高鹽下條斑紫菜光合特性和S-腺苷甲硫氨酸合成酶基因表達的變化
2013-12-20 05:59:42周向紅易樂飛徐軍田李信書閻斌倫
生態學報 2013年20期
周向紅,易樂飛,徐軍田,李信書,閻斌倫
(1.淮海工學院江蘇省海洋生物技術重點實驗室,連云港 222005;2.淮海工學院海洋學院,連云港 222005)
條斑紫菜(Porphyra yezoensis Ueda)是紅藻門中的一種大型海藻,作為重要的經濟藻類廣泛栽培于中國長江口以北、日本和韓國的近海區域,其2008年的產量已達5.5萬噸干重,產值達13億美元[1]。條斑紫菜不僅具有較高的食用和保健價值,還具有較高的學術研究價值。條斑紫菜葉狀體生長于潮間帶,由于潮汐變化其每天都會周期性地經歷干出與覆水過程,期間可能遭受到鹽度、溫度、水分、光照和紫外線變化引起的多種非生物脅迫[2],但是長期的演化使得條斑紫菜適應了這種潮間帶生活。因此條斑紫菜特殊的生活習性和脅迫適應機理引起了學者們的廣泛興趣。
紫菜耐受鹽度脅迫的機理是復雜的、多方面的,全面細致的機理仍然知之甚少[1],不過目前已有學者研究了紫菜在鹽度脅迫下的一些生理和生化變化過程,為闡明紫菜鹽度脅迫適應機理奠定了基礎。45—65鹽度的海水能誘導條斑紫菜積累滲透調節劑甘露醇,同時高鹽也誘導了脂質過氧化產物丙二醛的急劇增加,隨后丙二醛會被自由基清除系統清除[3]。在1/16×—1×海水范圍內,紫紅紫菜(P.purpurea)的滲透調節劑紅藻糖苷(floridoside)含量隨著鹽度的降低而降低,在1×—3×海水范圍內,紅藻糖苷含量隨著鹽度的增加而增加[4]。在9%以上的NaCl脅迫下4種紫菜的光化效率先急劇下降后緩慢下降,高鹽脅迫解除后光化效率先急劇上升后緩慢上升,最后恢復正常,其中2種高潮位紫菜(P.suborbiculata和P.pseudolineari s)的光化學效率比2種低潮位紫菜(P.seriata和P.yezoensi s)的恢復得快[5]。研究還發現了高鹽在光合系統的至少3個過程中影響紫菜(P.perforate)的光合作用[6]。
生物對逆境的適應與其生活習性密不可分,面對常態化的逆境,不同的生物采用不同的生存策略。仙人掌科植物采用特化的組織器官來適應常年的干旱環境,其根系發達,葉子退化成針狀,莖表覆以蠟質,從而減少水分蒸發和陽光照射。處于活動狀態的酵母、線蟲和緩步類動物會以一種稱之為隱生(cryptobiosis)的休眠機制來應對逆境,當面對干旱、低溫、高鹽和高壓時,它們的新陳代謝、生長、繁殖和衰老等活動均趨于減弱或暫時性停止,當環境條件一旦恢復后,將立即恢復到活動狀態[7]。例如干燥條件下,緩步類動物身體失水高達95%,身體收縮成桶狀,此時新陳代謝完全停止,機體依靠海藻糖和熱激蛋白等保護細胞基本結構免遭損害,隱生狀態下的緩步類動物能抵抗極端的冷、熱、旱、化學物質以及離子輻射[7-8]。
在紫菜中尚未發現有特化的組織器官來應對常態化的環境劇變[9],因此紫菜有可能采用了類似隱生的休眠機制來度過逆境,這樣它們能將能耗和損傷降低到最低限,這在理論上是有意義的。為此,本文擬從分子和生理兩個層面上來進行舉證。在生理水平上主要研究高鹽度脅迫對條斑紫菜光合放氧與呼吸耗氧的影響,在分子水平上主要研究高鹽度脅迫對抗逆基因S-腺苷甲硫氨酸合成酶(S-Adenosylmethionine synthetase,命名為PySAMS)表達的影響。
1 材料與方法
1.1 條斑紫菜葉狀體采集及脅迫處理
