G蛋白偶聯受體激酶在心血管疾病中的作用*
2013-12-01 02:28:58田艷君劉海青
中國病理生理雜志 2013年7期
田艷君, 劉海青, 陳 京, 白 波
(1曲阜師范大學,山東曲阜273165;2泰山醫學院,山東 泰安271000;3濟寧醫學院,山東濟寧272000)
G蛋白偶聯受體 (G-protein-coupled receptors,GPCRs)是與G蛋白偶聯的膜受體,也是目前已知最大的細胞膜受體家族。GPCRs能夠被不同配體所激活,可將激素、神經遞質、藥物、趨化因子以及光線等多種化學性和物理性細胞外信號轉導入細胞內,在多種生理活動和病理過程中發揮重要的調節作用。GPCRs被認為是高血壓、心力衰竭、帕金森氏綜合癥等多種疾病的藥物治療靶點,市售藥物約50%都是以 GPCRs為靶點的[1]。
G蛋白偶聯受體激酶(G-protein-coupled receptor kinases,GRKs)是一種絲氨酸/蘇氨酸激酶,主要由7個成員組成,按發現時間順序分別命名為GRK1~GRK7,它們擁有共同的功能結構(圖1),包括1個中心催化區、1個供底物識別和含有G蛋白信號調節蛋白(regulators of G-protein signaling,RGS)樣結構的氨基末端以及1個作用于細胞膜的長度可變的羧基末端[2-3]。RGS區具有三磷酸鳥苷(guanosine triphosphate,GTP)酶活性,能促進GTP(束縛于Gi和Gq的α亞基)的水解,從而緩沖經由Gαi和Gαq通路的信號轉導[4]。
GRKs是GPCRs信號轉導的關鍵負調節因子,GPCRs與配體結合后,除了能促進G蛋白的激活,還可以引發GRKs參與的GPCRs的磷酸化。GPCRs一旦發生磷酸化,抑制蛋白(β-arrestins)便會迅速與之結合,這將大大削弱受體與G蛋白對刺激物的應答作用。這種受體應答作用迅速喪失的現象稱為脫敏(圖2)。近年來的研究表明,GRKs的作用不止是使GPCR脫敏,它們還可以磷酸化非受體底物,如微管蛋白、突觸核蛋白、光傳感因子等[5]。此外,GRKs還可以與信號轉導和運輸中的多種蛋白[如Gαq、Gβγ、G蛋白偶聯受體激酶相互作用蛋白(GRK-interacting proteins,GIT)、磷脂酰肌醇3-激酶(phosphatidylinositol 3-ki-nase,PI3K)、網格蛋白和小窩蛋白等]發生相互作用, 以一種磷酸化非依賴的方式調節細胞功能。
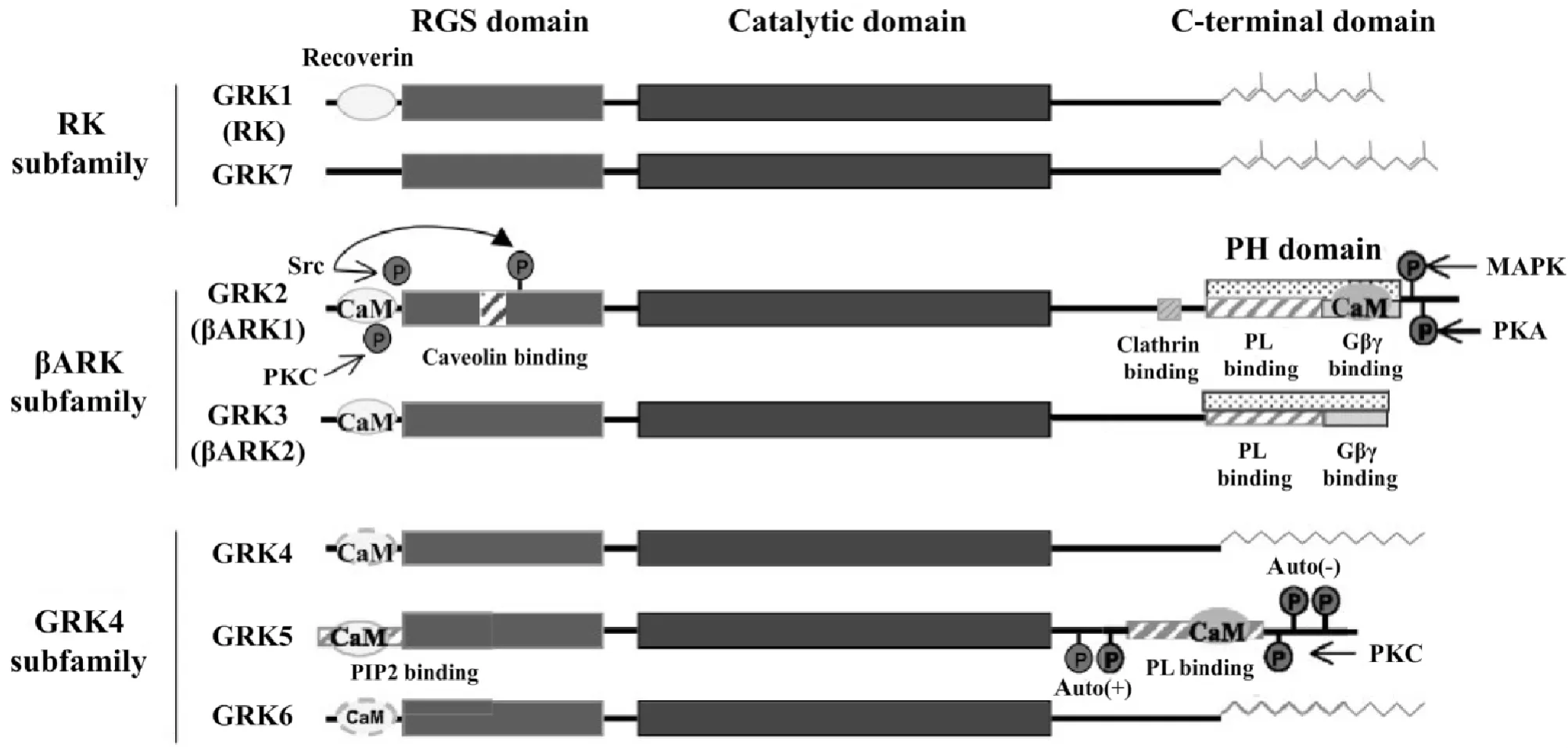
Figure 1.The construction features and classification of GRKs[2].GRKs are multidomain proteins interacting with a variety of cellular proteins.Such interactions determine the different cellular functions of these kinases and also contribute to modulating their