c-Jun氨基末端激酶1和2在角質形成細胞系Hacat細胞中的作用
2013-08-27 03:25:06陸承榮段連寧向培德孫麗亞
解放軍醫學院學報 2013年4期
羅 淵,陸承榮,王 喆,段連寧,向培德,孫麗亞
空軍總醫院 臨床航空醫學實驗室,北京 100142
人角質形成細胞是表皮的重要組成部分,不但可以產生角蛋白、形成角質層,構建完善的物理屏障;而且還可對部分抗原進行攝取和分解,并可產生多種細胞因子,參與皮膚免疫應答[1-2]。同時,它還是皮膚創傷修復的關鍵細胞,在促進傷口愈合、瘢痕形成以及再塑修復過程中起重要作用[3]。在炎性細胞因子和環境應激作用下,可導致JNK、ERK等通路的激活,并參與諸如皮膚老化、黑色素瘤、病毒感染等疾病的發生和發展[4-5]。絲裂原活化蛋白激酶(mitogen-activated protein kinases,MAPKs)是細胞內的一類絲氨酸/蘇氨酸蛋白激酶,是真核生物信號傳遞網絡中的重要途徑之一,在基因表達調控和細胞功能活動中發揮關鍵作用[6]。MAPK有3個主要家族:ERK、JNK和p38MAPK。ERK通路主要參與細胞的增殖與分化,JNK通路和p38MAPK通路主要參與細胞的應激反應和凋亡[7]。其中,JNK由3個同工酶組成,即組織中廣泛表達的JNK1、JNK2,以及在心臟、睪丸和腦中特異表達的JNK3。近來有研究報道JNK1和JNK2的功能存在一定的差異性,但其在角質形成細胞中是否也存在差異仍不明確[8]。本研究以人角質形成細胞系—Hacat細胞為研究對象,通過敲低其JNK1或JNK2的表達,探討JNK1和JNK2的功能差異性。
材料和方法
1 細胞系 人Hacat細胞和293T細胞為本實驗室常規保存。Hacat細胞的全培養基為含1×人類角質形成細胞生長添加劑(HKGS)的專用培養基。293T細胞的全培養基為含10%特級胎牛血清的高糖DMEM。依據細胞濃度,每2~3 d換液或消化、傳代。
2 主要試劑與儀器 人類角質形成細胞生長添加劑(S0015)、專用培養基(M-EPI-500-CA)、含EDTA的胰酶(25200-072)、高糖DMEM(11965118)和轉染試劑Lipofectamine LTX(15338-100)均購自美國Invitrogen公司。特級胎牛血清購自北京元亨圣馬公司。穿梭質粒(plko.1)和包裝質粒(PMD2G、psPAX2)均購于美國Addgene公司。JNK1/2 siRNA的oligo序列由Invitrogen公司合成。CCK-8試劑盒(CK-04)購自日本同仁化學研究所。細胞周期檢測試劑盒(340242)和細胞凋亡檢測試劑盒(559763)均購自美國BD公司。主要儀器設備包括:細胞培養箱(MCO-15,日本三洋)、倒置相差顯微鏡(CK-3,日本奧林巴斯)、酶標儀(RT-6000,深圳雷杜生命科學股份有限公司)、流式細胞儀(FACS Calibur,美國BD公司)。
