二斑葉螨侵染下菜豆葉片交替呼吸途徑對光合作用、葉溫和間接防御的影響
2013-09-28 01:44:26馮漢青張晉平賈凌云
植物保護 2013年1期
馮漢青, 魏 陽, 張晉平, 孫 坤, 陳 紋, 賈凌云
(西北師范大學生命科學學院,蘭州 730070)
在自然環境中,植物頻繁地遭到害蟲的侵染。近年來,害蟲侵染下植物代謝及防御反應的研究已經成為植物學研究的重點和熱點問題之一。然而,盡管線粒體呼吸是植物物質和能量代謝的中心環節[1],其在蟲害侵染下的變化以及其可能的生理學功能到目前為止尚不清楚,這制約了人們對蟲害侵染下植物生理學反應更加全面的認識。
交替呼吸途徑是高等植物呼吸系統區別于動物呼吸系統的重要特征之一。這條呼吸途徑的運行可以將電子繞過細胞色素氧化酶而直接傳遞到交替氧化酶,并將分子氧還原成水。由于交替呼吸途徑的運行使得電子傳遞越過了形成質子梯度的兩個位點,故能將形成腺嘌呤核苷三磷酸(ATP)的部分化學能以熱的形式放出[2]。研究發現,交替呼吸途徑能夠被病原菌侵染、低溫、干旱等多種環境壓力因素所誘導,并在植物抵抗上述環境壓力中扮演著重要的角色[3]。但到目前為止,是否交替呼吸途徑也和植物的抗蟲反應相關國內外尚無報道。
研究發現,在害蟲侵染下,植物能夠通過氣味來吸引害蟲的天敵,從而間接減少害蟲在植物上的數量,這種防御機制被命名為間接防御反應[4]。而早期的研究發現,交替呼吸路徑所介導的植物產熱作用能夠提升花器的氣味釋放繼而吸引昆蟲為之授粉[5]。盡管目前尚不清楚植物通過氣味吸引害蟲天敵的作用是否和交替呼吸路徑所介導的產熱功能有關,但植物吸引授粉昆蟲和害蟲天敵行為上的相似性提示了交替呼吸路徑有可能和間接防御反應之間存在著一定的聯系。此外,光合作用通常會因害蟲的侵染而降低[6];但光合作用被發現為植物吸引害蟲天敵氣味的合成提供了底物[7];故在害蟲侵染下,光合作用的維持和保護可能對于植物的間接防御具有重要的作用。而目前的研究發現,在干旱、高光等環境因素造成光合作用抑制時,交替呼吸路徑能保護光合作用[8]。因此,交替呼吸路徑也可能通過光合作用和間接防御反應之間存在著聯系。基于以上分析,交替呼吸路徑的生理學特性迎合了植物間接防御反應的生理代謝需求,可能是植物呼吸代謝和植物抗蟲策略之間關系研究的重要切入點之一。基于此,我們在由菜豆(Phaseolus vulgaris Linn.)葉片、害蟲二斑葉螨(Tetranychus urticae Koch)以及二斑葉螨的捕食天敵智利小植綏螨(Phytoseiulus persimilis Athias-Henriot)組成的三級互作體系中初步分析了交替呼吸途徑和植物間接防御反應的關系及其可能的內在機理。相信該研究有利于進一步了解蟲害下線粒體呼吸代謝的變化情況及其可能扮演的生理學角色。
1 材料與方法
1.1 植物材料的培養
菜豆種子經次氯酸消毒和萌發后種植于營養土中,在培養室中生長,生長溫度為25℃,光強為100μmol photons/m2·s光照周期為 L∥D=16h∥8h。部分植物材料用于害蟲及其天敵的飼養;另選取2~3周齡的菜豆幼苗作為供試材料進行處理和生理指標的測量(見1.2~1.7)。
1.2 植物材料的害蟲侵染
挑選30頭成年雌性二斑葉螨,用毛刷轉移到供試菜豆植株第一葉上,使其均勻分布;以未經害蟲侵染處理植株的第一葉作為對照。之后將植物置于上述生長條件下繼續培養。