條斑紫菜葉狀體于2012年1月從連云港海區的紫菜養殖場采集,選取健康葉片并剪成2 cm×2 cm大小,暫養于30鹽度的過濾海水中,控制培養溫度為8℃、光照為100μmol photons·m-2·s-1、光周期為12L∶12D,24h通空氣,每天更換海水和ES培養基。暫養1周后將葉狀體分別轉移到30,40,50,60,70和80鹽度的海水中進行鹽度脅迫處理,處理于每日的10:00—14:00之間進行,每個處理3個重復,處理1h后取樣。
1.2 條斑紫菜葉狀體光合特性的檢測
采用氧電極法測定條斑紫菜葉狀體在鹽脅迫下的光合特性。參照鄒定輝等[10]的方法,用鹵鎢燈提供照明,通過調節鹵鎢燈距離將光照強度設定為400μmol photons·m-2·s-1;用低溫恒溫槽的循環水流將測定溫度控制在8℃。Oxygraph液相氧電極和配套軟件的使用參考儀器說明書進行,并且在正式測量之前先在8℃下標定氧電極。然后取鹽脅迫處理后的藻體小片塊約20mg加入到氧電極反應杯中,杯中反應介質為相應鹽度的滅菌海水,連續攪拌反應介質并記錄反應杯中氧的時間進程曲線。首先暗培養并紀錄耗氧數據,以此作為藻體的呼吸強度;然后再在光照下培養并記錄放氧數據,以此作為藻體的光合強度。
1.3 條斑紫菜葉狀體PySAMS基因表達的檢測
總RNA抽提參照TRIzol試劑(Invitrogen)說明書進行,抽提后的總RNA按DNase I(Fermentas)說明書進行處理,以避免殘留的微量基因組DNA影響定量PCR結果的準確性。使用核酸蛋白定量儀(Bio-Rad)檢測總RNA的純度及濃度,電泳檢測總RNA的完整性。
根據易樂飛等[11]克隆的 PySAMS基因序列(FJ404748)設計定量 PCR引物,正向引物是 5'AACGAGAGCGAGGACCTGATG 3',反向引物是5'CGTGCTGCGTGCTGATGAC 3',預期擴增產物長199bp。參考周向紅等[12]的方法,選用18SrRNA序列(DQ666486)作內參進行上樣誤差校正和標準化,其定量PCR的正向引物是5'TGCCAGCACTGCGTTCTTACC 3',反向引物是5'AGCCTTCCGACCCAGGACTATC 3',預期擴增產物長163bp。所有引物均由生工生物工程(上海)有限公司合成。
以Oligo(dT)和Random 6 mers為引物,使用PrimeScript? RT reagent Kit(TaKaRa)進行反轉錄反應。在IQ5 PCR儀(Bio-Rad)上進行實時熒光定量PCR反應。25μL的反應體系中包含12.5μL 2×SYBR? Premix Ex TaqTMII(TaKaRa)、0.2μmol/L每種引物和2μL反轉錄產物。采用兩步PCR法進行擴增,即首先95℃預變性1min;然后進入45個循環,每個循環中95℃變性10s,62℃延伸30s;循環結束后,從55℃緩慢升溫到95℃,制備熔解曲線。同時還以10×系列稀釋的反轉錄產物為模板進行定量PCR,以制作PySAMS和內參的標準曲線。每次反應都設置陰性對照和無模板對照,每個反應設3個復孔。
1.4 數據分析
應用PFAFFL[13]建立的數學模型對定量PCR數據進行處理,從而獲得PySAMS在不同鹽度下的相對表達量;藻體放氧和耗氧速率以每克鮮葉片每小時產生和消耗的O2的量為單位。所有數據采用SPSS進行統計分析,并以均數±標準差表示。因為各組數據都通過了正態性和方差齊性檢驗,所以組間差異采用單因素方差分析(one-way ANOVA),組間多重比較采用Ducan檢驗,P<0.05表示差異顯著。