activity and subcellular targeting.The figure depicts the regions where the different functional domains and regulatory phosphorylation and interaction sites have been mapped.RGS:regulators of G-protein signalling;CaM:calmodulin;PL:phospholipids;PH:pleckstrin homology;Auto(±):stimulatory or inhibitory autophosphorylation sites.圖1 GRKs的結構特點和分類
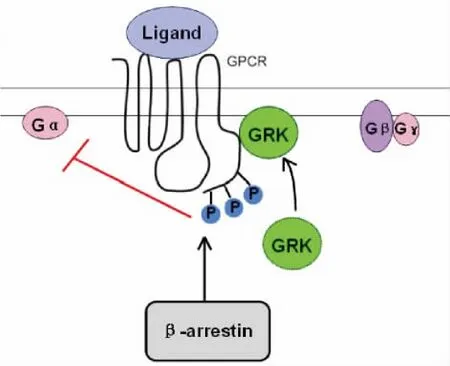
Figure 2.Desensitization of GPCRs.GRKs are recruited to,and phosphorylate ligand-occupied GPCRs on the cytoplasmic carboxyl-terminal tail.β-arrestins then recruit to phosphorylated GPCRs with enhanced affinity,thereby creating a platform for blocking recoupling of the dissociated G-protein subunits to the GPCRs,and preventing further receptor activation(ie,desensitization).GRK:G-protein-coupled receptor kinase;GPCR:G-protein-coupled receptor.圖2 GPCRs的脫敏過程
1 GRKs的分類和分布
已知哺乳動物體內的7種GRKs中,GRK1和GRK7分別分布于視網膜的視桿細胞和視錐細胞,介導光信號轉導;GRK4僅在小腦、睪丸和腎臟表達;而GRK2、GRK3、GRK5和 GRK6在多數組織中均有表達[6]。
基于序列同源性的高低和組織表達特異性不同,GRKs可以分為3個亞家族:視紫紅質激酶(rhodopsin kinase,RK)亞家族(GRK1 和 GRK7)、β-腎上腺素受體激酶(β-adrenergic receptor kinase,βARK)亞家族(GRK2和 GRK3)和 GRK4亞家族(GRK4、GRK5和 GRK6),見圖1。
2 GRKs在心臟的表達和分布
研究顯示,GRK2、GRK3和GRK5在健康人的心臟中均有表達,然而GRK4、GRK6和GRK7幾乎檢測不到[7]。其中,由于 C端功能區的差異,GRK2和GRK3位于胞漿內,而GRK5位于細胞膜。這3種GRKs在心臟的表達雖然存在某些共性,但它們是有區別的、具有調節特性的酶。體內實驗顯示,GRK3對凝血酶受體和α1B腎上腺素受體(α1B-adrenergic receptor,α1BAR)有特異性,GRK5對血管緊張素Ⅱ受體有選擇性,而GRK2和GRK5在心肌的β-腎上腺素能受體(β-adrenergic receptor,βAR)βAR 系統中發揮重要作用[7-8]。而且,這些激酶及其受體的細胞分布和亞細胞定位也為GPCR-GRK相互作用的特異性提供了有利條件。