1.3 呼吸抑制劑處理
在害蟲侵染第10天 的16h光照結束后,將10個連有葉柄的葉片切下,放置于內部盛有一定量蒸餾水的2L玻璃瓶中(葉柄朝下浸沒在蒸餾水中以避免葉片水分喪失),先于黑暗下適應4h。之后,依據Martim等描述的方法,將蒸餾水替換為1mmol/L的水楊基氧肟酸 (SHAM,salicylhydroxamic acid;該抑制劑濃度已被報道不會對光合作用等其他代謝造成影響[9]),并在黑暗下維持4h,使SHAM 通過葉柄輸入到葉片組織中以抑制交替呼吸途徑[10];以未經SHAM溶液處理的葉片作為對照。將SHAM處理的葉片和對照葉片經歷3d的光周期(瓶口打開,瓶內光強保持在5 000lx,光周期L∥D=16h∥8h),每天光照結束后替換一次SHAM以保持對葉片交替呼吸途徑的抑制。于第4天進行指標測定。
1.4 不同溫度和光照的處理
在害蟲侵染第10天的16h光照結束后,依據上述辦法(見1.3)將10個連有葉柄的侵染葉片放置于盛有蒸餾水的2L玻璃瓶中,先于黑暗下適應8h,之后進行不同溫度和光照處理。不同溫度處理:將盛有葉片的玻璃瓶在兩種不同溫度下(20℃和25℃)維持3d(瓶口打開,瓶內光強保持在5 000lx,光周期L∥D=16h∥8h),于第4天利用Y型嗅覺儀觀察不同溫度生長下植物對害蟲天敵吸引力的變化。不同光照的處理:將盛有葉片的玻璃瓶放置于兩種不同光照條件下(瓶內光強分別為2 800lx和5 000lx)維持3d(瓶口打開,溫度為25℃,光周期L∥D=16h∥8h),于第4天利用Y型嗅覺儀觀察不同光強生長下的植物對害蟲天敵吸引力的變化。
1.5 利用Y型嗅覺儀觀察植物對害蟲天敵的吸引作用
Y型嗅覺儀由一條玻璃中管和兩條等長的玻璃臂管組成(臂管長10cm;中管長12cm;內徑為4cm)。兩個臂管分別連接兩個盛有葉片的2L玻璃瓶,每個玻璃瓶中放置10個葉片作為氣味源(見1.3和1.4)。密封瓶口,使經活性炭凈化的空氣(流速為3L/min)經過氣味源后流向Y型嗅覺儀的臂管。將捕食螨智利小植綏螨引入Y型嗅覺儀中管下風口處,觀察捕食螨對兩個臂管的選擇行為。如果捕食螨在5min內進入了某一臂管,則認為其做了趨向性選擇;如果在5min內捕食螨未能進入任何一個臂管,則認為其沒有做出趨向性選擇。整個觀察時間為3h,并對結果進行卡方(χ2)檢測。
1.6 呼吸的測量
在呼吸測量之前,將葉片上二斑葉螨移除以避免其對葉片呼吸的干擾。之后將葉片稱重并剪成小塊,利用Clark氧電極依據以前的方法測量葉片的總呼吸和交替呼吸的容量[11]。
1.7 光合速率和葉片溫度的測定
光合速率利用便攜式開放氣路型光合儀測量(PP Systems,Hertfordshire,UK),葉室的氣體流速為300mL/min,光合速率和葉片溫度的測量依據操作手冊進行。
2 結果
2.1 二斑葉螨侵染對菜豆葉片總呼吸和交替呼吸途徑水平的影響
二斑葉螨的侵染顯著地增加了菜豆葉片總呼吸和交替呼吸途徑容量的水平(表1)。在未遭害蟲侵染的對照葉中,交替呼吸途徑容量在總呼吸中的比值為30.3%,害蟲侵染葉中這一比值上升至為36.8%,表明害蟲侵染提升了交替呼吸途徑在總呼吸中的比例。