2 結果與分析
2.1 條斑紫菜葉狀體光合特性
將藻體從30鹽度轉移至40鹽度的海水中培養1h后,光合放氧率從197.3μmolO2·g-1鮮重·h-1下降至182.2μmolO2·g-1鮮重·h-1;雖然40鹽度下的放氧率稍有降低,但是與30鹽度的無顯著差異。更高鹽度(50—80鹽度)的海水顯著性地抑制了藻體光合作用,光合放氧率下降至149.7—118.2μmolO2·g-1鮮重·h-1范圍,而且鹽度越高,光合放氧率越低(圖1)。高鹽度海水培養的藻體呼吸強度的變化趨勢類似于光合作用的變化。在40鹽度海水條件下,呼吸耗氧率與30鹽度的幾乎沒有變化,維持在22.6μmolO2·g-1鮮重·h-1左右。更高鹽度(50—80鹽度)的海水則顯著性地抑制了藻體呼吸作用,呼吸耗氧率下降至15.1—12.1μmolO2·g-1鮮重·h-1范圍,而且鹽度越高,呼吸耗氧率越低,但彼此間尚無顯著差異(圖1)。
高鹽度海水培養不同程度地抑制了藻體的光合和呼吸作用。放氧率降低表明藻體的有機物合成下降,耗氧率降低表明藻體的生理活性下降,因此50—80鹽度不利于藻體生長,而且鹽度越大則影響越大。
2.2 條斑紫菜葉狀體PySAMS基因表達
核酸分離純化是各類分子操作的第一步,因此總RNA質量對后續反轉錄與PCR反應具有較大影響。本實驗中,抽提的所有總 RNA 經檢測后顯示 A260/A280的比值介于 1.9—2.1,A260/A230的比值介于 2.0—2.3,而且經普通瓊脂糖凝膠電泳后顯示出明亮且清晰的28S和18SrRNA條帶以及一些細胞器rRNA條帶。上述結果表明總RNA中沒有明顯的蛋白質、多糖或多酚等殘留,總RNA片段完整且沒有明顯降解,進一步的DNase I處理清除了總RNA中殘留的DNA。因此總RNA質量可以滿足后續實驗要求。

圖1 條斑紫菜葉狀體在不同鹽度海水培養1h后的光合特性Fig.1 Photosynthetic characteristics of P.yezoensis thalli under salinity stress
PySAMS基因的擴增曲線呈規則的“S”形,基線平整、指數區明顯;熔解曲線僅顯示單一的特異峰;標準曲線的斜率為3.45,擴增效率為95%,相關系數是0.998。陰性對照和無模板對照的擴增曲線均呈水平直線狀,復孔間擴增曲線基本重疊。這些說明擴增產物特異性高,無引物二聚體及非特異性擴增,確保了定量的準確性。
定量PCR結果顯示高鹽度的海水對PySAMS基因的相對表達量具有顯著影響(圖2)。將條斑紫菜葉狀體轉移至40鹽度海水中培養,1h后檢測到PySAMS被誘導表達,該表達量是30鹽度下的1.58倍,且與之差異顯著。50鹽度海水培養1h后也檢測到PySAMS表達量升高,該表達量是30鹽度下的1.43倍。更高鹽度(60—80鹽度)海水脅迫后,PySAMS的表達量不但沒有升高反而不同程度的下降,是30鹽度下的0.70—0.73倍,且與之差異顯著。
3 討論
光合與呼吸作用是植物體內非常重要的生理過程,但容易受到外界環境的影響。研究表明葉綠體的類囊體膜是光能吸收、傳遞和轉換的結構基礎;鹽脅迫下,過量鹽離子積累會使類囊體膜糖脂含量顯著下降,不飽和脂肪酸含量降低,飽和脂肪酸含量升高,垛疊狀態的類囊體膜的比例減少,類囊體排列紊亂、膨大,基粒排列方向改變,基粒和基質片層界限模糊不清,葉綠素含量降低,從而降低了植物對光量子的有效吸收、傳遞和利用,使激發能不能迅速傳遞和轉化,降低了光能轉化為生物化學能的效率,最終降低了光合能力[14]。本文研究結果也顯示了類似的抑制效應,即鹽度越高對藻體光合作用的抑制越強(圖1)。
