長江中下游湖群大型底棲動物群落結構及影響因素
2013-12-08 12:40:33蔡永久姜加虎陳宇煒龔志軍
生態學報 2013年16期
蔡永久,姜加虎,張 路,陳宇煒,龔志軍
(中國科學院南京地理與湖泊研究所, 湖泊與環境國家重點實驗室, 南京 210008)
長江中下游湖群大型底棲動物群落結構及影響因素
蔡永久,姜加虎,張 路,陳宇煒,龔志軍*
(中國科學院南京地理與湖泊研究所, 湖泊與環境國家重點實驗室, 南京 210008)
長江中下游地區是我國淡水湖泊分布最為密集的區域,其中面積大于10 km2的湖泊總面積占相同級別中國淡水湖泊總面積的51.3%。目前對本地區湖泊大型底棲動物研究主要是關于單個湖泊或幾個湖泊之間的比較,將區域內湖泊作為一個整體來分析的研究較少。為揭示現階段長江中下游淺水湖泊底棲動物群落現狀及其主要影響因素,于2008年和2009年夏季對本地區5個湖群69個湖泊大型底棲動物和水化學進行了調查,并分析區域過程和局域環境條件在決定該地區底棲群落結構中的相對重要性。結果表明水體礦化度、電導率及氮磷指標在不同湖群間具有顯著差異,而高錳酸鹽指數、葉綠素a及營養狀態指數無顯著差異。密度方面,以寡毛類和搖蚊幼蟲為優勢類群的湖泊共46個,占總數量的66.7%,以螺類為優勢類群之一的湖泊16個,占總數量的23.2%;生物量方面,以螺類為優勢的湖泊數量最多(33個),占總數量的46.4%,但以寡毛類和搖蚊幼蟲占優勢的湖泊亦有27個,占總數量的39.1%,雙殼類僅在9個湖泊占據優勢。典范對應分析結果表明該地區底棲動物群落結構是局域環境條件和區域過程共同作用的結果,兩類因子共解釋了33.9%的底棲動物群落變異,其中局域環境因子占被解釋量的48.1%,空間變量占35.4%。空間變量較高的解釋量表明對整個長江中下游地區湖泊而言,區域過程對底棲動物的分布也起著非常重要的作用。
長江中下游; 湖群; 淺水湖泊; 局域和區域過程; 大型底棲動物
湖沼學研究開展以來,對于單個湖泊長期深入的研究,加深了對湖泊生態系統結構和重要過程的認識,這也是湖沼學研究開展以來湖沼學家研究的重點。近年來,湖沼學研究不再局限于單個湖泊,而將視野擴大到湖泊所在的地理區域和流域范圍,甚至全球尺度[1- 2]。湖泊比較是區域湖沼學的重要研究內容,多個湖泊的比較研究對湖沼學理論的發展具有重要意義[1],同時對湖泊生態系統的管理和保護也具有指導作用[3]。湖群的存在為比較湖沼學提供了得天獨厚的條件。Riera等將湖群定義為一塊湖泊富集的景觀,這些湖泊具有相同的起源、氣候和流域特征[4]。盡管如此,湖群內湖泊的物理、化學和生物特征也可能存在著顯著差異。了解這些差異背后的驅動因素是區域湖沼學的主要內容之一。在區域層次上,通過對多個湖泊的比較研究,更有助于找出湖泊間和湖群間各種差異的內在機制。然而,對于湖群的研究不能只考慮湖泊自身的環境特征,同時需要將湖泊所在的地理位置考慮在內[5]。
種庫假說認為:生物群落的結構不僅與所在地的環境條件和生態過程(如競爭和捕食)有關,也受其潛在的物種庫即區域種庫所左右。在區域和局域尺度上起主導作用的過程是不同的。捕食、寄生、競爭以及環境波動(即干擾)等生態過程的作用主要發生在局域水平上,而物種的長距離擴散、物種形成、大范圍的絕滅以及物種分布區的變化等生物地理學過程主要發生在區域尺度[6]。局域和區域過程共同作用于群落的結構和物種組成,并且兩者的相對作用大小隨群落類型、環境條件及尺度而變化[7]。自然界中,生物群落受到多尺度過程的影響,群落結構多尺度調控的概念來源于環境篩選。要想成功建立一個局域群落,每個物種必須通過多尺度下一系列的環境篩選[7]。在大尺度上,歷史事件和氣候限制形成了各區域獨特的物種庫,且只有物種庫的一部分物種最終能在局域群落中共存。對于湖泊生態系統來說,局域群落的物種必須通過區域、流域、湖群、生境等多個尺度環境因子的篩選。這些因子對物種的特性具有篩選作用,只有那些具有特定特性且能通過各級因子篩選的物種才能最終形成局域群落。
長江中下游平原是我國淡水湖泊分布最密集的核心區,且主要分布在長江兩側。根據其地理位置和流域特征,可分為洞庭湖群、江漢湖群、鄱陽湖群、華陽湖群、蘇皖湖群和太湖湖群[8],為開展區域湖沼學研究提供了天然條件。關于長江中下游地區湖泊底棲動物群落結構及其影響因素已有一些研究,但主要是關于單個湖泊或幾個湖泊之間的比較,將區域或流域內湖泊作為一個整體來分析的更少[9]。本文嘗試將長江中下游湖泊作為一個整體,假設底棲動物受到局域和區域過程的共同作用,通過比較分析不同湖群水體理化參數和底棲動物群落結構,并利用partial CCA解析區域過程和局域環境條件在決定本地區湖泊底棲動物群落結構中的相對重要性。
1 材料與方法
1.1 研究區域及底棲動物樣品采集
共調查大于10 km2的湖泊69個,分屬洞庭湖群、江漢湖群、鄱陽湖群、華陽湖群和太湖湖群(圖1)。采樣時根據湖泊面積大小,在敞水區布設若干采樣點(附錄1)。底棲動物定量采集用1/16 m2改良Peterson采泥器,每個樣點采集1—2次,泥樣經60目尼龍篩洗凈后,剩余物至于白磁盤中將底棲動物活體逐一挑出,樣本用10%福爾馬林溶液保存。樣品帶回實驗室鑒定至盡可能低的分類單元[10- 12],統計各個分類單元的數量,然后用濾紙吸去表面固定液,置于電子天平上稱重,最終結果折算成單位面積的密度和生物量。樣品采集時間為2008年6月至8月及2009年7月至8月。
