟲的發(fā)育率低而且蟲體較小[10-11]。東方田鼠是目前為止發(fā)現(xiàn)的唯一一種對日本血吸蟲具有抗性的宿主,日本血吸蟲尾蚴可以感染東方田鼠,感染之后蟲體發(fā)育遲緩,在感染尾蚴15 d后,東方田鼠體內(nèi)基本上找不到存活的蟲體[12-13]。不同的宿主為日本血吸蟲提供不同的生存環(huán)境,影響到蟲體的生存和發(fā)育。不同宿主環(huán)境中生長的蟲體在蛋白表達上也存在一定的差異,而這些差異表達的蛋白可能是蟲體生存發(fā)育所需要的關鍵分子。在本實驗室之前的研究中發(fā)現(xiàn),SjTOM34蛋白在小鼠來源10 d童蟲中的含量明顯高于大鼠和東方田鼠來源的10 d童蟲的含量,因此對這種在不同易感性宿主環(huán)境中日本血吸蟲生長發(fā)育差異的蛋白作進一步的研究具有理論意義,同時也可為血吸蟲新藥靶和疫苗候選分子篩選提供基礎。
TOM34是一種在哺乳動物細胞內(nèi)發(fā)現(xiàn)的線粒體外膜轉(zhuǎn)運結(jié)構(gòu)蛋白,該蛋白包含3個TPR結(jié)構(gòu),通過TPR結(jié)構(gòu)識別前體蛋白,與前體蛋白的成熟區(qū)域相結(jié)合,進而調(diào)節(jié)前體蛋白進出入線粒體。Chewawiwat等[8]發(fā)現(xiàn)在哺乳動物細胞中TOM34大量存在于胞質(zhì)中,線粒體膜上也有部分存在,并且TOM34是線粒體外膜輸入機制的重要組成成分,在線粒體前體蛋白進入線粒體的過程中發(fā)揮著重要的作用。同時他們還發(fā)現(xiàn),在體外抗TOM34的抗體和缺乏N端疏水序列的TOM34能強烈阻止前體蛋白進入大鼠的線粒體內(nèi)。Mukhopadhyay等[9]認為重組表達的人TOM34蛋白能夠與胞質(zhì)中線粒體前體蛋白的成熟區(qū)域發(fā)生相互作用,使這些蛋白能夠保持一種未折疊的狀態(tài),從而能夠更容易地進入線粒體內(nèi),但是它不能與前體蛋白的引導序列結(jié)合。Yang等[14]利用酵母雙雜交技術篩選能夠與人TOM34結(jié)合的蛋白,結(jié)果發(fā)現(xiàn)胞質(zhì)中的TOM34首先與ATP酶相關的纈酪肽包含蛋白(VCP)和溶酶體H+運輸ATP酶膜蛋白 M(ATP6M)相結(jié)合,它們能夠形成一個400 k D的復合物,而201-256的TPR基序能夠與其它蛋白相結(jié)合,并且他們認為TOM34似乎不是線粒體外膜轉(zhuǎn)移蛋白家族中的一員,而是在蛋白轉(zhuǎn)移至線粒體過程中具有伴侶樣功能的蛋白分子。Shimokawa等[15]通過cDNA芯片發(fā)現(xiàn),TOM34基因在人的結(jié)腸癌細胞中表達上調(diào),對結(jié)腸癌細胞HCT116的TOM34基因進行RNA干擾后,能夠有效的抑制該基因的表達并且抑制該細胞的生長,其可以作為一種潛在的診斷抗原或抗結(jié)腸癌的藥靶。Faou等[6]發(fā)現(xiàn),TOM34能夠作為一個伴侶分子與熱激蛋白70(Hsp70)和熱激蛋白90(Hsp90)形成一個胞質(zhì)復合物,從而使線粒體前體蛋白能夠進入線粒體,當TOM34過量、Hsp70和Hsp90不足的時候,線粒體的前體蛋白進入線粒體的數(shù)量是不會增加的,并且TOM34具有結(jié)合以TOM70轉(zhuǎn)移途徑進入線粒體的前體蛋白的能力。上述的這些研究結(jié)果都說明了TOM34在前體蛋白進入線粒體的過程中以及對細胞正常的生長都具有重要的作用。
本實驗所克隆的SjTOM34基因經(jīng)過生物信息學分析,發(fā)現(xiàn)該序列未含有信號肽,在胞質(zhì)和細胞核中都可能存在,這與之前報道的哺乳動物TOM34蛋白所在位置的結(jié)果較為一致。而同源性比對和系統(tǒng)進化樹的分析結(jié)果顯示,日本血吸蟲TOM34與曼氏血吸蟲的同源性較高,而與人和其它哺乳動物的同源性較低,這為將來以該蛋白作為新藥物靶標或疫苗候選分子的可能性提供了一定的理論基礎。通過實時定量和Western blotting分析發(fā)現(xiàn),TOM34基因在日本血吸蟲各個發(fā)育階段都有表達且表達水平相對穩(wěn)定,這可能與該蛋白在轉(zhuǎn)移線粒體蛋白進入線粒體內(nèi)這一過程中的重要作用有關。而本實驗室之前研究發(fā)現(xiàn)日本血吸蟲童蟲在非適宜和抗性宿主中生長狀況不好,可能與該蛋白在這兩種宿主來源蟲體中的含量低,影響了寄生蟲正常的蛋白合成與代謝有一定關系。同時 Western blotting的結(jié)果表明該重組表達產(chǎn)物具有良好的免疫原性,可以誘導BALB/c小鼠特異性抗體的產(chǎn)生。而免疫組化實驗表明,該蛋白廣泛分布在蟲體的實質(zhì)組織中,可能對蟲體各種細胞線粒體前體蛋白的正常代謝都會起到一定的作用。
本研究首次克隆、原核表達了SjTOM34基因,分析了該基因在日本血吸蟲不同期別和性別蟲體中的表達情況,發(fā)現(xiàn)該蛋白主要分布在蟲體的實質(zhì)組織中,并且重組的SjTOM34蛋白是具有良好免疫原性的。本文為深入研究該基因在血吸蟲生長發(fā)育中的生物學功能及其作為抗日本血吸蟲病候選疫苗分子或藥物靶標的潛力奠定了基礎。
[1]Bergquist NR.Schistosomiasis:from risk assessment to control[J].Trends Parasitol,2002,18(7):309-314.DOI:10.1016/S1471-4922(02)02301-2
[2]Lei ZL,Zheng H,Zhang LJ,et al.Schistosomiasis status in People’s Republic of China in 2010[J].Chin J Schisto Ctrl,2011,23(6):599-604.(in Chinese)
雷正龍,鄭浩,張利娟,等.2010年全國血吸蟲病疫情通報[J].中國血吸蟲病防治雜志,2011,23(6):599-604.