呼吸作用為植物的生命活動提供大部分能量,當然也為植物抵御鹽脅迫提供能量;植物積累或拒絕鹽離子以及合成有機滲透調節物來適應或抵抗鹽脅迫等一系列過程都需要消耗大量能量,因此低濃度鹽脅迫下或鹽脅迫早期植物呼吸強度通常會增加,然而更高強度的鹽脅迫會抑制呼吸[14]。本文研究結果與高等植物的略有不同。條斑紫菜體細胞能正常生長發育的鹽度范圍在19.6—32.8之間[15],因此40鹽度已屬輕度鹽脅迫。雖然鹽度越高對藻體呼吸作用的抑制越強,但是輕度鹽脅迫(40鹽度)并沒有增加藻體的呼吸強度(圖1B)。類似現象在其它耐鹽或耐旱的藻類中也有發現。例如,培養液中NaCl濃度的升高引起發狀念珠藻(Nostoc flagelliforme,俗稱“發菜”)藻體呼吸作用的降低,在 0—0.1mol/L NaCl條件下,呼吸維持在 3.57—3.60μmol·g-1·h-1,一旦鹽度增加到0.2mol/L NaCl,呼吸迅速下降至2.44μmol·g-1·h-1,而且鹽度越高呼吸作用越低[16]。將 1.5mol/L NaCl培養的杜氏鹽藻(Dunaliella salina)轉移至 2.5mol/L NaCl中培養 20min,呼吸作用也顯著降低[17]。
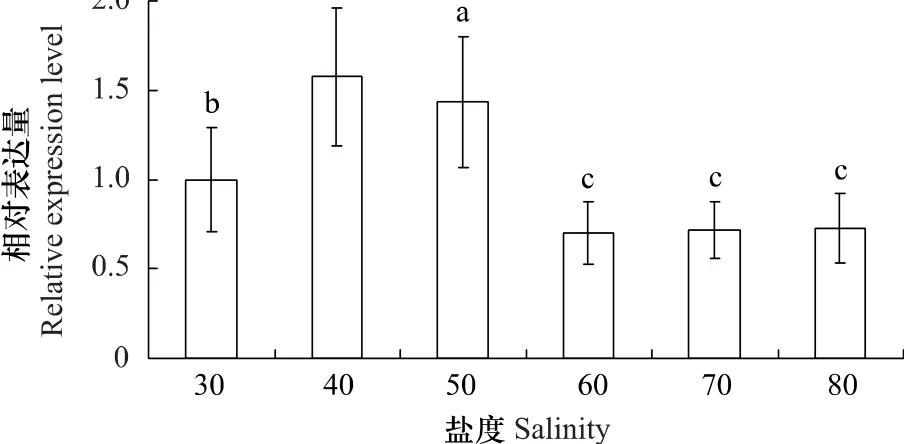
圖2 條斑紫菜葉狀體PySAMS基因在鹽度脅迫下的相對表達量Fig.2 The relative mRNA expression levels of PySAMS gene in P.yezoensis thalli under salinity stress
SAMS在生物體內催化ATP和L-甲硫氨酸反應生成S-腺苷甲硫氨酸(S-adenosylmethionine,SAM),SAM作為重要的中間代謝物參與轉甲基、轉硫和轉氨丙基等反應[18],反應能生成乙烯、多胺、還原型谷胱甘肽和甜菜堿等分子[11],這些物質能參與植物對逆境的抵抗過程,因此SAMS在植物的抗逆過程中發揮著積極作用。大量數據表明SAMS基因的確受環境脅迫的誘導表達[19-21],而且超表達SAMS還能提高轉基因植物的抗逆能力[22-23]。與陸地棉(Gossypium hirsutum)[24]、裙帶菜(Undaria pinnatifida)[25]等這些陸生植物和海洋藻類的SAMS一樣,PySAMS受到鹽脅迫的誘導表達。但是,PySAMS又有著其特殊的表達模式,即在40和50鹽度時PySAMS誘導表達明顯,在更高鹽度(60、70和80鹽度)脅迫下不被誘導(圖2)。
綜合生理和分子水平上的數據來看,30至40鹽度范圍內條斑紫菜光合與呼吸作用均不受影響,那么在40鹽度下被誘導表達的PySAMS則可能參與維持了細胞內環境穩定,進而維持光合與呼吸作用的正常進行。在50鹽度下,雖然PySAMS被誘導表達,但是PySAMS等抗逆基因以及滲透調節物質可能不足以維持內環境的穩定,于是光合與呼吸作用受到了鹽脅迫的抑制。更高鹽度(60至80鹽度)下,藻體的光合與呼吸作用以及PySAMS基因表達都受到了明顯抑制;光合作用的減弱將直接影響到藻體生長,呼吸作用的減弱將直接減少細胞內能量的產生,PySAMS表達量的減少將消弱維持細胞內環境穩定的能力。