3 慢病毒法建立JNK1、 JNK2敲低細胞株及鑒定根據在線網站工具(http://jura.wi.mit.edu/siRNAext)設計針對JNK1和JNK2的siRNA靶序列,將靶序列代入穿梭質粒的框架后送公司合成。分別將上下游oligo溶解后退火,與經EcoRI和AgeI雙酶切后回收的plko.1載體連接,并轉化感受態細胞,挑選成功的重組載體。將293T細胞鋪板,密度為7×105細胞/60 mm平皿,加入2 μg穿梭質粒和包裝質粒各1 μg進行共轉染,第2天換新鮮培養基,24 h后收獲病毒液,經0.44 μm過濾后分裝、凍存于-80 ℃。培養Hacat細胞,約5×105細胞/60 mm平皿,加入1 ml病毒液和3 ml全培養基,添加polybrene(8 μg/μl),5 h后換新鮮培養基,48 h后加入嘌呤霉素(2 μg/μl)開始篩選。適當收獲一些細胞,裂解后進行JNK1、 JNK2的Western Blot鑒定。
4 CCK-8法檢測細胞增殖曲線 在96孔板中分別鋪Hacat細胞、JNK1和JNK2敲低細胞,每孔鋪104個細胞,檢測時間點設為0 h、18 h、36 h和54 h,每個時間點設3個復孔和3個空白對照孔(不加CCK-8)。到相應時間點時,向每個實驗孔中加入10 μl CCK-8,振蕩混勻后放回37 ℃孵箱再培養4 h,然后進行雙波長酶標儀檢測,主波長為450~490 nm,次波長為600~650 nm,以實驗孔OD值減去相應的空白對照孔OD值為最終結果。
5 流式細胞術檢測細胞周期 在6孔板中分別鋪Hacat細胞、JNK1和JNK2敲低細胞,每孔鋪4×105個細胞,每種細胞各做3個復孔。細胞貼壁后開始血清饑餓24 h,即換成只含0.5%血清的DMEM培養基。第2天,重新換成專用全培養基,啟動細胞增殖。24 h后胰酶消化細胞,收集后按BD細胞周期試劑盒進行染色,流式細胞儀檢測。
6 流式細胞術檢測細胞凋亡 在6孔板中分別鋪Hacat細胞、JNK1和JNK2敲低細胞,每孔鋪4×105個細胞,細胞貼壁后分別加入不同濃度的Taxol(0 nmol/L、100 nmol/L、200 nmol/L 和 400 nmol/L),每個劑量點做3個復孔。藥物作用48 h后,收集細胞,按照BD凋亡檢測試劑盒說明書進行檢測。7 統計學分析 采用SPSS17.0軟件包進行統計學分析。單因素方差分析檢驗Hacat細胞、JNK1和JNK2敲低細胞的組間差異,q檢驗進行兩兩比較。
結 果
1 JNK1、JNK2敲低表達Hacat穩轉株的鑒定 分別構建JNK1、JNK2敲低的細胞株,JNK1(46 kU)敲低約50%,而JNK2(54 kU)幾乎完全敲除,結果見圖1,siRNA序列見表1。
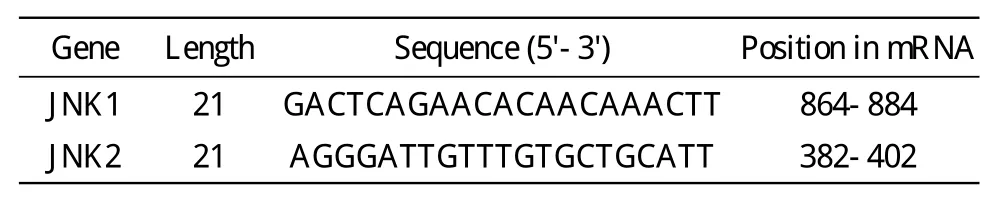
表1 有效的針對JNK1/2的siRNA序列Tab.1 Sequences of JNK1/2 siRNA