1.2 環境因子的測定
采集底棲動物樣品時,同時現場測定環境指標并采集水樣分析水化學參數。透明度(SD)用賽氏盤測定,pH、電導率(Cond)和礦化度(TDS)于現場用YSI 6600 V2 多參數水質監測儀測定。采集水樣冷凍保存帶回實驗室,氨氮(NH4-N)、硝態氮(NO3-N)、正磷酸鹽磷 (PO4-P)用微量流動注射分析儀(Skalar-SA1000)測定,總懸浮顆粒物(TSS)、總氮(TN)、總磷(TP)、葉綠素a (Chl a)及高錳酸鹽指數(CODMn)的測定參考標準方法[13]。水生植被蓋度指數(Macrophytes):現場采樣時估計采樣點附近區域水生植物覆蓋情況,分為5個等級0,1—25%,26—50%,51—75%,76—100%,分別賦值為0—4。為評價各湖泊營養狀態,參照蔡慶華等改進的湖泊富營養化評價綜合模型計算各點的營養狀態指數(TSI)[14],計算公式為:
TSI = 0.540 TSI (Chl a) + 0.297 TSI (SD) + 0.163 TSI (TP)
式中,TSI (Chl a)、TSI (SD)、TSI (TP)按照 Carlson 提出的營養狀態指數計算方法獲得,計算公式分別為:
TSI (Chl a)=10×(6-(2.04-0.68 (ln (Chl a)))/ln2)
TSI (SD)=10×(6-ln(SD)/ln2)
TSI (TP)=10×(6-ln(48/TP)/ln2)
式中,Chl a、SD、TP 單位分別為μg/L、m、μg/L。
1.3 數據分析
數據分析前,各湖泊多個采樣點的環境因子數據和底棲動物數據進行平均,理化數據進行Z標準化(平均值0,標準差1),物種數據進行對數轉換。
1.3.1 湖群比較分析:
由于不同湖群湖泊數量相差較大,且數據轉換后仍不能滿足方差分析的要求,故采用Kruskal-Wallis非參數檢驗比較各湖群水體理化參數的差異性,多重比較采用Games-Howell方法。由于部分環境因子之間存在高度相關性,利用主成分分析對環境因子進行降維分析。
運用Bray-Cutis相似性指數分析不同湖群湖泊底棲動物群落相似性,并利用相似性分析 (ANOSIM) 檢驗各湖群湖泊底棲動物群落差異顯著性,多元分析采用對數轉換后的密度數據,分析軟件用PRIMER 5.0[15]和PAST 2.01[16]。
1.3.2 影響因素解析
用partial CCA解析局域環境條件和區域過程對本地區底棲動物群落的影響,分析時需構建兩個解釋變量矩陣,分別是局域環境因子和空間變量[17]。局域環境因子矩陣 [E],包括水體理化因子、TSI指數、水生植被蓋度指數以及主成分分析中各湖泊在前二個主成分的得分,將主成分作為環境因子的原因是許多環境因子之間存在共線性,而主成分作為多個因子的線性組合,在實際分析中可能會更好地反映出環境梯度的影響。空間變量利用湖泊的經緯度為基礎構建,鑒于空間過程的影響并不是簡單的線性格局,也包括其它復雜的過程(如隔離、斑塊),在這里采用9個空間變量組合,分別為X、Y、XY、X2、Y2、X2Y、XY2、X3、Y3,X和Y分別為各湖泊采樣點經度和緯度的平均值(附錄1)。同時考慮到不同湖群底棲動物可能具有顯著差異,將5個湖群作為啞變量進入分析,經緯度變量和啞變量共同構成空間變量矩陣 [S]。CCA分析時,分別用局域環境因子和空間變量作為解釋變量,物種數據為對數轉換后的密度數據,并剔除出現率小于10%的物種,因子篩選采用向前引入法(α=0.05),基于Monte Carlo permutation檢驗(n=9999)。
根據Borcard的方法[17],將物種數據矩陣的總變異分解成不同組分:1)將篩選后環境因子和空間變量一起進入CCA分析,計算出總解釋量[E+S];2)環境因子獨立解釋量[E|S],分析時環境因子為解釋變量,空間變量為協變量;3)空間變量獨立解釋量[S|E],分析時空間變量為解釋變量,環境變量為協變量,排序結果的顯著性用Monte Carlo permutation 檢驗(n=9999)。空間變量和環境變量共同解釋部分[E∩S]計算公式為[E+S]-[E|S]-[S|E],未解釋部分為1-[E+S],分析軟件采用CANOCO 4.5[18]。
2 結果
2.1 水體理化特征
表1為不同湖群水體理化參數和營養狀態指數,非參數分析結果表明電導率、礦化度、TSS及水體中氮磷濃度在各湖群間具有顯著差異。電導率和礦化度呈現出一致的變化趨勢,其中鄱陽湖群最低(平均值分別為150.4 μs/cm、99.04 mg/L),洞庭湖群、江漢湖群和華陽湖群處于中間水平,太湖湖群最高,其電導率和礦化度的平均值分別為532.03 μs/cm和345.86 mg/L。水體中TN和TP的共同變化特征是太湖湖群顯著高于其他湖群(除和鄱陽湖群TP無顯著差異),洞庭湖群和華陽湖群水體TN和TP含量均較低(表1),水體中CODMn、Chl a及TSI指數在各湖群間無顯著差異。