[3]Fallon PG,Sturrock RF,Niang AC,et al.Short report:diminished susceptibility to praziquantel in a Senegal isolate ofSchistosomamansoni[J].Am J Trop Med Hyg,1995,53(1):61-62.
[4]Liang YS,Li HJ,Dai JR,et al.Studies on resistance ofSchistosomato praziquantel XIII resistance ofSchistosomajaponicumto praziquantel is experimentally induced in laboratory[J].Chin J Schisto Ctrl,2011,23(6):605-610.(in Chinese)
梁幼生,李洪軍,戴建榮,等.血吸蟲對吡喹酮抗藥性的研究ⅩⅢ日本血吸蟲吡喹酮抗藥性的實驗誘導[J].中國血吸蟲病防治雜志,2011,23(6):605-610.
[5]Hong Y,Peng JB,Jiang WB,et al.Proteomic analysis ofSchistosomajaponicumschistosomulum proteins that are differentially expressed among hosts differing in their susceptibility to the infection[J].Mol Cell Proteomics,2011,10(8):M110 006098.DOI:10.1074/mcp.M110.006098
[6]Faou P,Hoogenraad NJ.Tom34:a cytosolic cochaperone of the Hsp90/Hsp70 protein complex involved in mitochondrial protein import[J].Biochim Biophys Acta,2012,1823(2):348-357.DOI:10.1016/j.bbamcr.2011.12.001
[7]Gebert N,Ryan MT,Pfanner N,et al.Mitochondrial protein import machineries and lipids:a functional connection[J].Biochim Biophys Acta,2011,1808(3):1002-1011.DOI:10.1016/j.bbamem.2010.08.003
[8]Chewawiwat N,Yano M,Terada K,et al.Characterization of the novel mitochondrial protein import component,Tom34,in mammalian cells[J].J Biochem,1999,125(4):721-727.
[9]Mukhopadhyay A,Avramova LV,Weiner H.Tom34 unlike Tom20 does not interact with the leader sequences of mitochondrial precursor proteins[J].Arch Biochem Biophys,2002,400(1):97-104.DOI:10.1006/abbi.2002.2777
[10]Senft AW,Gibler WB,Knopf PM.Scanning electron microscope observations on tegument maturation inSchistosomamansonigrown in permissive and non-permissive hosts[J].Am J Trop Med Hyg,1978,27(2 Pt 1):258-266.
[11]Knopf PM,Mangold BL,Makari GJ.Recovery of parasites at different stages of migration following infection of rats withSchistosomamansoni[J].Parasitology,1983,86(Pt 1):37-49.
[12]Li SK.Non-infectivity forMicrotusfortisagainstingSchistosomajaponicum[J].Parasitol Acta,1965,1:103.(in Chinese)
黎申愷.東方田鼠對日本血吸蟲的不感染性[J].寄生蟲學報,1965,1:103.
[13]Li H,He YY,Lin JJ,et al.The observation for the phenomenon ofMicrotusfortisagainstingSchistosomajaponicum[J].Chin J Vet Parasitol,2000,8(2):12-15.(in Chinese)
李浩,何艷燕,林矯矯,等.東方田鼠抗日本血吸蟲病現(xiàn)象的觀察[J].中國獸醫(yī)寄生蟲病,2000,8(2):12-15.
[14]Yang CS,Weiner H.Yeast two-h(huán)ybrid screening identifies binding partners of human Tom34 that have ATPase activity and form a complex with Tom34 in the cytosol[J].Arch Biochem Biophys,2002,400(1):105-110.DOI:10.1006/abbi.2002.2778
[15]Shimokawa T,Matsushima S,Tsunoda T,et al.Identification of TOMM34,which shows elevated expression in the majority of human colon cancers,as a novel drug target[J].Int J Oncol,2006,29(2):381-386.
