玉米種子老化過程中抗氧化酶活性的變化
2013-12-23 06:25:10劉建軍馬俊華孟俊文王自霞周小梅
山西農業科學 2013年9期
劉建軍,馬俊華,孟俊文,王自霞,周小梅
(1.山西省農業科學院,山西太原030006;2.山西大學生命科學學院,山西太原030006)
玉米是種植最廣泛的谷類作物之一。全世界有100多個國家種植玉米,玉米在世界谷物總產量中僅次于小麥,位居第2。我國玉米常年種植面積近2 400萬hm2,總產量約1.28億t,占糧食總產量的1/4左右。玉米產量的高低,直接影響到我國的糧食生產總量[1]。目前,生產上所用的玉米良種、玉米自交系以及玉米種質資源因保存不妥或存放時間過長引起的種子老化或劣變會造成種子活力喪失及發芽率降低等不可逆轉的后果,給農業生產帶來嚴重的經濟損失[2]。植物體內超氧化物歧化酶(SOD)、過氧化物酶(POD)、過氧化氫酶(CAT)、抗壞血酸專一性過氧化物酶(APX)等是植物體內活性氧的清除劑,稱之為抗氧化酶類,它們共同作用可使細胞內的活性氧維持在正常水平,從而防止活性氧對植物細胞造成的傷害[3-4]。但種子老化會使植物體內活性氧的產生和清除失衡,造成細胞內氧自由基的積累,導致種子失去活力。
本研究選用2個玉米自交系進行玉米種子老化過程中抗氧化酶的變化研究,以探討種子老化與抗氧化酶活性的相互關系,為種子的安全保存提供一定理論依據。
1 材料和方法
1.1 試驗材料
供試材料為玉米自交系478和Mo17,由山西豐禾種業2010年在壽陽基地隔離種植收獲。其中,478種子水分含量為11.04%,百粒質量為28.7 g;Mo17種子水分含量為10.51%,百粒質量為25.36 g。
1.2 試驗方法
1.2.1(58±1)℃熱水人工加速老化法(HW) 將種子置于(58±1)℃恒溫水浴中處理5~40 min(處理間隔為5 min)(同時以未處理為對照),取出自然晾干,待種子含水量降至初始狀態后進行試驗。
1.2.2 酶液制備 參照李忠光等[5]的方法,隨機選取30粒玉米種子,在溫度為26℃的發芽箱中進行萌發,3次重復,取不同萌發時間(0,8,12,16,24,32,36,40,48 h)的種胚,按照種胚的質量∶提取液為1∶5的比例(緩沖液為50 mmol/L,pH值7.0的Tris-HCl)冰浴研磨提取。提取液于2~4℃條件下10 000 r/min離心20 min,取上清液在0~4℃下保存備用。