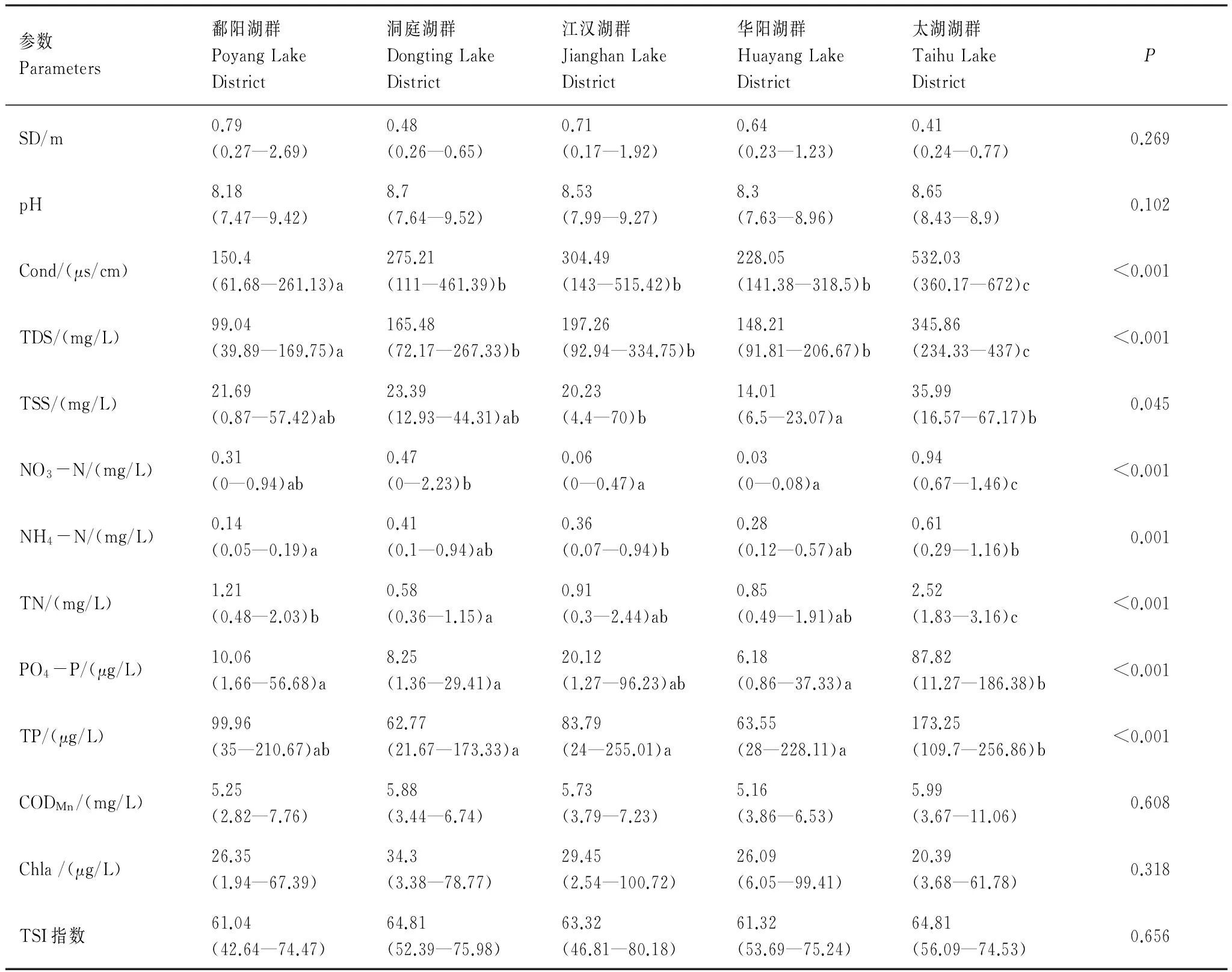
表1 各湖群水體理化特征及非參數Kruskal-Wallis檢驗結果
不同字母間表示差異顯著

圖1 長江中下游湖泊空間分布圖 Fig.1 Spatial distribution of the studies lakes in the middle of lower reaches of the Yangtze River
Table2StatisticalcharacteristicsofPCA,includingtheeigenvalues,percentageofvarianceexplained,andcorrelationsforeachenvironmentalvariablewiththefirst3principlecomponents
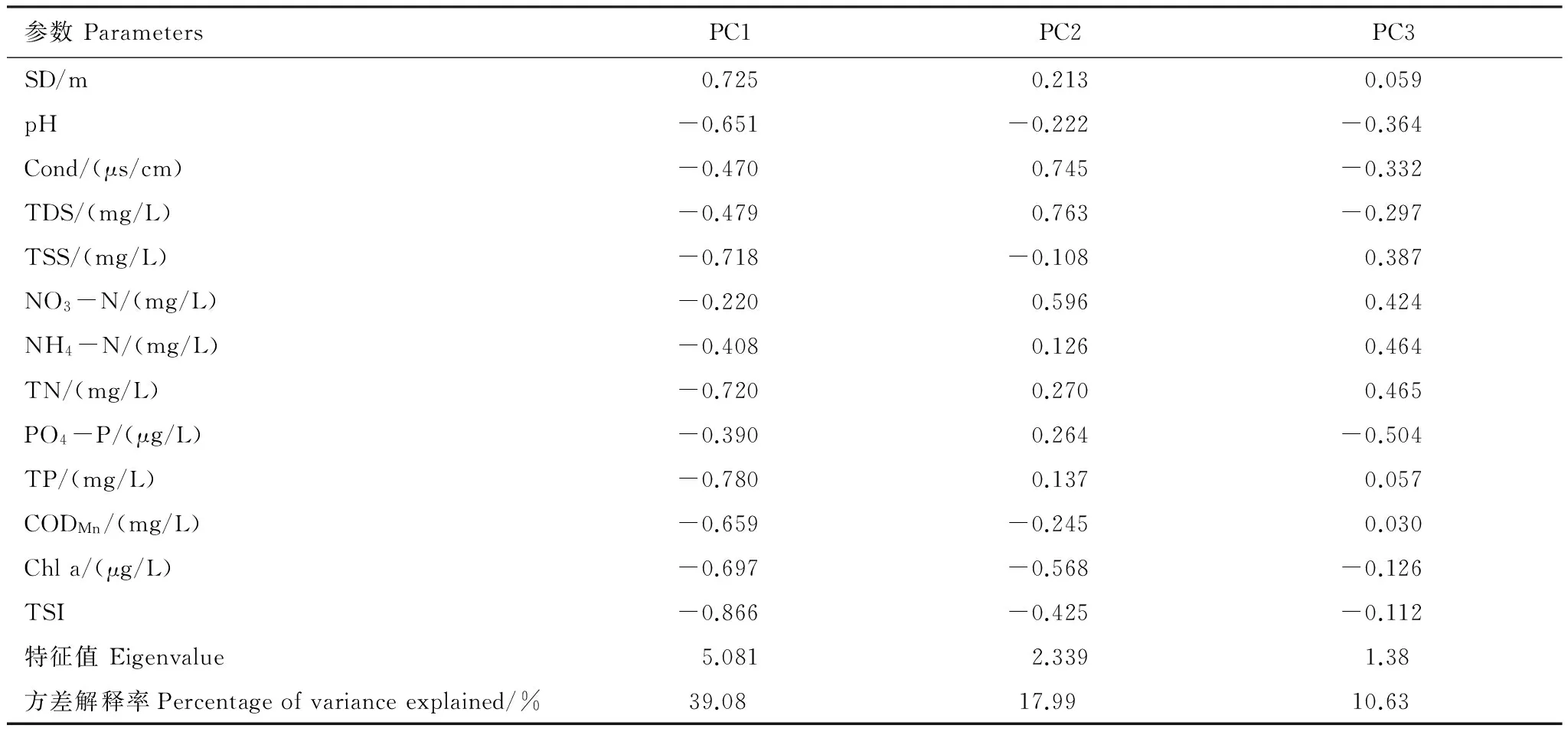
參數ParametersPC1PC2PC3SD/m0.7250.2130.059pH-0.651-0.222-0.364Cond/(μs/cm)-0.4700.745-0.332TDS/(mg/L)-0.4790.763-0.297TSS/(mg/L)-0.718-0.1080.387NO3-N/(mg/L)-0.2200.5960.424NH4-N/(mg/L)-0.4080.1260.464TN/(mg/L)-0.7200.2700.465PO4-P/(μg/L)-0.3900.264-0.504TP/(mg/L)-0.7800.1370.057CODMn/(mg/L)-0.659-0.2450.030Chla/(μg/L)-0.697-0.568-0.126TSI-0.866-0.425-0.112特征值Eigenvalue5.0812.3391.38方差解釋率Percentageofvarianceexplained/%39.0817.9910.63
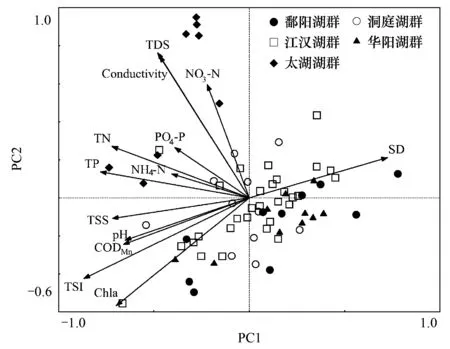
圖2 69個湖泊水體理化參數主成分分析 Fig.2 Principal components analysis (PCA) of physicochemical parameters in the 69 studied lakes
2.2 不同湖群底棲動物類群組成
圖3為不同類群底棲動物在各湖泊總密度和總生物量中所占比重。密度方面,鄱陽湖群中,赤湖以搖蚊幼蟲和螺類為優勢類群,雙殼類(主要是河蜆)和寡鰓齒吻沙蠶在珠湖和軍山湖比重較高,鄱陽湖的優勢種為河蜆,其他中小型湖泊為寡毛類和搖蚊幼蟲共同主導。洞庭湖群中,搖蚊幼蟲在芭蕉湖和珊珀湖占據絕對優勢,城市湖泊岳陽南湖完全被寡毛類主導(97.39%),平均密度為2192個/m2,黃蓋湖為寡毛類和搖蚊幼蟲共同主導,螺類在湖南東湖等6個湖泊中占據一定比重。根據優勢類群,江漢湖群的30個湖泊可分為兩組,其中以寡毛類和搖蚊幼蟲占據優勢的湖泊有22個,而以螺類和寡毛類及搖蚊幼蟲占主導的湖泊有8個,其中大冶湖、斧頭湖和洪湖螺類所占比重較高,分別為95.71%、71.88%和63.34%。華陽湖群類群組成較為多樣,6個中小型湖泊(武山湖、太白湖、武昌湖、升金湖和破崗湖)以寡毛類和搖蚊幼蟲為優勢類群,菜子湖和白蕩湖的優勢類群為螺類,大型湖泊龍感湖和泊湖的優勢種為寡鰓齒吻沙蠶。太湖湖群中除澄湖和淀山湖的優勢種分別為鉤蝦和寡鰓齒吻沙蠶外,其它湖泊優勢類群均為寡毛類和搖蚊幼蟲。對所有湖泊進行分析發現,以寡毛類和搖蚊幼蟲為優勢類群的湖泊共46個,占所調查湖泊的66.7%,以螺類為優勢類群之一的湖泊16個,占總數量的23.2%,雙殼類和其它種類占優勢的湖泊共7個(表3)。
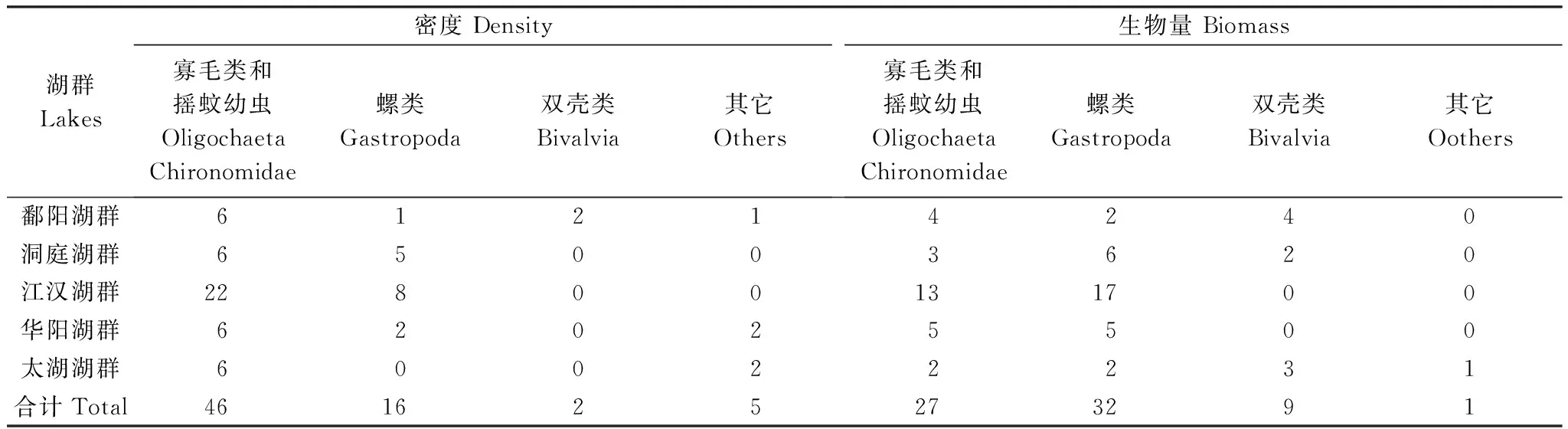
表3 不同類群底棲動物占優勢湖泊數量在各湖群的分布
生物量方面,雖然螺類和雙殼類個體較大,在多數湖泊對總生物量貢獻較大,但寡毛類和搖蚊幼蟲的貢獻率在部分湖泊亦較高。鄱陽湖群中,以寡毛類和搖蚊幼蟲占優勢的湖泊有4個,螺類和雙殼類占優勢的湖泊分別為2個和4個。洞庭湖群大部分湖泊為螺類(6個)和雙殼類(2)所主導,其余3個湖泊為搖蚊幼蟲所主導。江漢湖群螺類為優勢的湖泊有17個,寡毛類和搖蚊幼蟲為主的湖泊13個,數量大致相當。華陽湖群中5個湖泊為螺類所主導,2個為寡毛類和搖蚊幼蟲共同主導,其余3個湖泊以這3個類群共同主導。太湖湖群中各類群占優的湖泊數量相當,其中鉤蝦對澄湖生物量貢獻較大。整體分析發現,雖然以螺類為優勢的湖泊數量最多(32個),占總數量的46.4%,但以寡毛類和搖蚊幼蟲占優勢的湖泊亦較多(27個),占總數量的39.1%,雙殼類僅在9個湖泊占據優勢(表3)。

表4 不同湖群湖泊大型底棲動物群落Bray-Curtis相似性分析
*下方為不相似性百分比,上方為顯著性水平
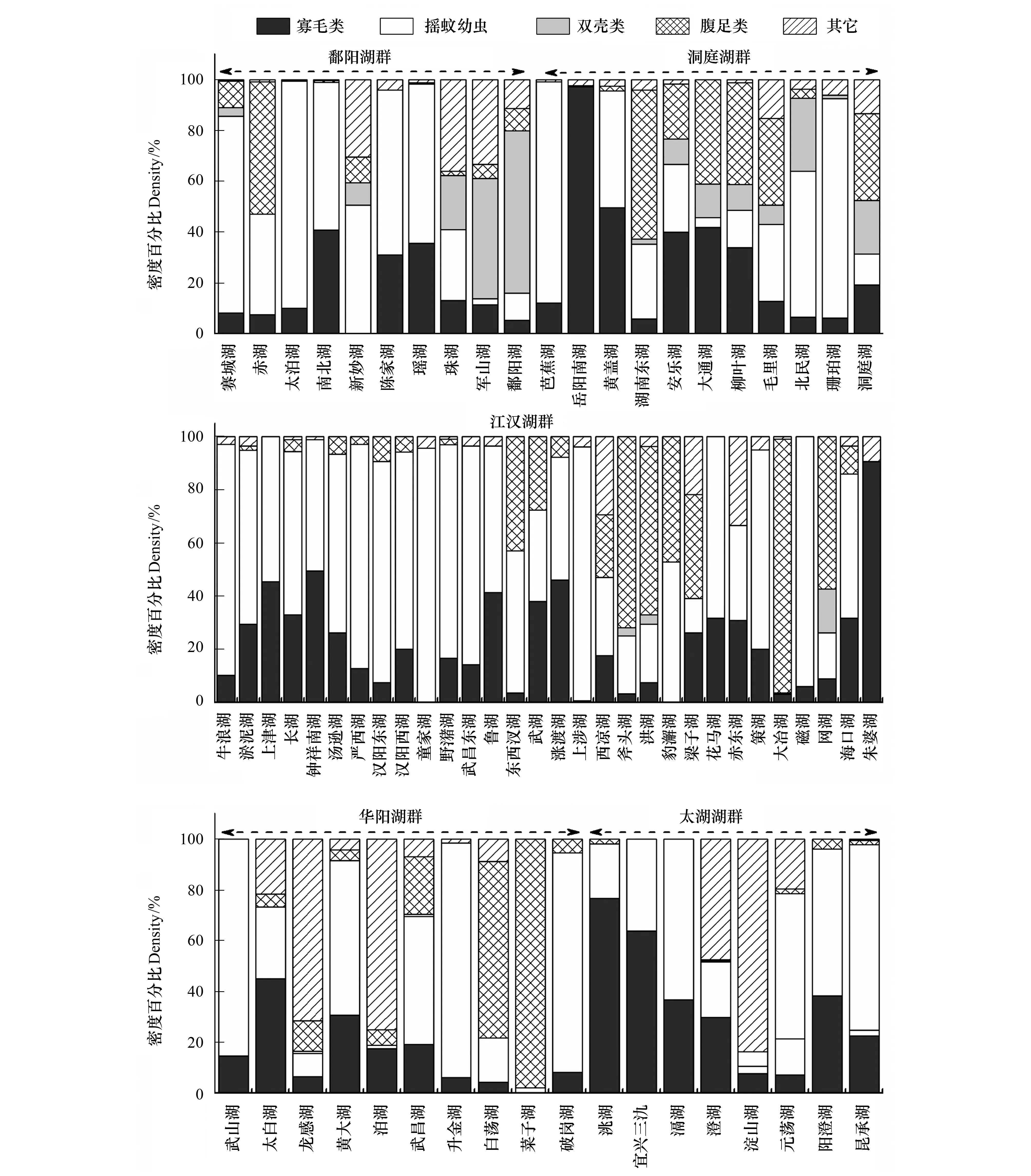
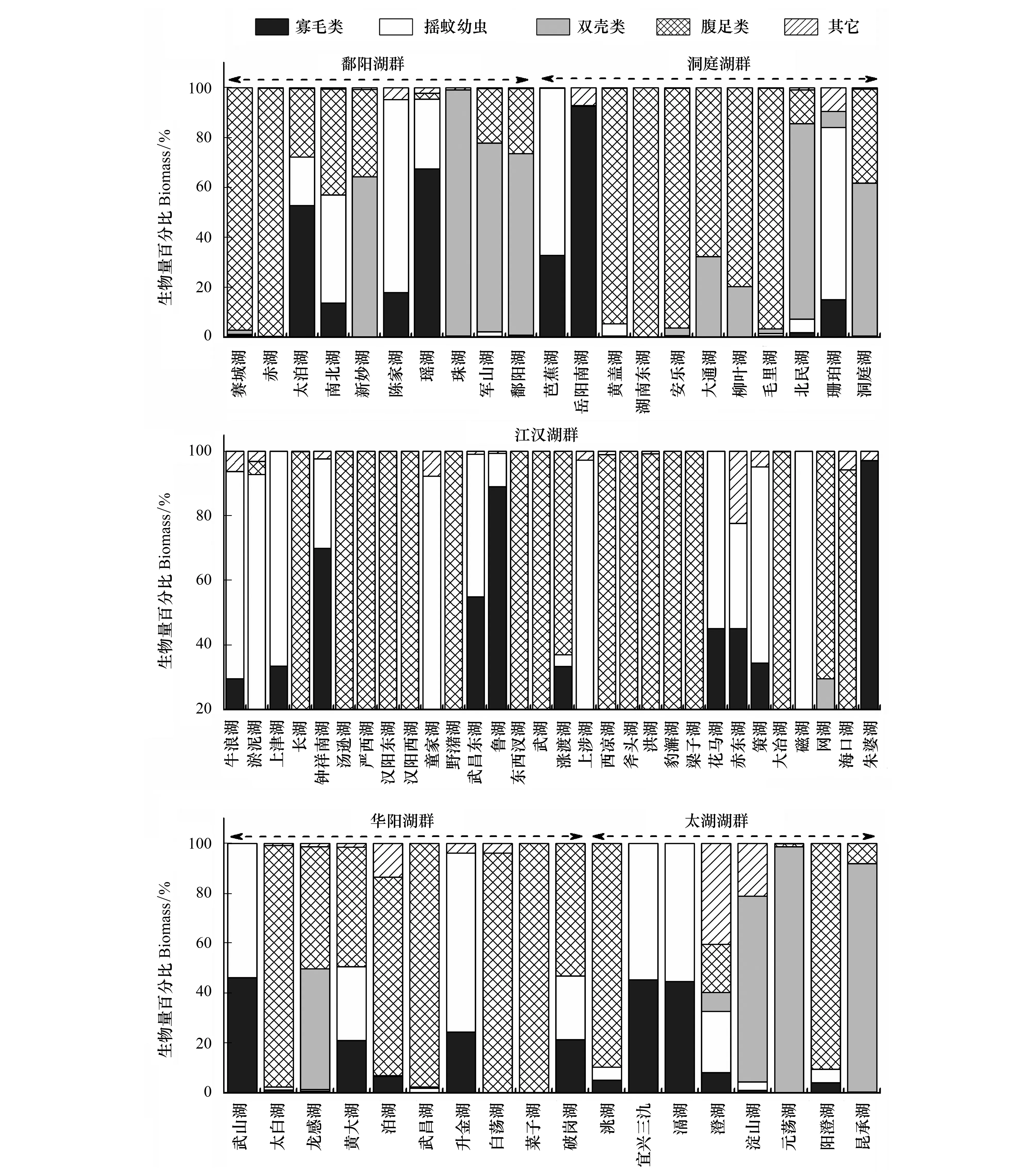