這些數據驗證了我們的推斷,即在面對逆境時條斑紫菜葉狀體可能采用隱生策略來逐步降低體內新陳代謝以度過不良環境。這一假設也得到了其它數據的支持,例如隨著脫水的增加,條斑紫菜葉狀體光合和呼吸都逐步減弱,最后幾近暫停[26],抗逆基因HSP70和HSP90的表達也受抑制[27-28]。雖然隱生的具體機理不詳,但一般認為高濃度海藻糖可代替丟失的水,在細胞內形成氫鍵框架以抵御脅迫,熱激蛋白則在脅迫解除后的恢復過程中發揮作用[29]。紅藻含有高濃度的可溶性糖——紅藻糖苷(floridoside),其在細胞質內也可以形成氫鍵框架[9]。因此這些數據暗示著條斑紫菜葉狀體能采用隱生策略來度過不良環境。
為什么高鹽脅迫抑制了PySAMS基因表達呢?對于條斑紫菜葉狀體,高鹽不僅抑制了PySAMS表達,還抑制了滲透調節物質甘露醇的合成,45鹽度脅迫后的12h內藻體內甘露醇含量遠遠高于同期55和65鹽度脅迫下的甘露醇含量[3]。雖然上述現象容易讓人認為PySAMS和甘露醇不是藻體耐鹽的主要物質,但是我們更傾向于認為:在細胞內存在某種胞質濃度感應裝置,當胞質濃度增加并超過某一閾值之后細胞將降低體內新陳代謝以度過不良環境;那么在輕度鹽脅迫時,胞質濃度達到閾值所經歷時間長或者達不到閾值,此期間有更多的機會合成抗逆物質,反之,在重度鹽脅迫時,胞質濃度達到閾值所經歷時間短,此期間可能來不及合成抗逆物質。這一假設不僅能解釋鹽脅迫下的這些特殊現象,甚至還能對干出脅迫進行部分解釋。條斑紫菜葉狀體HSP70和HSP90在失水脅迫后不被誘導表達的原因可能是藻體一旦干出后,水分會從細胞內迅速流失,胞質濃度快速達到閾值,于是沒有足夠的時間去合成這些基因。
條斑紫菜耐受鹽度脅迫的機理是復雜的、多方面的,仍然有許多機理不明[1],雖然本文提出了條斑紫菜的隱生假說,但是藻體如何感知胞質濃度變化,如何在高鹽環境中生存以及如何在鹽脅迫解除后迅速恢復生長等,詳細的機理還有待于進一步研究。
[1]Blouin N A,Brodie JA,Grossman A C,Xu P,Brawley SH.Porphyra:a marine crop shaped by stress.Trends in Plant Science,2011,16(1):29-37.
[2]Davison I R,Pearson G A.Stress tolerance in intertidal seaweeds.Journal of Phycology,1996,32(2):197-211.
[3]Feng C,Lu X Z,Yu W G.Biochemical and physiological effects of adversity stress on Porphyra yezoensis.Transactions of Oceanology and Limnology,2004,26(3):22-26.
[4]Reed R H,Collins J C,Russell G.The effects of salinity upon galactosyl-glycerol content and concentration of the marine red alga Porphyra purpurea(Roth)C.Ag.Journal of experimental botany,1980,31(6):1539-1554.