在胚胎形成和心臟發育過程中,不同的GRKs作用于不同的信號轉導通路。敲除GRK2基因小鼠由于嚴重的心臟發育不全和心力衰竭,一般在胎齡12.5~15.5 d死亡;然而,缺乏GRK3或GRK5的小鼠則能成活,不會出現上述情況[7,9]。這種由于GRK2基因敲除而引起的胚胎致死和心血管損傷表明,GRK2在胚胎形成和心臟發育中擁有其它GRKs亞型所沒有的特殊的關鍵功能。
雖然以上3種GRKs在心臟均有分布,但是它們在心臟的各種細胞類型中的分布卻有差異。大鼠心臟的免疫組織化學分析顯示,GRK2在心肌細胞中僅有微弱的表達,主要分布于血管內皮細胞;GRK3僅局限于心肌細胞;而GRK5則在心臟的各種類型細胞中均有分布[3]。心肌組織中不同GRKs亞型的分布反映了它們生理學作用底物的偏向性和在心臟病理學中的作用差異性。
3 心臟GRKs水平和功能的調節機制
3.1 相互作用蛋白的調控 如前所述,GPCRs可激活其偶聯的G蛋白,完成信號轉導。例如,βAR激動劑與βAR結合后激活G蛋白,使 G蛋白解離為Gα和 Gβγ兩部分,這兩部分分別介導不同的信號轉導過程。但此過程受GRKs的調節。GRKs可使βAR發生磷酸化,磷酸化的βAR募集β-arrestins,導致βAR脫敏、內化。
強效刺激能使 GRKs與Gβγ或活化的 GPCRs結合。除此之外,GRKs的活性還能被可以牽制其催化活性的其它相互作用蛋白所調控,其中較為重要的一種是小窩蛋白(caveolin)。小窩蛋白可作為多種信號分子[其中包括β2AR和不同的絲裂原活化蛋白激酶(mitogen-activated protein kinases,MAPKs)以及G蛋白]的骨架蛋白,還可以幫助限制或區分信號轉導。GRK2可與存在于心臟的不同小窩蛋白亞型產生相互作用,尤其是在細胞膜小窩蛋白富集區若出現GRK2,小窩蛋白就易于與之產生相互作用。臨床病例報道,心力衰竭病人的心臟內,小窩蛋白水平有所降低;并且,小窩蛋白基因敲除小鼠中,心肌肥大和心肌收縮功能障礙的發病率明顯提高[10]。因此,GRK2-caveolin的相互作用的改變必然對心臟功能有重要調節作用。
最近發現,GRK2的另一個相互作用蛋白是Raf激酶抑制蛋白(Raf kinase inhibitor protein,RKIP)。RKIP被PKC磷酸化后,將會從Raf上轉移下來,同時與GRK2的結合增強[11]。RKIP水平的降低將導致GRK2活性增強,心肌細胞收縮反應性受損。
3.2 鈣離子結合蛋白的調控 細胞內鈣離子水平的變化與心力衰竭的發展有明顯相關性。不同細胞內,鈣離子敏感蛋白會與不同的GRKs產生相互作用,從而調節后者的活性[2]。GRK5對鈣離子結合的鈣調蛋白(calmodulin,CaM),即Ca/CaM的存在很敏感,CaM可以阻止GRK5靶向細胞膜,增加GRK5對可溶性底物的活性;相反,GRK2只有在CaM濃度很高時才會受影響。由于GRK5對CaM具有高親和力,在大多數類型細胞中當GPCR被激活而導致鈣離子濃度升高時,GRK5會優先被抑制。