圖3 各湖群4種常見類群大型底棲動物在總密度和生物量中所占比重Fig.3 Relative abundance and biomass of the four common taxonomic groups in the five lake districts
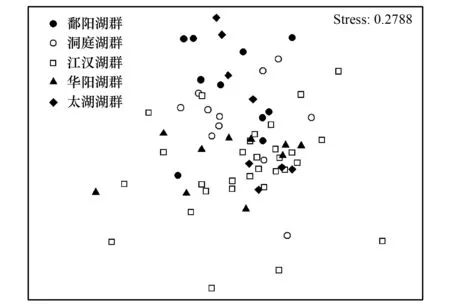
圖4 基于69個湖泊大型底棲動物密度的Bray-Curtis相似性系數的非度量多維標度排序圖Fig.4 Ordination plot of macrozoobenthic communities based on Bray-Curtis similarity coefficients in the 69 lakes
Bray-Curtis相似性系數結果表明不同湖群底棲動物群落不相似性百分比較高,介于67.52%至73.48%(表4)。ANOSIM分析結果表明鄱陽湖群和洞庭湖群與華陽湖群和太湖湖群底棲動物群落組成具有顯著差異,鄱陽湖群和洞庭湖群之間無顯著差異;江漢湖群除與鄱陽湖群具有顯著差異外,與其它湖群均無顯著差異,這可能是因為江漢湖群湖泊數量較多,按底棲優勢類群可分為兩組,將江漢湖群湖泊分為兩組進入分析時發現,所有湖群間均具有顯著差異(鄱陽湖群和洞庭湖群間除外)。NMDS排序分析顯示除江漢湖群部分湖泊分布較為集中外,其它湖群湖泊空間聚集現象并不明顯,這表明湖群內部底棲動物群落變化也較大,湖群因子不能很好地反映本區域湖泊底棲動物群落的變化。
2.3 影響因素解析
2.3.1 局域環境條件
典范對應分析表明局域環境因子中TSI指數、Macrophytes、Chl a、TDS 及PC2與69個湖泊底棲動物群落關系最顯著。第一軸和第二軸的特征值較大,分別為0.198和0.146,分別解釋了10.2%和7.4%的物種數據變異及43.9%和32.3%的物種-環境關系變異,第三軸解釋了3.0%的物種數據變異和12.9%的物種-環境關系變異,第四軸特征值和解釋率都相對較低,前3軸基本反映了底棲群落與局域環境因子的關系。蒙特卡羅檢驗結果表明CCA前3個排序軸均顯著(P1,2lt;0.001,P3=0.008),第四軸不具有顯著性(P4= 0.53)。第一軸與TSI、Macophytes、Chl a相關性較高,其主要反映了水體營養狀態及生境的異質性;第二軸與Chl a相關性最高,第三軸與TDS和PC2呈顯著正相關(表5和圖5)。從各因子進入CCA分析的順序及其解釋量可以看出,TSI指數、Macrophytes和Chl a 解釋量相對較高,而TDS和PC2相對較低。物種與環境因子的排序圖顯示水生蓋度指數與螺類相關性較高,TSI和Chl a與搖蚊科幼蟲及顫蚓科的種類相關性較高,PC2則與鉤蝦、雙殼類及寡鰓齒吻沙蠶呈正相關關系(圖5)。
2.3.2 空間過程
面積適宜性主要考慮對關鍵生境及敏感生態系統的保護需要,特別是目標種群產卵生物量的積累,其大小和范圍取決于保護目的、關鍵生境質量和數量、資源利用程度、其它管理工具的有效性以及目標生物種群特征等因素。邊界和面積可依照海域地形特征、生態地理屬性,以及目標種群的擴散類型及開發程度等因素來確定。
利用空間變量和湖群啞變量作為解釋因子,CCA分析最終篩選出5個空間變量組合最大程度地解釋了底棲動物群落變異,而5個啞變量并沒有在組合之內。蒙特卡羅檢驗結果表明CCA分析前兩個排序軸均顯著(P1,2lt; 0.01),第三軸和第四軸不具有顯著性(P3= 0.08,P4= 0.11)。相比于局域環境因子的CCA分析,空間變量的CCA分析前兩軸特征值較小,分別為0.159和0.081,共解釋了12.3%的物種數據變異和66.7%的物種-環境關系變異(表6)。空間變量與第一軸相關性較低,第二軸與變量Y、X3及X相關性較強。從排序圖中可以看出,各湖群湖泊在排序圖中能較好的區分開來,其中洞庭湖群湖泊位于排序圖的下方,江漢湖群湖泊主要位于排序圖左方,鄱陽湖群湖泊位于排序圖右側,太湖湖群湖泊較為分散地分布區排序圖上方,華陽湖群湖泊位于排序圖中心區域(圖6)。
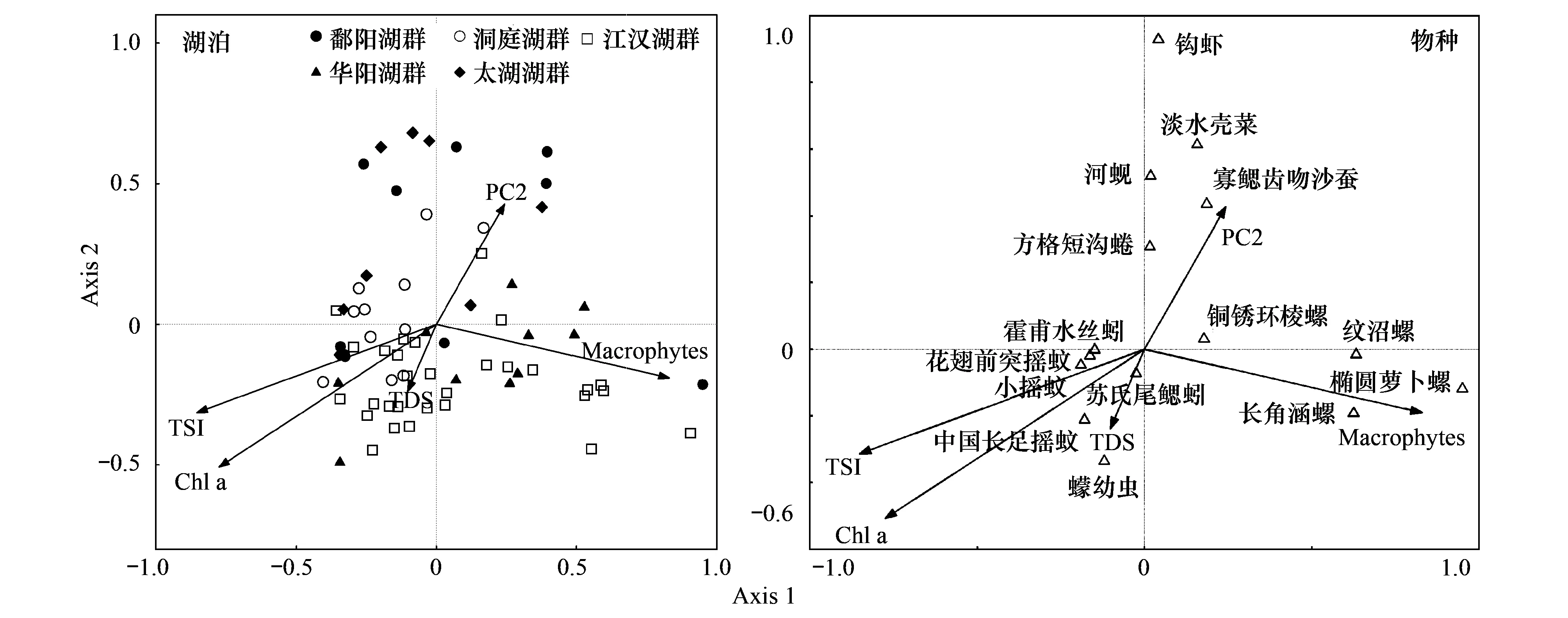
圖5 69個湖泊大型底棲動物群落與局域環境因子的典范對應分析排序:湖泊,物種,僅顯示與環境因子相關性較高的種類Fig.5 CCA biplots of macrozoobenthic communities in 69 lakes, showing the first three axes and the most strongly correlated local environmental variables: lakes, species, only show species with high correlations with environmental variables
2.3.3 方差分解
局域環境因子和空間變量對底棲動物群落變異的總解釋量為33.9%,其中局域環境因子獨立解釋的部分為16.3%,稍大于空間變量的解釋率12.0%,由空間過程控制的局域環境因子解釋量最小,僅為5.6%。partial CCA分析發現當以空間變量為協變量時,PC2和TDS并不具有顯著性,表明電導率和礦化度主要受湖泊所處地理區域控制。方差分解結果表明長江中下游地區湖泊底棲動物并不簡單受到局域環境因子的影響,區域過程在調節底棲群落結構中同樣起著重要的作用。