[5]Hong D D,Kim T H,Hwang M S,Chung I K,Lee C H.Effects of salinity on chlorophyll fluorescence from Porphyra thalli and comparison of species with different intertidal distribution.Journal of Fisheries Science and Technology,1998,1(1):122-128.
[6]Satoh K,Smith C M,Fork D C.Effects of salinity on primary processes of photosynthesis in the red alga Porphyra perforata.Plant Physiology,1983,73(3):643-647.
[7]Schill R O,Mali B,Dandekar T,Schn?lzer M,Reuter D,Frohme M.Molecular mechanisms of tolerance in tardigrades:New perspectives for preservation and stabilization of biological material.Biotechnology Advances,2009,27(4):348-352.
[8]Su L N,Li X C.Overview on dormancy of tardigrada.Sichuan Journal of Zoology,2006,25(1):191-195.
[9]Liu Y-C.Mechanism for differential desiccation tolerance in Porphyra species[D].Boston:Northeastern University,Department of Biology,2009.
[10]Zou D H,Gao K S.The effects of elevated inorganic carbon concentration on photosynthesis of Ulva lactuca under different temperature.Marine Sciences,2002,26(3):49-52.
[11]Yi L F,Wang P,Zhou X H,Liu C W.cDNA cloning and bioinformatic analysis of SAMS gene from Porphyra yezoensis.China Biotechnology,2009,29(7):43-49.
[12]Zhou X H,Yi L F,Li X S,Wang P,Yang P L.Molecular cloning and expression pattern of glutathione S-transferase gene in Porphyra yezoensis Ueda(Bangiales,Rhodophyta).Journal of Fisheries of China,2011,35(9):1354-1361.
[13]Pfaffl M W.A new mathematical model for relative quantification in real-time RT-PCR.Nucleic acids research,2001,29(9):e45.
[14]Yang X H,Jiang W J,Wei M,Yu H J.Review on plant response and resistance mechanism to salt stress.Journal of Shandong Agricultural University(Natural Science),2006,37(2):302-305,308.
[15]Yan X H,Jiang H B.Effects of salinity on the growth and development of somatic cells and preliminary observation of lower salinity Resistant in Porphyra yezoensis.Journal of Shanghai Fisheries University,1993,2(1):34-40.
[16]Bi Y H,Deng Z Y,Hu Z Y,Xu M.Response of Nostoc flagelliforme to salt stress.Acta Hydrobiologica Sinica,2005,29(2):125-129.
[17]Liu X D,Shen Y G.Salt shock induces state IItransition of the photosynthetic apparatus in dark-adapted Dunaliella salina cells.Environmental and Experimental Botany,2006,57(1/2):19-24.
[18]Zhang J G,Li X H,Yuan Z Y.Research advances in gene and structure of S-adenodylL-methionine synthetase.Industrial Microbiology,2005,35(3):39-44.
[19]Ma X,Wang Z,Qi Y,Zhao Y,Zhang H.Isolation of S-adenosylmethionine synthetase gene from Suaeda salsa and its differential expression under NaCl stress.Acta Botanica Sinica,2003,45(11):1359-1365.
[20]Lin F Y,Wang SQ,Hu Y G,He B R.Cloning of a S-adenosylmethionine synthetase gene from broomcorn millet(Panicum miliaceum L.)and its expression during drought and re-watering.Acta Agronomica Sinica,2008,34(5):777-782.
[21]Hwang SS,Cheah SC,Kulaveerasingam H,Tan S H.Molecular Cloning and Characterization of S-adenosylmethionine Synthetase Isolated from Suspension Culture Cdna Library of Oil Palm(Elaeis guineensis Jacq.).Pakistan Journal of Biological Sciences,2003,6(16):1468-1475.
[22]Fan JP,Bai X,Li Y,Ji W,Wang X,Cai H,Zhu Y M.Cloning and function analysis of gene SAMSfrom Glycine soja.Acta Agronomica Sinica,2008,34(9):1581-1587.