表1 二斑葉螨侵染對菜豆葉片總呼吸和交替呼吸途徑容量水平的影響1)Table 1 Effects of T.urticaeinfestation on total respiration(Vt)and alternative pathway capacity(Valt)
2.2 交替呼吸途徑影響了害蟲侵染葉片對天敵的吸引作用
二斑葉螨侵染的葉片較之未遭侵染的葉片能夠吸引更多的天敵智利小植綏螨(圖1)。而較之未經SHAM處理的二斑葉螨侵染葉片,SHAM處理后的害蟲侵染葉片對智利小植綏螨的吸引作用有所降低(圖1),表明了交替呼吸途徑的抑制降低了害蟲侵染葉片對捕食螨的吸引能力。
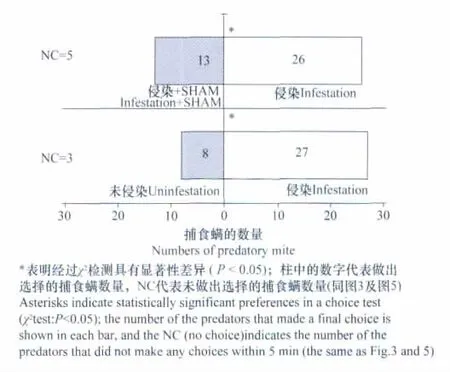
圖1 智利小植綏螨在Y型嗅覺儀中對二斑葉螨侵染葉片、未侵染葉片及水楊基氧肟酸(SHAM)處理的二斑葉螨侵染葉片選擇趨向性的比較Fig.1 Responses of P.persimilisin a Y-tube olfactometer when offered the spider mite-infested leaves vs.the uninfested leaves or the infested leaves treated with SHAM
2.3 交替呼吸途徑的抑制對害蟲侵染葉片溫度的影響
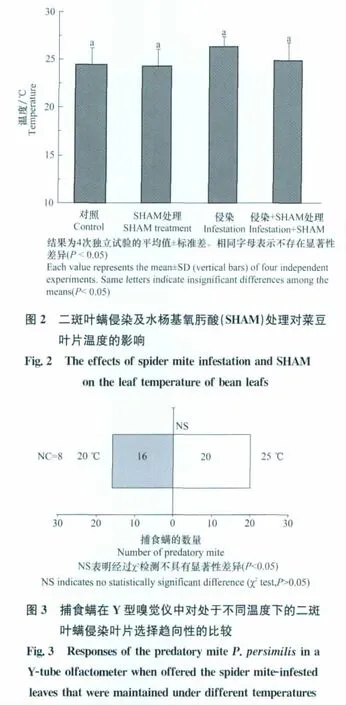
在未遭二斑葉螨侵染的葉片中,SHAM處理沒有對葉片的葉溫產生影響;二斑葉螨侵染葉片的葉溫比對照葉的葉溫略高,但并沒有顯著差異(圖2)。較之未經SHAM處理的二斑葉螨侵染葉,SHAM處理后的害蟲侵染葉片的葉溫平均降低了1.5℃,但兩者之間也無顯著性差異(圖2)。
試驗發現,SHAM處理二斑葉螨侵染葉片所引起的最大葉溫的降低是5℃,進一步的試驗比較了這個水平上的溫度降低是否會對蟲害侵染葉片吸引害蟲天敵的作用產生影響。結果顯示,處于20℃下的害蟲侵染葉片和25℃下的害蟲侵染葉片吸引智利小植綏螨的數量之間沒有顯著性差異(圖3)。上述的觀察(圖2和3)表明,SHAM處理所造成的害蟲侵染葉片對捕食螨吸引作用的降低可能與溫度的變化無關。
2.4 交替呼吸途徑的抑制對害蟲侵染葉片光合速率的影響
在未遭二斑葉螨侵染的葉片中,SHAM處理沒有對葉片的光合速率產生顯著性影響;二斑葉螨侵染葉較之對照葉片有著較低的光合速率;而在二斑葉螨侵染葉中,SHAM處理進一步顯著降低了葉片的光合速率(圖4)。
較低的光強度(大約為2 800lx)照射二斑葉螨侵染葉片使其光合速率低到和SHAM處理后的二斑葉螨侵染葉片光合速率相似的水平,影響到害蟲侵染葉片對捕食螨的吸引作用(圖5);結合上述的觀察(圖4和5),SHAM處理所造成的害蟲侵染葉片對捕食螨吸引作用的降低可能和其引起的光合速率的下降有關。

3 討論
受二斑葉螨侵染的葉片較之未遭侵染的葉片能夠顯著地吸引更多的捕食螨(圖1),且有著較高水平的交替呼吸途徑(表1)。而用SHAM處理明顯降低了害蟲侵染葉片對捕食螨的吸引作用(圖1),表明了害蟲侵染葉片對捕食螨的吸引作用可能和交替呼吸途徑的上升有關。