表5底棲動物群落與局域環境因子的典范對應分析結果,各因子括號中的數值分別代表因子篩選時進入CCA分析的順序和對底棲群落變異的解釋量
Table5Summarystatisticsforthefirstfouraxesofthecanonicalcorrespondenceanalysis(CCA)ofspecies-environment,Valuesaftertheenvironmentalvariablesrefertotheentryranksofvariablesinforwardselectionandthepercentageofvarianceexplainedinspeciesdata

軸Axes1234特征值Eigenvalues0.1980.1460.0580.029種類-環境相關性Species-environmentcorrelations0.8320.760.7460.519物種數據累積變化百分比Cumulativepercentagevarianceofspeciesdata10.217.620.622.1物種-環境關系累積變化百分比Cumulativepercentagevarianceofspecies-environmentrelationship43.976.289.195.5TSI(1,8.37%)-0.853-0.3140.0420.380Macrophytes(2,7.70%)0.830-0.1910.1220.505Chla(3,8.26%)-0.775-0.508-0.1290.349TDS(4,3.34%)-0.102-0.2380.965-0.028PC2(5,4.67%)0.2440.4280.851-0.102
表6底棲動物群落與空間變量的典范對應分析結果,各因子括號中的數值分別代表因子篩選時進入CCA分析的順序和對底棲群落變異的解釋量
Table6Summarystatisticsforthefirstfouraxesofthecanonicalcorrespondenceanalysis(CCA)ofspecies-environment,Valuesaftertheenvironmentalvariablesrefertotheentryranksofvariablesinforwardselectionandthepercentageofvarianceexplainedinspeciesdata

軸Axes1234特征值Eigenvalues0.1590.0810.0560.046種類-環境相關性Species-environmentcorrelations0.7980.7350.7310.563物種數據累積變化百分比Cumulativepercentagevarianceofspeciesdata8.212.315.217.6物種-環境關系累積變化百分比Cumulativepercentagevarianceofspecies-environmentrelationship44.266.782.295Y(1,4.47%)-0.4260.7220.5100.166X2(2,4.21%)0.4770.3720.783-0.002X3(3,3.95%)0.2860.7920.417-0.220XY(4,2.98%)0.3220.1560.783-0.157X(5,3.59%)0.2270.8390.015-0.046
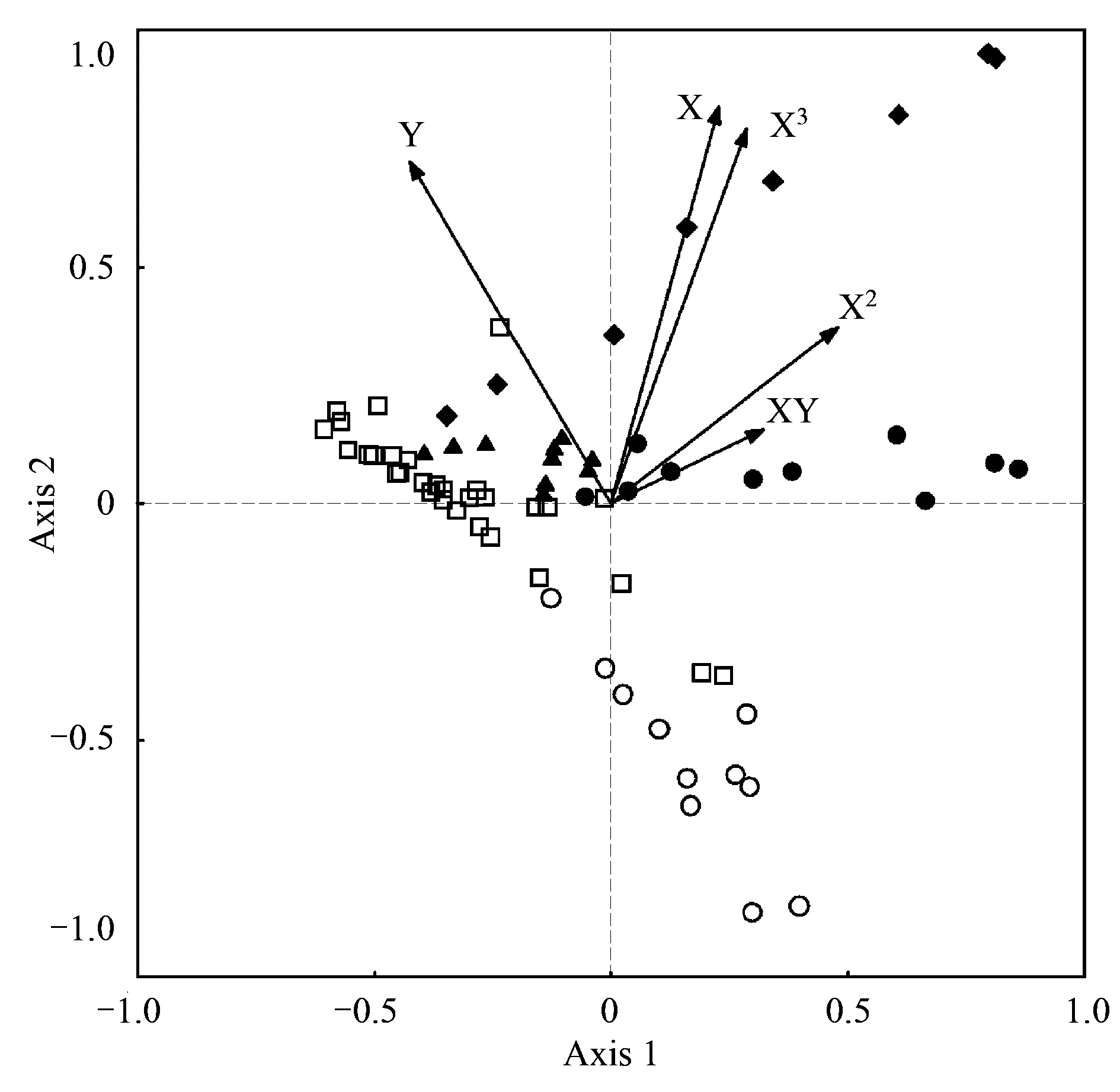
圖6 69個底棲動物群落與空間變量的典范對應分析排序圖Fig.6 CCA biplots of macrozoobenthos communities in 69 lakes, showing the first two axes and the most strongly correlated spatial variables
3 討論
3.1 水體理化特征
電導率和礦化度在不同湖群間差異顯著。電導率和礦化度是湖泊水化學的重要基礎參數,它們直接反映了水體的離子總量,又可間接反映湖泊鹽類物質積累的環境條件。礦化度受不同湖群所處流域的自然限制,因而呈現出明顯的區域差異。在五個湖群中,鄱陽湖群礦化度最低,所有湖泊的最低值也出現在該湖群的軍山湖(39.9 mg/L),相比之下,太湖湖群湖泊的電導率都較高,如淀山湖、元蕩湖和澄湖都超過了400 mg/L。湖群之間礦化度的顯著差異可能與地質背景、流域降水及湖泊換水周期有一定的關系。
長江中下游地區的年降水量一般都在1000 mm以上,但其在各大湖群也呈現出地區性的分異,如鄱陽湖地區的鄱陽縣年降水量比洞庭湖地區的岳陽、常德、沅江多250—300mm,而后者又比太湖地區的吳縣東山及上海多200—300mm,表明鄱陽湖區年降水量最豐、洞庭湖區次之,太湖地區降水量相對最少[8]。另一方面,鄱陽湖群降水豐沛,水系發達,它上承贛、撫、信、饒、修5條河流,來水量豐富,水體交換快,可溶性鹽類不易因水蒸發而積累。此外,鄱陽湖流域土壤類型多為紅壤,而紅壤呈酸性,含鹽量較低[19],故入湖河流的礦化度也較低,這些因素共同造就了鄱陽湖群成為本區礦化度最低的湖群。