2 JNK1/2敲低對Hacat細胞增殖的影響 在細胞培養早期(18 h),三種細胞的增殖變化不明顯(P=0.626>0.05)。培養中期(36 h),三種細胞的增殖差異有統計學意義(P=0.000<0.05)。經兩兩比較,對照組高于JNK1敲低組(P=0.000<0.05)和JNK2敲低組(P=0.017<0.05),JNK2敲低組也高于JNK1敲低組(P=0.002<0.05)。培養后期(54 h),因未進行細胞換液,部分細胞死亡,故OD值下降,見圖2。
3 JNK1/2敲低對Hacat細胞周期的影響 三組細胞在G0/G1期的分布無統計學差異(P=0.245>0.05),而S期和G2/M期均有差異(P=0.000<0.05)。兩兩比較發現,S期的JNK1敲低組低于對照組和JNK2敲低組(P=0.000<0.05),對照組和JNK2敲低組間無統計學差異(P=0.065>0.05)。G2/M期的JNK1敲低組高于對照組和JNK2敲低組(P=0.000<0.05),對照組和JNK2敲低組間無統計學差異(P=0.088>0.05)。可見,JNK1敲低使增殖期細胞顯著減少,細胞阻滯于G2/M期,而JNK2敲低對細胞周期影響不大,見圖3。
4 JNK1/2敲低對Hacat細胞凋亡的影響 在不同劑量點,組間比較均有統計學差異(P<0.05)。兩兩比較發現,JNK2敲低組的凋亡率在每個劑量點都高于對照組和JNK1敲低組(P<0.05),而JNK1敲低組和對照組在100 nmol/L和400 nmol/L時差異不顯著,而在200 nmol/L時對照組凋亡率增高。表明JNK2抗凋亡作用高于JNK1,而JNK1可能有促凋亡作用,見圖4。

圖1 JNK1/2敲低效果的Western Blot鑒定(JNK1-46 kU、JNK2-54 kU)Fig.1 Western blot showing the knocked-down JNK1-46 kU and JNK2-54 kU
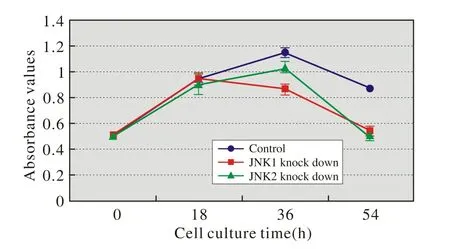
圖2 JNK1/2敲低對Hacat細胞增殖的影響(CCK-8法)Fig.2 CCK-8 assay showing effect of knocked-down JNK1/2 on proliferation of Hacat cells
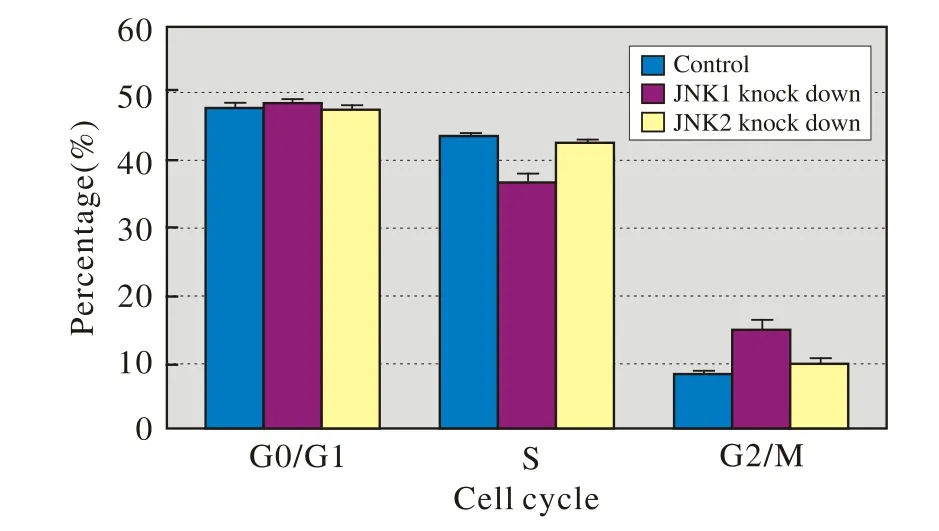
圖3 JNK1/2敲低對Hacat細胞周期的影響(流式細胞術法)Fig.3 Flow cytometry showing effect of knocked down JNK1/2 on cell cycle of Hacat cells
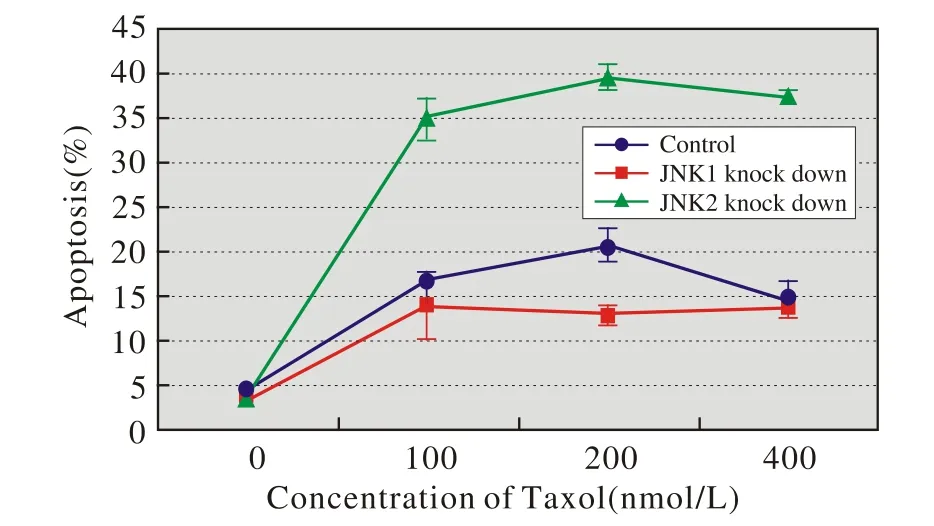
圖4 JNK1/2敲低對Hacat細胞凋亡的影響(流式細胞術法)Fig.4 Flow cytometry showing effect of knocked-down JNK1/2 on apoptosis of Hacat cells