如前文所述,產熱植物中交替呼吸途徑水平的升高能夠促進植物產熱,從而提升了花器氣味的釋放以吸引昆蟲為之授粉。其后,在一些非產熱植物的組織中(如煙草和高粱的葉片)也發現了交替途徑水平上升促進組織熱量生成的現象[12-13]。因此,推測交替呼吸途徑抑制所引起的害蟲侵染葉片對捕食螨吸引作用下降的現象(圖1)可能和交替呼吸途徑的產熱功能有關。但試驗結果顯示,害蟲侵染并未引起葉片溫度的顯著性升高;且SHAM的處理也沒有引起害蟲侵染葉片葉溫的顯著下降(圖2)。在試驗中觀察到,SHAM處理害蟲侵染葉片所引起的最大水平的溫度降低是5°C;但此溫度不會影響害蟲侵染葉片對捕食螨的吸引作用(圖3)。綜上結果表明,交替呼吸途徑抑制所引起的害蟲侵染葉片對捕食螨吸引作用下降的現象可能和溫度的變化無關。有研究表明,線粒體的豐度、呼吸底物的供應水平以及交替氧化酶碳端等影響線粒體將化學能轉化為熱量的因素在產熱植物中和非產熱植物中均有著明顯不同,這可能制約了非產熱植物利用交替呼吸途徑去產生熱量的能力[14]。而本研究的觀察也反映了盡管交替途徑的抑制能輕微地降低非產熱植物葉片的溫度,但這種影響是非常有限且不顯著。據此,本研究認為非產熱植物中可能并不存在通過交替呼吸途徑的產熱來吸引害蟲天敵的生理機制。
盡管交替呼吸途徑看上去并非通過葉片溫度的變化來影響害蟲侵染葉片對捕食螨的吸引作用,但本研究發現,SHAM的處理導致了害蟲侵染葉片光合速率的顯著下降(圖4),表明了害蟲侵染下交替呼吸途徑能夠保護植物的光合作用。許多研究也發現,在干旱和高光等逆境下,交替呼吸途徑的抑制也會導致植物葉片光合作用的下降[8]。有學者提出了逆境下交替呼吸途徑對植物光合作用影響的內在機理,認為逆境脅迫會導致葉綠體中還原力的過度產生,如果這些過多的還原力沒有被消散就會導致葉綠體中活性氧水平的上升,從而對葉綠體造成氧化損傷并繼而降低光合作用;而植物可以通過蘋果酸-草酰乙酸穿梭等路徑將葉綠體中過多的還原力傳遞到線粒體,并被交替呼吸途徑所消散,從而保護了光合作用[8]。已有報道表明,害蟲侵染也會導致葉片中還原力的過度積累和氧化損傷[15-16]。據此,本研究認為,在害蟲侵染下交替呼吸途徑可能通過上述機制保護了葉片的光合作用。而如前文所述,植物吸引害蟲天敵的氣味的體內合成所需的底物主要通過光合作用產生。當用一個較低的光強度照射二斑葉螨侵染的葉片使其光合速率降低到了和SHAM處理后的二斑葉螨侵染葉片光合速率相似的水平時,害蟲侵染葉片對捕食螨的吸引作用也有所降低(圖5),表明了害蟲侵染葉片對捕食螨的吸引作用可能和其光合速率水平有關。由此猜測,交替呼吸途徑抑制劑處理下害蟲侵染葉片對捕食螨吸引作用降低的現象可能其是導致光合速率降低的結果。
必須提出的是,未經二斑葉螨侵染的葉片較之侵染的葉片有著更高的光合速率,但卻無法吸引更多的捕食螨(圖1和4)。這似乎與葉片對捕食螨的吸引作用可能和光合速率有關這一結論有矛盾之處。但有研究提出,害蟲侵染是引起葉片對捕食螨產生吸引作用的前提條件:害蟲侵染引發了植物體內信號路徑的改變,從而激活了一些特異性基因的表達,這些基因的表達產物利用了光合作用所提供的底物以合成吸引害蟲天敵的氣味[17]。
交替呼吸途徑也可能以其他方式去影響植物的間接防御反應,但本文的試驗結果表明了光合作用很可能是聯系交替呼吸途徑和植物間接防御反應的重要節點。
[1] Plaxton W,Podesta F.The functional organization and control of plant respiration[J].CRC Critical Reviews Plant Sciences,2006,25:159-198.