湖泊換水周期也直接決定了離子在湖泊水體中的積累。鄱陽湖、洞庭湖和石臼湖是目前長江中下游僅有的天然通江湖泊,雖分屬不同的湖群,地質背景差異較大,但礦化度均較低,分別為73、155及122 mg/L。這主要是因為它們與長江的水體交換頻繁,換水周期較短,稀釋作用較為明顯,因而礦化度也較低。華陽湖群湖泊多為大中型湖泊,部分湖泊季節性與長江相通,其礦化度也較低。
研究發現反映水體營養鹽含量的指標在湖群間存在顯著差異,水體中營養鹽含量除與地質背景有關,其受人類活動影響更多。華陽湖群總氮和總磷水平均最低,這是因為該地區湖泊多為大中型湖泊,且季節性與長江相通,不適宜進行施肥養殖,養殖方式多為人放天養,因此其氮磷含量較低。鄱陽湖群、洞庭湖群和江漢湖群除有幾個大型(鄱陽湖、洞庭湖)和大中型湖泊(洪湖、軍山湖、梁子湖)外,其他湖泊均面積較小,湖泊多被集體、個人或公司承包經營,養殖過程中為追求產量,多采用投餌投肥的養殖方式,因此營養鹽含量較高,且3個湖群之間無顯著差異。太湖湖群不僅擔任著養殖的功能,其周邊工農業發達、人口眾多,污水不可避免地進入湖泊水體,過去幾十年中一直接納生產生活污水,因此營養鹽含量最高。研究發現Chl a在湖群間不具有顯著差異,表明其并不簡單受水體營養鹽含量影響,Chl a主要反映了浮游植物的生物量,其它因素(如魚類和浮游動物捕食、氣候條件等)對其控制作用也不可忽視,可惜未能獲得這方面數據。
3.2 影響因素解析
3.2.1 局域環境因子
CCA分析中篩選出5個局域環境因子與底棲動物群落關系最密切。其中TSI指數和Chl a反映了水體的營養水平,Macrophytes主要反映了生境的異質性程度,PC2主要代表了電導率和礦化度,先前分析表明電導率和礦化度不僅僅受地質背景影響,更反映了湖泊的換水周期和稀釋條件。從排序圖中可以看出,PC2和Macrophytes均位于TSI指數和Chl a的反方向,CCA分析中相關系數介于-0.370至-0.542間,換水周期直接影響了水體的營養狀態和Chl a濃度,而水草覆蓋度較高的湖泊水體營養狀態一般較低,說明5個環境因子之間是相互關聯的。因此,篩選出的環境因子主要反映了水體的營養狀態和生境的異質性。關于水體營養狀態和水生植被對底棲動物群落結構影響的研究較多,長江中下游地區湖泊水體營養狀態較高,一方面與其較高的本底值有關,另一方面是人類活動的結果,本文研究的湖泊營養水平均已到達或超過中營養水平。富營養化對底棲動物的影響往往是間接的,主要是改變了生境條件和食物質量,如氧含量、底質粒徑和異質性、有機質含量、食物中元素比例和必需脂肪酸含量[20]。水生植被的存在為底棲動物提供了棲息和躲避捕食的場所,同時其復雜的構件結構也增加了可供多個物種共存的空間[21]。
3.2.2 區域過程
研究淡水底棲動物分布的一個重要方面就是分析地理因素和環境因素對其分布的限制作用。某物種在特定生境的缺失可能是該物種并沒有擴散到該生境,也可能是它已經擴散到了該地區,但是由于環境條件不利于其存活或競爭,從而導致它沒有繁衍下去。相反,一個物種在特定地點的出現可能是它競爭優勢更強,也有可能是它到了這里,而它潛在的競爭對手并沒有擴散到這里。擴散受多種因素的影響,如高山、距離、擴散方式、種群大小[7]。一個種群在新生境的競爭和繁衍能力取決于它對新環境的適應能力。許多底棲動物是世界性分布種類,而另一些種類則僅出現在特定的氣候區或地理區域。
生態學研究中,關于底棲動物群落大尺度空間格局的研究較少。推測起來,區域過程的影響在小范圍地理區域可以忽略。然而,對整個長江中下游地區的研究不能忽視區域過程的影響。本文研究結果表明超過40%的被解釋的底棲動物群落變化可歸結于空間因素,盡管部分空間因素解釋量可能是未測量環境因子共同的解釋量,但是如此高的比重表明至少部分物種受空間過程影響更明顯。事實上,在調查中也發現部分物種值出現在某些特定湖泊,如耳河螺(Rivulariaauriculata)和雙龍骨河螺(R.bicarinata)只在洞庭湖群和鄱陽湖群湖泊中采集到,寡鰓齒吻沙蠶在調查湖泊中的分布上限為鄱陽湖群,其主要分布在長江下游兩側的湖泊中,而在中游的洞庭湖群和江漢湖群均未采集到,寡鰓齒吻沙蠶為海洋河口性種類,可見其在本區湖泊中的分布受到空間擴散的限制。在CCA分析中,篩選的空間變量包括X、Y、XY、X2及X3,表明底棲動物群落并不簡單受空間經度和緯度梯度的影響,而更復雜的景觀格局(表現為高次方的空間變量),如斑塊、隔離等在決定底棲群落結構中也起著重要作用。CCA分析中,5個湖群啞變量并沒有進入最終的CCA模型,可能是因為啞變量與某些空間變量存在相關,單獨分析每個啞變量對底棲群落的解釋能力發現,4個湖群啞變量(除華陽湖群)均與底棲群落顯著相關,解釋率介于3.08%—4.36%,與空間變量的解釋率相當,這種結果說明湖群并不是解釋本區底棲群落變化的最佳空間變量,這可能是因為部分湖泊在歷史上同屬于一個湖群,如洞庭湖群和江漢湖群由云夢澤演化形成,而鄱陽湖群和華陽湖群在歷史上均是彭蠡澤的一部分,因此它們的物種庫可能存在重疊。