討 論
本研究利用慢病毒載體系統,成功構建了JNK1或JNK2敲低表達的Hacat穩定轉染細胞株,先后進行了細胞增殖、細胞周期和Taxol的凋亡誘導實驗,通過和正常Hacat細胞對比,發現培養36 h后,JNK1/2敲低時細胞增殖顯著減緩,尤以JNK1敲低更顯著。細胞周期結果表明,啟動培養24 h后,三組細胞在G0/G1期的分布無統計學差異,而S期和G2/M期均有統計學差異。其中,在S期JNK1敲低組明顯低于對照組和JNK2敲低組,而JNK1敲低組的G2/M期明顯高于其余兩組,可見JNK1敲低使增殖期細胞顯著減少,細胞阻滯于G2/M期,而JNK2敲低對細胞周期影響不大。凋亡誘導實驗表明,JNK2敲低組的凋亡率在每個劑量點都顯著高于對照組和JNK1敲低組,可見JNK2抗凋亡作用顯著高于JNK1,而JNK1甚至可能有促凋亡作用。初步表明,JNK1和JNK2的作用存在不同。在細胞增殖方面,JNK1作用強于JNK2;而在抗凋亡方面,JNK2作用強于JNK1。
目前,關于JNK的研究已較為深入,已發現MEK4和MEK7兩種直接上游激酶以及若干種下游 分 子, 如 c-Jun、ATF2、P53、Elk-1、c-Myc2、Bcl-2、Bim、BAD等[9]。Ke等學者的研究表明,在70%的人表皮細胞瘤中JNK2高度活化,而且通過藥物或基因方法抑制JNK2的活性可抑制腫瘤發生,可見JNK2的抗凋亡作用較大[8]。Liu等學者通過基因敲除小鼠也發現,TNF-α引起的凋亡在JNK1缺乏的成纖維細胞中受到抑制,而JNK2缺乏的反而增強,這與我們的研究結果一致[10]。而Zhong等學者研究發現,JNK1可磷酸化Elk-1而導致TBP表達增高,并最終促進增殖,而JNK2起抑制作用。相應地,JNK1缺失時,細胞增殖變緩,而JNK2缺失時,細胞增殖加快[11]。這與我們的結果部分一致,可能是由不同細胞或不同刺激類型所致。
JNK在細胞增殖、分化、凋亡中的作用機制錯綜復雜。不同的組織細胞來源、不同的刺激因素或不同的JNK同工酶,可激活不同的信號轉導通路[12]。大量的研究表明,JNK信號通路密切參與許多疾病的發生和發展,如腫瘤、神經系統疾病、免疫性疾病等,而且可能是某些疾病的治療靶點[13-15]。本研究發現JNK1和JNK2在人類皮膚角質形成細胞中存在不同的功能,尤其是抑制JNK2可以大大促進細胞凋亡,為皮膚疾病的相關研究打下了基礎。但也存在一些不足,如本研究未對原代細胞進行相應的驗證,而且沒有進行JNK1/2過度表達的研究,這也是我們后續的研究思路。
1 Morita K, Miyachi Y, Furuse M. Tight junctions in epidermis: from barrier to keratinization[J]. Eur J Dermatol, 2011, 21(1): 12-17.