[2] Millenaar F F,Lambers H.The Alternative oxidase:in vivo regulation and function[J].Plant Biology,2003,5:2-15.
[3] Van Aken O,Giraud E,Clifton R,et al.Alternative oxidase:a target and regulator of stress responses[J].Physiologia Plantarum,2009,137:354-361.
[3] Degenhardt J.Indirect defense responses to herbivory in grasses[J].Plant Physiology,2009,149:96-102.
[5] Meeuse B D J.Thermogenic respiration in Aroids[J].Annual Review of Plant Biology,1975,26:117-126.
[6] Kerchev P I,Fenton B,Foyer C H,et al.Plant responses to insect herbivory:interactions between photosynthesis,reactive oxygen species and hormonal signalling pathways[J].Plant Cell Environment,2012,35(2):441-53.
[7] Staudt M,Lhoutellier L.Monoterpene and sesquiterpene emissions fromQuercus coccifera exhibit interacting responses to light and temperature[J].Biogeosciences,2011,8:2757-2771.
[8] Arnholdt-Schmitt B,Costa J H,deMelo D F.AOX-a functional marker for efficient cell reprogramming under stress[J].Trends Plant Science,2006,11:281-287.
[9] Bartoli C G,Gómez F,Gergoff G,et al.Up-regulation of the mitochondrial alternative oxidase pathway enhances photosynthetic electron transport under drought conditions[J].Journal of Experimental Botany,2005,56(415):1269-1276.
[10] Martim S A,Santos M P,Pe?anha A L,et al.Photosynthesis and cell respiration modulated by water deficit in grapevine(Vitis vinifera L.)cv[J].Cabernet Sauvignon Brazilian Journal of Plant Physiology,2009,21:95-102.
[11] Feng H Q,Wang Y Y,Li H Y,et al.Salt stress-induced expression of rice AOX1ais mediated through an accumulation of hydrogen peroxide[J].Biologia,2010,65:868-873.
[12] Van Der Straeten D,Chaerle L,Sharkov G,et al.Salicylic acid enhances the activity of the alternative pathway of respiration in tobacco leaves and induces thermogenicity[J].Planta,1995,196:412-419.
[13] Kumar S,Sinha S K.Possible role of alternative respiration in temperature rise of water stressed plants[J].Journal of Biosci-ences,1994,19(3):331-338.
[14] Grant N,Onda Y,Kakizaki Y,et al.Two Cys or not two Cys?That is the question;alternative oxidase in the thermogenic plant sacred lotus[J].Plant Physiology,2009,150:987-995.
[15] Heng-Moss T M,Ni X,Macedo T,et al.Comparison of chlorophyll and carotenoid concentrations among Russian wheat aphid (Homoptera:Aphididae)-infested wheat isolines[J].Journal of Economic Entomology,2003,96(2):475-81.
[16] Botha A M,Lacock L,van Niekerk C,et al.Is photosynthetic transcriptional regulation in Triticum aestivum L. cv.′TugelaDN′a contributing factor for tolerance to Diuraphis noxia (Homoptera:Aphididae)[J].Plant Cell Reports,2006,25(1):41-45.
[17] Arimura G,K?pke S,Kunert M,et al.Effects of feeding Spodoptera littoralis on lima bean leaves:IV.diurnal and nocturnal damage differentially initiate plant volatile emission[J].Plant Physiology,2008,146(1):965-973.
猜你喜歡
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
中國衛生(2016年3期)2016-11-12 13:23:26
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