3.3 磷酸化調控 雖然GRKs可以使多種GPCRs磷酸化,但是GRKs本身也會被多種激酶磷酸化,第二信使依賴的蛋白激酶C(protein kinase C,PKC)和蛋白激酶A(protein kinase A,PKA)可調節多種GRKs的活性和膜靶向性。這種相互作用在心臟疾病中尤為重要,因為在人類心力衰竭中,PKC和PKA的表達都是上調的[3]。PKC可使GRK2和GRK5磷酸化,但是二者產生的效應不同。GRK2磷酸化后,它對GPCRs的催化活性增強(可能是通過緩解鈣調蛋白引起的抑制效應);而GRK5磷酸化后,其催化活性顯著降低[2,5]。另一方面,被以與 Gαs偶聯為主的受體激活的PKA可直接磷酸化GRK2,該磷酸化并不影響GRK2自身的催化活性,而是增強GRK2與Gβγ亞基的結合,從而促進GRK2與細胞膜結合,增強其對活化受體的作用[2]。
GPCRs活化也可以引發非受體酪氨酸激酶的調節,如c-Src、MAPK。已有研究證明β2AR受刺激后,c-Src可以直接使GRK2磷酸化,進一步引起β2AR脫敏、內化,這一過程依賴β-arrestins招募c-Src的能力[12]。經酪氨酸磷酸化的GRK2對多種底物的活性也因此而增強;但從長期效應來看,該磷酸化可以促進對GRK2細胞水平的調節。
GRK2的活性還受細胞外信號調節激酶(extracellular signal-regulated kinase 1/2,ERK1/2)的調控。體外和原位實驗均表明,ERK1可以磷酸化GRK2的Gβγ結合區。MAPK磷酸化可顯著影響 GRK2和Gβγ的相互作用,從而抑制激酶的易位和對受體底物的催化活性。此外,GRK2被c-Src和MAPK磷酸化還能觸發后續的激酶降解。
3.4 轉錄和降解的調控 在一些病理條件下,不同的GRKs mRNA表達水平有所改變。已有研究報道,在主動脈平滑肌細胞,誘發血管收縮和心肌肥大的試劑(例如佛波醇)可顯著增強GRK2啟動子的活性[3]。相反地,促炎細胞因子,如白細胞介素1β(interleukin 1β,IL-1β)、腫瘤壞死因子 α(tumor necrosis factorα,TNF-α)或干擾素 γ(interferon γ,IFN-γ),可引起相反的效應。
GRKs穩定性的調節對其表達水平的調控來說是一個重要的機制。GRK2是一個短壽命蛋白,要經歷多聚泛素化并且可被c-Src和MAPK磷酸化,通過蛋白酶復合體通路降解[13]。此過程必須在一種穩定的包括β-arrestin的亞細胞環境中進行。
4 心血管疾病中GRKs水平和活性的變化
βARs是維持正常心臟功能的全程調節者,在人類心肌細胞,主要是由β1AR和β2AR兩個亞型通過與Gαs偶聯來介導正性變時、變力、變轉導的腎上腺素能效應。在心力衰竭病人心臟中,βAR密度和反應性降低,主要是因為β1AR出現表達下調。有研究證實,心臟功能性的β3AR(通過Gαi蛋白進行信號轉導)在心力衰竭的心肌中的表達是上調的[14]。