3.2.3 區域過程和局域環境條件的相對重要性
底棲動物受到從局域到區域多重因素的影響,本研究中,大尺度的空間變量和局域環境因子對解釋底棲動物群落變化都非常重要。淡水生物群落的地理格局常對應于大尺度的歷史、氣候和地理因素。一些學者認為大型無脊椎動物群落結構變化與大尺度的區域過程關系更密切,其次才是局域環境條件[22]。在極地溪流和湖泊中,底棲群落變化經常與緯度梯度聯系密切[23- 24]。區域過程和局域環境條件對生物群落的影響與尺度相關,一般而言,隨著研究范圍的減小,局域環境因子的解釋量逐漸增加,而空間變量和受空間限制的環境因子的解釋量卻逐漸降低。本地區湖泊雖覆蓋了很大的地理范圍,但是在決定底棲群落結構時局域環境條件比大尺度的空間趨勢更重要。這可能與本地區湖泊的特征有關,所調查的湖泊雖屬于不同湖群或流域,但均為整個長江流域的一部分,多數湖泊的形成都與長江有密切的關系,且許多物種在該區域是廣泛分布的,可能并沒有受到擴散的限制,更多的是受到局域環境條件的限制。Johnson和Goedkoop對瑞典湖泊[25]及Sandin和Johnson對瑞典溪流[26]底棲動物的研究也發現局域環境因子較空間變量更重要,這可能與其較小的研究區域范圍有關。本文中與緯度相關的區域過程對本區湖泊底棲動物群落的影響較小,這可從CCA分析篩選出的空間變量組合看出,其原因是本區湖泊緯度跨度小,湖泊主要是沿經度上分布,而緯度對生物種群的影響一般較經度更明顯。另外,Donohue 等研究發現富營養化導致底棲動物群落在局域和區域尺度上均趨于同質化[27],而本區湖泊營養狀態普遍較高,可能導致該區湖泊底棲群落趨于同質化,從而降低了空間變量的解釋率。
本研究中,CCA解釋的底棲動物群落變異量較低,共為33.9%,?kland指出低的解釋率所提供的信息也是非常有價值的[28]。低解釋率在生態學數據中是很常見的,主要是因為生態學數據繁雜且龐大,另外野外調查取得數據也不可避免地與實際情況存在差異,特別是物種數據,由于野外工作條件及人力財力的限制,不可能獲得所有物種的數據。本文中局域環境因子的解釋也較低,獨立解釋率為16.3%,這主要是因為測定的因子數量較少,本文使用的因子主要是水體的理化參數,而有關沉積物和生境的數據未能獲得,如沉積物有機質含量、粒徑組成、底質類型、魚類潛在捕食壓力、與河流和其它湖泊的連通性。這些因子都對底棲動物的組成、密度和生物量有顯著影響,因此導致局域環境因子的解釋率較低。
致謝:周方正、鄧建明、吳召仕幫助采集野外樣品, 特此致謝。
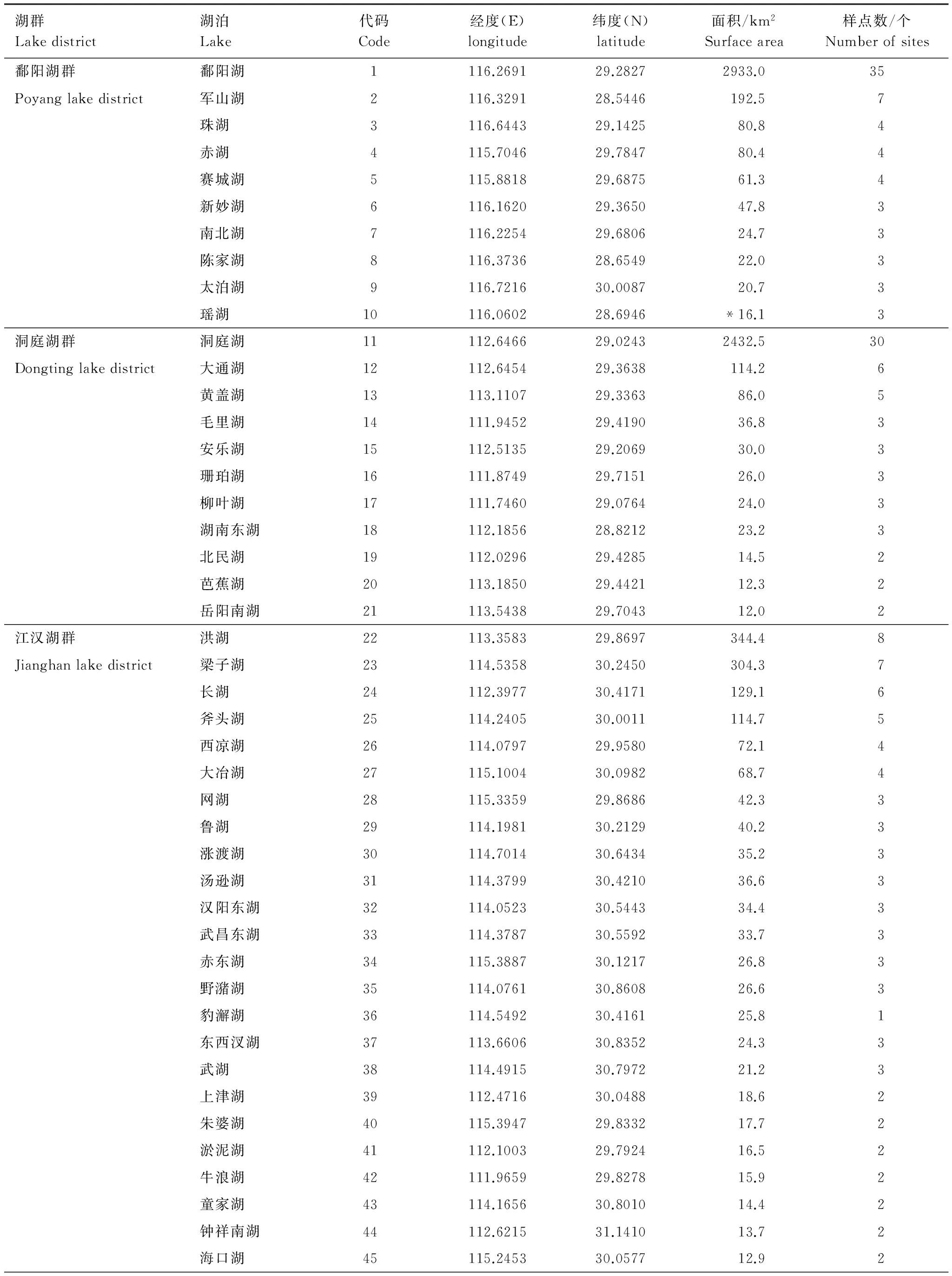
附錄1 研究湖泊經緯度、面積及采樣點個數
續表
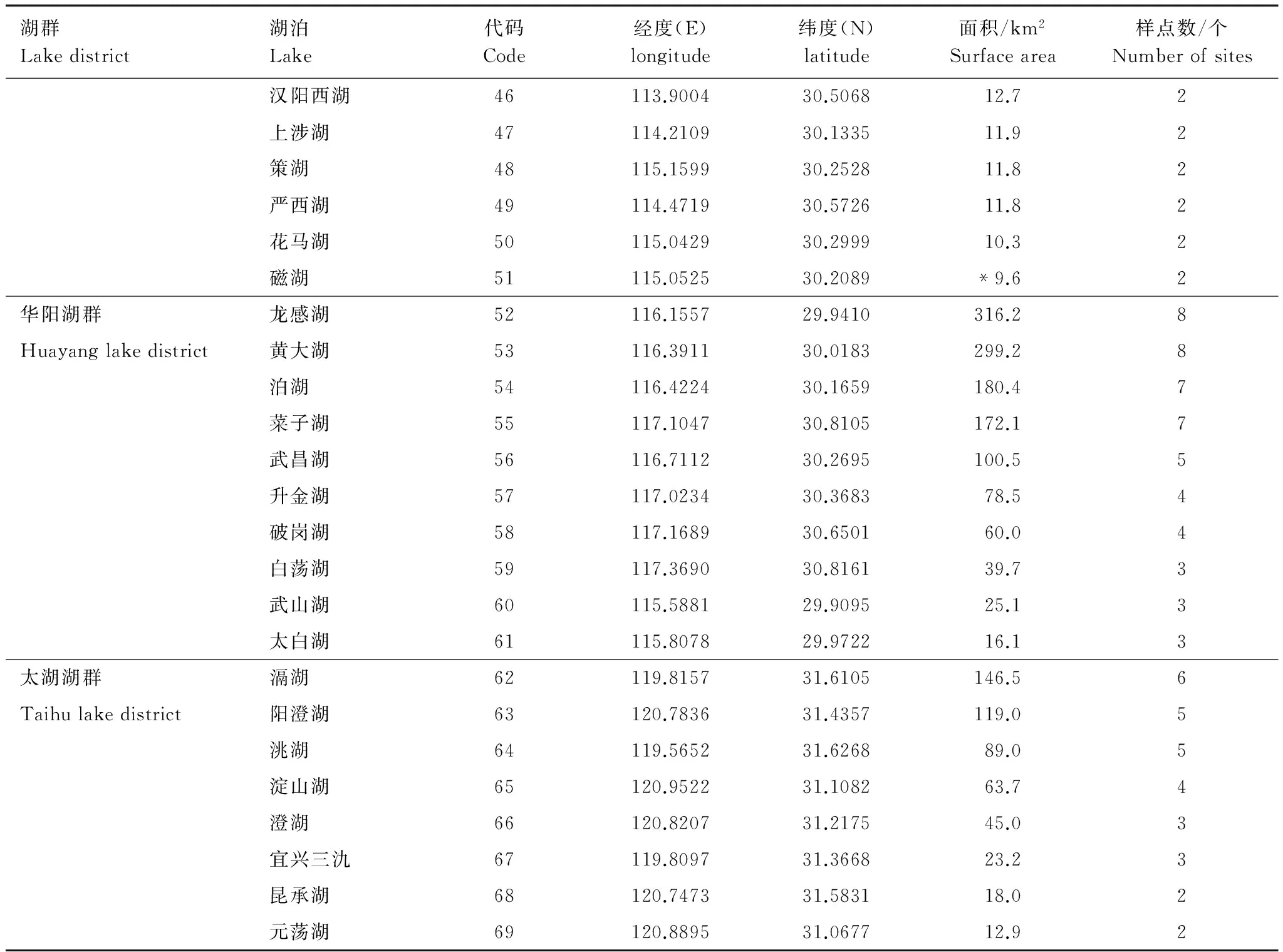
湖群Lakedistrict湖泊Lake代碼Code經度(E)longitude緯度(N)latitude面積/km2Surfacearea樣點數/個Numberofsites漢陽西湖46113.900430.506812.72上涉湖47114.210930.133511.92策湖48115.159930.252811.82嚴西湖49114.471930.572611.82花馬湖50115.042930.299910.32磁湖51115.052530.2089*9.62華陽湖群龍感湖52116.155729.9410316.28Huayanglakedistrict黃大湖53116.391130.0183299.28泊湖54116.422430.1659180.47菜子湖55117.104730.8105172.17武昌湖56116.711230.2695100.55升金湖57117.023430.368378.54破崗湖58117.168930.650160.04白蕩湖59117.369030.816139.73武山湖60115.588129.909525.13太白湖61115.807829.972216.13太湖湖群滆湖62119.815731.6105146.56Taihulakedistrict陽澄湖63120.783631.4357119.05洮湖64119.565231.626889.05淀山湖65120.952231.108263.74澄湖66120.820731.217545.03宜興三氿67119.809731.366823.23昆承湖68120.747331.583118.02元蕩湖69120.889531.067712.92
湖泊面積數據源自《中國湖泊志》[29],*瑤湖和磁湖在書中無記錄,面積數據源于Google earth面積測量工具,經緯度數據為用于CCA分析中構建空間變量的基礎數據
[1] Kratz T K, Frost T M. The ecological organisation of lake districts: general introduction. Freshwater Biology, 2000, 43: 297- 299.