2 Boulais N, Misery L. The epidermis: a sensory tissue[J]. Eur J Dermatol, 2008, 18(2):119-127.
3 Suter MM, Schulze K, Bergman W, et al. The keratinocyte in epidermal renewal and defence[J]. Vet Dermatol, 2009, 20(5-6):515-532.
4 Sabapathy K. Role of the JNK pathway in human diseases[J]. Prog Mol Biol Transl Sci, 2012, 106 :145-169.
5 孫琳,劉湘源,趙金霞,等.Raf/MEK/ERK信號通路對CXCL16誘導的CIA淋巴細胞增殖的影響[J].軍醫進修學院學報,2010,31(11):1113-1115,1128.
6 Huang P, Han J, Hui L. MAPK signaling in inflammation-associated Cancer development[J]. Protein Cell, 2010, 1(3): 218-226.
7 Plotnikov A, Zehorai E, Procaccia S, et al. The MAPK cascades: signaling components, nuclear roles and mechanisms of nuclear translocation[J].Biochim Biophys Acta, 2011, 1813(9): 1619-1633.
8 Ke H, Harris R, Coloff JL, et al. The c-Jun NH2-terminal kinase 2 plays a dominant role in human epidermal neoplasia[J]. Cancer Res, 2010, 70(8):3080-3088.
9 Dhanasekaran DN, Reddy EP. JNK signaling in apoptosis[J].Oncogene, 2008, 27(48): 6245-6251.
10 Liu J, Minemoto Y, Lin A. c-Jun N-terminal protein kinase 1(JNK1), but not JNK2, is essential for tumor necrosis factor alphainduced c-Jun kinase activation and apoptosis[J]. Mol Cell Biol,2004, 24(24):10844-10856.
11 Zhong S, Fromm J, Johnson DL. TBP is differentially regulated by c-Jun N-terminal kinase 1 (JNK1) and JNK2 through Elk-1,controlling c-Jun expression and cell proliferation[J]. Mol Cell Biol, 2007, 27(1): 54-64.
12 Cargnello M, Roux PP. Activation and function of the MAPKs and their substrates, the MAPK-activated protein kinases[J]. Microbiol Mol Biol Rev, 2011, 75(1): 50-83.
13 Borsello T, Forloni G. JNK signalling : a possible target to prevent neurodegeneration[J]. Curr Pharm Des, 2007, 13(18): 1875-1886.
14 Cui J, Zhang M, Zhang YQ, et al. JNK pathway : diseases and therapeutic potential[J]. Acta Pharmacol Sin, 2007, 28(5):601-608.
15 Heasley LE, Han SY. JNK regulation of oncogenesis[J]. Mol Cells, 2006, 21(2):167-173.
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
海峽科技與產業(2016年3期)2016-05-17 04:32:12