[23]Hua Y,Zhang B X,Cai H,Li Y,Bai X,Ji W,Wang Z Y,Zhu Y M.Stress-inducible expression of GsSAMS2 enhances salt tolerance in transgenic Medicago sativa.African Journal of Biotechnology,2012,11(17):4030-4038.
[24]Zhou K,Song L Y,Ye W W,Wang J J,Wang D L,Fan B X.Cloning and expression of GhSAMS gene related to salt-tolerance in Gossypium hirsutum L.Acta Agronomica Sinica,2011,37(6):1012-1019.
[25]Qiao K.Cloning,analysis and prokaryotic expression of SAMSgene in Undaria pinnatifida[D].Dalian:Liaoning Normal University,2011.
[26]Gao K,Aruga Y.Preliminary studies on the photsynthesis and respiration of Porphyra yezoensis under emersed conditions.Journal of the Tokyo University of Fisheries,1987,47(1):51-65.
[27]Zhou X,Wang P,Yan B,Li X,Yi L.Characterization and expression patterns of two 70-kDa heat shock protein genes in the intertidal red alga Porphyra yezoensis.Botanica Marina,2011,54(5):447-455.
[28]Zhou X H,Li X S,Wang P,Yan B L,Teng Y J,Yi L F.Molecular cloning and expression analysis of HSP90 gene from Porphyra yezoensis Udea(Bangiales,Rhodophyta).Journal of Fisheries of China,2010,34(12):1844-1852.
[29]Welnicz W,Grohme M A,Kaczmarek L,Schill R O,Frohme M.Anhydrobiosis in tardigrades the last decade.Journal of insect physiology,2011,57(5):577-583.
參考文獻:
[3]馮琛,路新枝,于文功.逆境脅迫對條斑紫菜生理生化指標的影響.海洋湖沼通報,2004,26(3):22-26.
[8]蘇麗娜,李曉晨.緩步動物休眠現象研究進展.四川動物,2006,25(1):191-195.
[10]鄒定輝,高坤山.不同溫度條件下無機碳濃度增高對石莼光合作用的影響.海洋科學,2002,26(3):49-52.
[11]易樂飛,王萍,周向紅,劉楚吾.條斑紫菜SAMS基因克隆與生物信息學分析.中國生物工程雜志,2009,29(7):43-49.
[12]周向紅,易樂飛,李信書,王萍,閻斌倫.條斑紫菜谷胱甘肽S-轉移酶基因的克隆與表達分析.水產學報,2011,35(9):1354-1361.
[14]楊曉慧,蔣衛杰,魏珉,余宏軍.植物對鹽脅迫的反應及其抗鹽機理研究進展.山東農業大學學報(自然科學版),2006,37(2):302-305+308.
[15]嚴興洪,江海波.鹽度對條斑紫菜體細胞生長發育的影響及耐低鹽應變異體的初步觀察.上海水產大學學報,1993,2(1):34-40.
[16]畢永紅,鄧中洋,胡征宇,徐敏.發狀念珠藻對鹽脅迫的響應.水生生物學報,2005,29(2):125-129.
[18]張建國,李新華,袁中一.腺苷甲硫氨酸合成酶的基因及結構研究進展.工業微生物,2005,35(3):39-44.
[20]林凡云,王士強,胡銀崗,何蓓如.糜子SAMS基因的克隆及其在干旱復水中的表達模式分析.作物學報,2008,34(5):777-782.
[22]樊金萍,柏錫,李勇,紀巍,王希,才華,朱延明.野生大豆S-腺苷甲硫氨酸合成酶基因的克隆及功能分析.作物學報,2008,34(9):1581-1587.
[24]周凱,宋麗艷,葉武威,王俊娟,王德龍,樊保香.陸地棉耐鹽相關基因GhSAMS的克隆及表達.大連:作物學報,2011,37(6):1012-1019.
[25]喬坤.裙帶菜SAMS基因的克隆、分析及原核表達[D].大連:遼寧師范大學,2011.
[28]周向紅,李信書,王萍,閻斌倫,滕亞娟,易樂飛.條斑紫菜HSP90基因的克隆與表達分析.水產學報,2010,34(12):1844-1852.