[2] Downing J A, Prairie Y T, Cole J J, Duarte C M, Tranvik L J, Striegl R G, McDowell W H, Kortelainen P, Caraco N F, Melack J M. The global abundance and size distribution of lakes, ponds, and impoundments. Limnology and Oceanography, 2006, 51: 2388- 2397.
[3] Heiskary S A, Wilson C B, Larsen D P. Analysis of regional patterns in lake water quality: Using ecoregions for lake management in Minnesota. Lake and Reservoir Management, 1987, 3: 337- 344.
[4] Riera J N. A geomorphic template for the analysis of lake districts applied to the Northern Highland Lake District, Wisconsin, USA. Freshwater Biology, 2000, 43: 301- 318.
[5] Kratz T K, Webster K E, Bowser C J, Maguson J J, Benson B J. The influence of landscape position on lakes in northern Wisconsin. Freshwater Biology, 1997, 37: 209- 217.
[6] Fang J Y, Wang X P, Tang Z Y. Local and regional processes control species richness of plant communities: the species pool hypothesis. Biodiversity Science, 2009, 17: 605- 612.
[7] Huston M A. Local processes and regional patterns: appropriate scales for understanding variation in the diversity of plants and animals. Oikos, 1999. 86: 393- 401.
[8] Jiang J H, Dou H S, Su S D. Freshwater lake groups in the middle and lower reaches of the Yangtze and Huaihe Rivers. Wuhan: Changjiang Press, 2009: 428.
[9] Xie Z C. Studies on ecological characteristics of macrozoobenthos and lake ecosystem health in lake districts along the mid-lower reaches of Changjiang River [D]. Wuhan: Institute of Hydrobiology, Chinese Academy of Sciences, 2003.
[10] Liu Y Y, Zhang W Z, Wang Y X, Wang E Y. Economic fauna of China: Freshwater Mollusca. Beijing: Science Press, 1979: 134.
[11] Morse J C, Yang L F, Tian L X. Aquatic Insects of China Useful for Monitoring Water Quality, Nanjing: Hohai University Press, 1994: 570.
[12] Wang H Z. Studies on Taxonomy, Distribution and Ecology of Microdrile Oligochaetes of China, with descriptions of Two New Species from the Vicinity of the Great Wall Station of China, Antarctica. Beijing: Higher Education Press, 2002: 228.
[13] Jin X C, Tu Q Y. The standard methods for observation and analysis of lake eutrophication, 2nd ed. Beijing: China Environmental Science Press, 1990: 137.
[14] Cai Q H, Liu J K, King L. A comprehensive model for assessing lake eutrophication. Chinese Journal of Applied Ecology, 2002, 13: 1674- 1678.
[15] Clarke K R and Warwick R M. Changes in marine communities: an approach to statistical analysis and interpretation, 2nd edition. PRIMER-E, Plymouth, 2001.
[16] Hammer ?, Harper D A T, Ryan P D. PAST: paleontological statistics software package for education and data analysis. Palaeontologia electronica, 2001, 4: 1- 9.
[17] Borcard D, Legendre P, Drapeau P. Partialling out the spatial component of ecological variation. Ecology, 1992, 73: 1045- 1055.
[18] ter Braak C J F,milauer P. CANOCO reference manual and CanoDraw for Windows user′s guide: software for canonical community ordination (version 4.5): Microcomputer Power Ithaca, NewYork, 2002.
[19] Kumar S. Effect of Different Vegetation Systems on Soil Erosion and Soil Nutrients in Red Soil Region of Southeastern China. Pedosphere, 2003, 13: 121- 128.
[20] Anderson D M, Glibert P M, Burkholder J M. Harmful algal blooms and eutrophication: Nutrient sources, composition, and consequences. Estuaries and Coasts, 2002, 25: 704- 726.
[21] Br?nmark C. Interactions between epiphytes, macrophytes and freshwater snails: a review. Journal of Molluscan Studies, 1989, 55: 299- 311.
[22] Richards C, Johnson L B, Host G E. Landscape-scale influences on stream habitats and biota. Canadian Journal of Fisheries and Aquatic Sciences, 1996, 53: 295- 311.
[23] Heino J, Muotka T, Paavola R, Paasivirta L. Among-taxon congruence in biodiversity patterns: can stream insect diversity be predicted using single taxonomic groups? Canadian Journal of Fisheries and Aquatic Sciences, 2003, 60: 1039- 1049.
[24] Johnson R K, Goedkoop W, Sandin L. Spatial scale and ecological relationships between the macroinvertebrate communities of stony habitats of streams and lakes. Freshwater Biology, 2004, 49: 1179- 1194.
[25] Johnson R K and Goedkoop W. Littoral macroinvertebrate communities: spatial scale and ecological relationships. Freshwater Biology, 2002, 47: 1840- 1854.
[26] Sandin L and Johnson R K. Local, landscape and regional factors structuring benthic macroinvertebrate assemblages in Swedish streams. Landscape Ecology, 2004, 19: 501- 514.
[27] Donohue I, Jackson A L, Pusch M T, Irvine K. Nutrient enrichment homogenizes lake benthic assemblages at local and regional scales. Ecology, 2009, 90: 3470- 3477.
[28] ?kland R H. On the variation explained by ordination and constrained ordination axes. Journal of Vegetation Science, 1999, 10: 131- 136.
[29] Wang S M and Dou H S. Chinese Lakes. Beijing, Science Press, 1998: 480.
參考文獻:
[6] 方精云, 王襄平, 唐志堯. 局域和區域過程共同控制著群落的物種多樣性:種庫假說. 生物多樣性, 2009, 17: 605- 612.
[8] 姜加虎, 竇鴻身, 蘇守德主編. 江淮中下游淡水湖群. 武漢: 長江出版社, 2009, 428.
[9] 謝志才, 湖群大型底棲動物的生態學特征及生態系統健康評價. 武漢: 中國科學院水生生物研究所, 2003.
[10] 劉月英, 張文珍, 王躍先, 王恩義. 中國經濟動物志·淡水軟體動物. 北京: 科學出版社, 1979: 134.
[12] 王洪鑄. 中國小蚓類研究——附中國南極長城站附近地區兩新種.北京: 高等教育出版社, 2002: 228.
[13] 金相燦, 屠清瑛. 湖泊富營養化調查規范. 北京: 中國環境科學出版社, 1990: 317.
[29] 王蘇民和竇鴻身主編. 中國湖泊志. 北京: 科學出版社, 1998: 480.
StructureofmacrozoobenthosinlakesalongtheYangtzeRiverandrelationshipswithenvironmentalcharacteristics
CAI Yongjiu, JIANG Jiahu, ZHANG Lu, CHEN Yuwei, GONG Zhijun*
StateKeyLaboratoryofLakeScienceandEnvironment,NanjingInstituteofGeographyandLimnology,ChineseAcademyofSciences,Nanjing210008,China
In the middle and lower reaches of the Yangtze River, there are 108 freshwater lakes with surface area larger than 10 km2, accounting for 51.3% of the total freshwater lake area in China. However, few studies of macrozoobenthos were conducted at the regional scale, contrasting with many studies focused on individual or several lakes. To elucidate macrozoobenthic community structure and their relationships with spatial and environmental factors in shallow lakes along the Yangtze River, physicochemical parameters and macrozoobenthos were measured in 69 lakes of this region during summer in 2008 and 2009.We also evaluated the relative influences of local environmental conditions versus regional processes on macrozoobenthic community structure. The results showed that total dissolved solids (TDS), conductivity, nitrogen and phosphorus differed significantly among the five lake districts, but not for chemical oxygen demand (CODMn), chlorophyll a and Trophic State Index (TSI) index. In terms of density, 46 lakes (66.7% of the total 69 lakes) were dominated by Oligochaeta and Chironomidae, and Gastropoda was one of the dominant taxa in 16 lakes. With respect to biomass, 27 lakes were also dominated by Oligochaeta and Chironomidae, and 33 and 9 lakes were dominated by Gastropod and Bivlavia due to their relatively large body size, respectively. Canonical correspondence analyses (CCA) showed that macrozoobenthic community structure reflected the influence of local conditions (TSI, Macrophytes, Chl a, TDS) as well as regional processes. Only 33.9% of variation in a density matrix was explained by local environmental conditions and regional processes. Of the total explained variation, 48.1% and 35.4% were purely related to local environmental parameters and spatial variables, respectively. The high proportion of variation related to spatial variables indicated the importance of regional processes on spatial patterns of macrozoobenthic assemblages in this region.
the middle and lower reaches of the Yangtze River; lake district; shallow lakes; local and regional processes; macrozoobenthos
國家重點基礎研究發展計劃資助項目(2012CB417000);中國科學院南京地理與湖泊研究所一三五重點項目(NIGLAS2012135002);國家自然科學基金資助項目(41230744,31070418); 科技部基礎性工作專項資助項目(2006FY110600)
2012- 05- 15;
2012- 10- 26
*通訊作者Corresponding author.E-mail: zjgong @niglas.ac.cn
10.5846/stxb201205150721
蔡永久,姜加虎,張路,陳宇煒,龔志軍.長江中下游湖群大型底棲動物群落結構及影響因素.生態學報,2013,33(16):4985- 4999.
Cai Y J, Jiang J H, Zhang L, Chen Y W, Gong Z J.Structure of macrozoobenthos in lakes along the Yangtze River and relationships with environmental characteristics.Acta Ecologica Sinica,2013,33(16):4985- 4999.
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
中國生殖健康(2020年6期)2020-02-01 06:28:50
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
中國生殖健康(2019年11期)2019-01-07 01:28:02
汽車觀察(2018年10期)2018-11-06 07:05:26
中國科技博覽(2016年2期)2016-04-25 20:32:39
小學生導刊(2016年34期)2016-04-11 00:49:44
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