βARs通過激活其偶聯的G蛋白而發揮作用。鑒于GRKs調節GPCRs信號轉導的普遍性,心血管疾病中,βARs出現改變,GRKs相應地發生變化。尤其值得注意的是,GRK2和GRK5這2種亞型在心臟的表達和活性的改變。在動物的一些心力衰竭實驗模型中,GRK2和GRK5的表達和活性明顯增加。GRK2的表達和活性的增加還與高血壓、心肌肥大和心肌缺血有關[7,15]。最有趣的是,GRK2 水平常在心力衰竭發生之前就已經增加,當βAR信號和心室功能改善時才恢復正常[16-17]。在高血壓、心力衰竭等病理情形下,GRKs的結構和功能會發生顯著的改變,因此,GRKs可以作為一種治療靶點和評價心血管功能的潛在生物指標。
GRKs主要通過血管平滑肌上的GPCRs的磷酸化來調節血壓。GRKs家族成員中,GRK2、GRK3、GRK4和GRK5對血壓都有影響,尤其是GRK2和GRK5在高血壓癥狀中發揮了關鍵作用。心力衰竭時,βAR信號轉導被擾亂,交感神經系統受到持續的刺激,循環的兒茶酚胺類物質增加,結果導致βARs的脫敏和/或表達下調。在這一過程中,βARK1(GRK2)的表達也增加,進一步抑制了βAR的信號轉導。GRK2和GRK5過量表達會嚴重損害βAR信號轉導和β激動劑介導的心肌收縮功能。心力衰竭時,心肌 GRKs的活性提高了2~3倍,并且 GRK2 mRNA水平和蛋白表達水平均顯著增加[18]。在其它如血壓超負荷、心肌缺血和心肌梗塞等心臟病理情形下,也觀察到類似的GRK2活性或表達水平的上調。此外,應用βARK-ct(βARK的C末端類似物,可競爭性抑制βARK1轉位至細胞膜)抑制βARK1活性可使一些心力衰竭動物模型受損的心臟功能得以逆轉,增加存活幾率,與βAR阻斷劑具有協同作用[3,19]。這表明,心力衰竭時 βARK1 表達增加并非保護性機制。與GRK2不同的是,GRK5的一個突變體——GRK5 Leu41起到類似βAR阻斷劑的作用,能夠延緩去甲腎上腺素刺激受體的間隔,具有抑制心力衰竭的作用[18]。
5 結語和展望
心血管疾病是造成人群死亡的首要原因。可以預計,未來10~20年心血管疾病的患病率和死亡率還將上升。目前,心血管疾病的診斷和防治是世界性研究熱點。在研究這類疾病的病理生理機制時,發現更為有效的治療靶點具有十分重要的理論和現實意義。
GRKs與其相互作用蛋白的功能復合體(GRK“interactome”)展示了這些激酶對不同的信號轉導通路的重要性,GRKs蛋白水平或活性的變化會影響信號轉導通路的效率或特征。在病理環境中,GRKs水平和/或功能的改變是它們正常的合成、降解和調控失衡的結果。在不同心臟細胞類型中這些過程的特異調控還有待進一步詳細研究。另一方面,GRKs功能改變的結果可以在它們增加的復合體“interactome”中觀測到,此復合體包括Gαq、PI3K和非GPCR底物。這一理論,結合心血管疾病動物模型的應用可幫助解決以下問題:GRK水平的改變是心血管疾病的正常結果還是突如其來的因子;GRKs水平的增加是有益的還是有害的(或者取決于疾病階段)。我們可以這樣理解:使用GRKs作為潛在的生物標志是一種新的診斷程序,或者是一種基于GRKs活性、水平或特異相互作用調控的新的治療策略。