1.2.3 酶活性的測定 其參照李忠光等[5-6]的方法進行測定。其中,SOD活性采用NBT光還原法測定;POD活性采用愈創木酚顯色法測定;CAT活性采用紫外吸收法測定;APX活性采用紫外吸收法測定。
1.3 數據分析
采用SPSS 13.0進行數據統計,采用Student-Newman-Keuls提出的q 測驗進行顯著性分析(大寫英文字母表示1%顯著水平),使用Origin 7.5軟件作圖。數據均用平均值±標準差表示。
2 結果與分析
對玉米自交系種子478,Mo17進行不同時間的(58±1)℃人工熱水老化處理,SOD,POD,CAT和APX活性的變化如表1,2所示。由表1,2可知,老化處理后,SOD活性隨老化時間的延長整體上呈遞增趨勢,POD,CAT和APX活性整體上呈下降趨勢,2個玉米種子的變化趨勢基本一致,抗氧化酶活性變化均達到顯著水平。
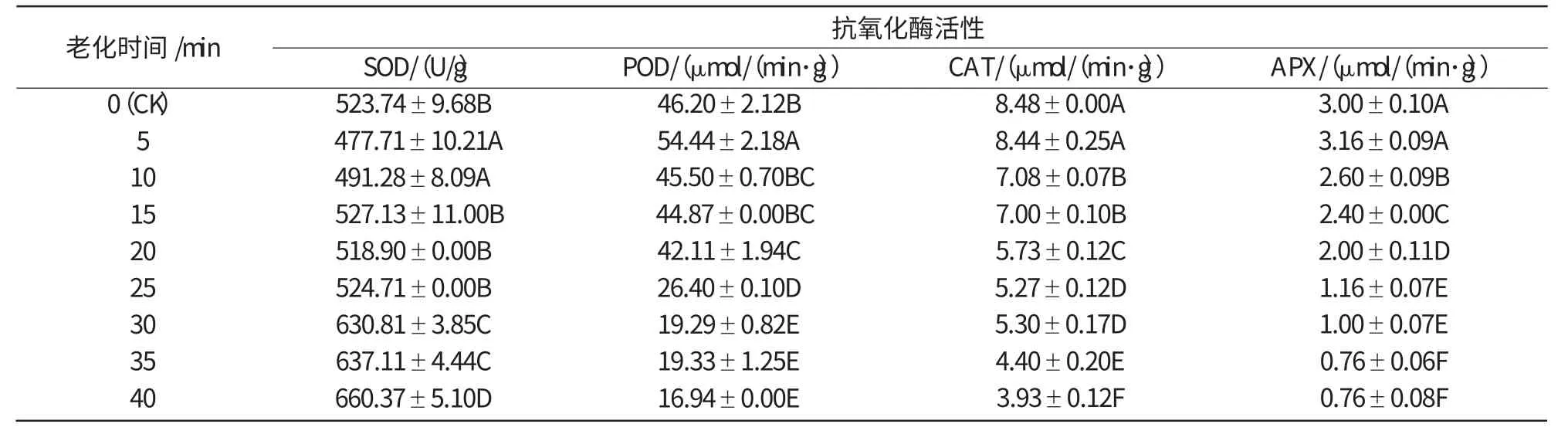
表1 (58±1)℃熱水老化對玉米478 種子抗氧化酶活性的影響
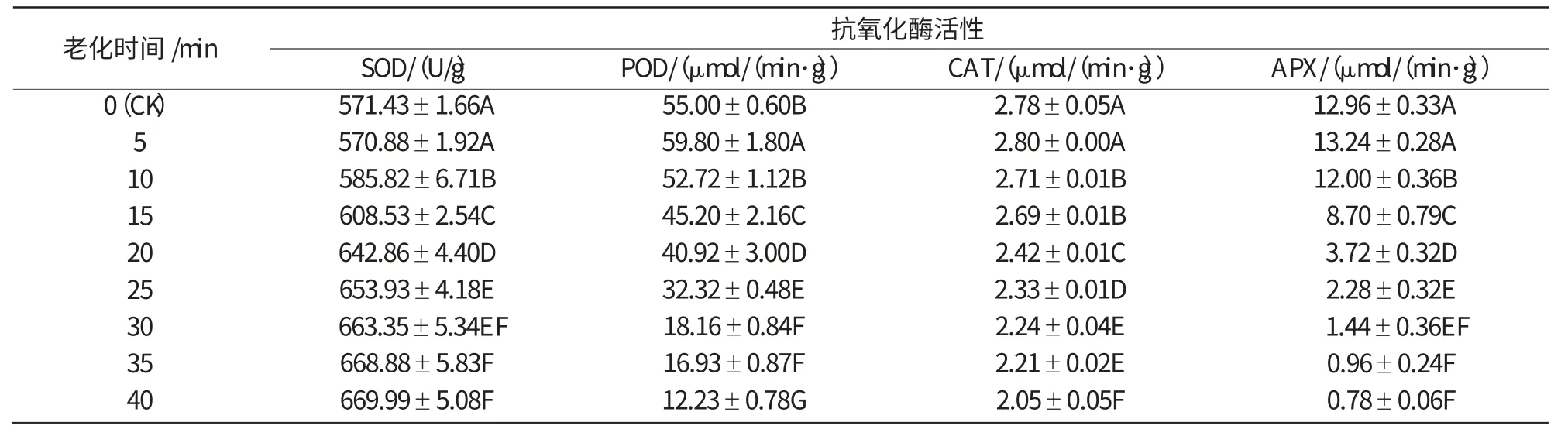
表2 (58±1)℃熱水老化對玉米Mo17 種子抗氧化酶活性的影響
2.1 玉米種子老化過程中SOD 活性的變化
由圖1可知,478玉米種子SOD活性隨老化時間的延長呈緩慢升高的趨勢。在0~25 min的老化時間段,SOD活性保持基本不變,僅老化5~10 min有明顯下降,這可能和短時間的熱水處理使發芽率提高有一定的關系。萌發初期,種子中SOD活性是隨種子萌發代謝的加強而下降,隨種子老化活力降低而升高。這可能是由于SOD在干種子內就有儲存,所以在種子萌發開始,新的SOD還沒有合成,但細胞代謝的加強,消耗了原有的SOD而導致該酶活性的下降。老化25 min后,種子的SOD活性迅速升高,與對照之間存在極顯著差異,表明了此時種子的活力迅速下降。有研究也證明,此時老化玉米種子的發芽勢和相對活力指數都降到50%以下[7]。說明種子老化到一定程度超出了細胞所能忍受的范圍時,其細胞自身的抗性調節能力開始下降。

由圖2可知,老化玉米Mo17種子SOD活性的變化趨勢與478相似,活性變化曲線緩慢升高。從老化5 min開始,SOD活性一直平穩升高,至20 min以后活性變化不大,并處于一個較高的水平。
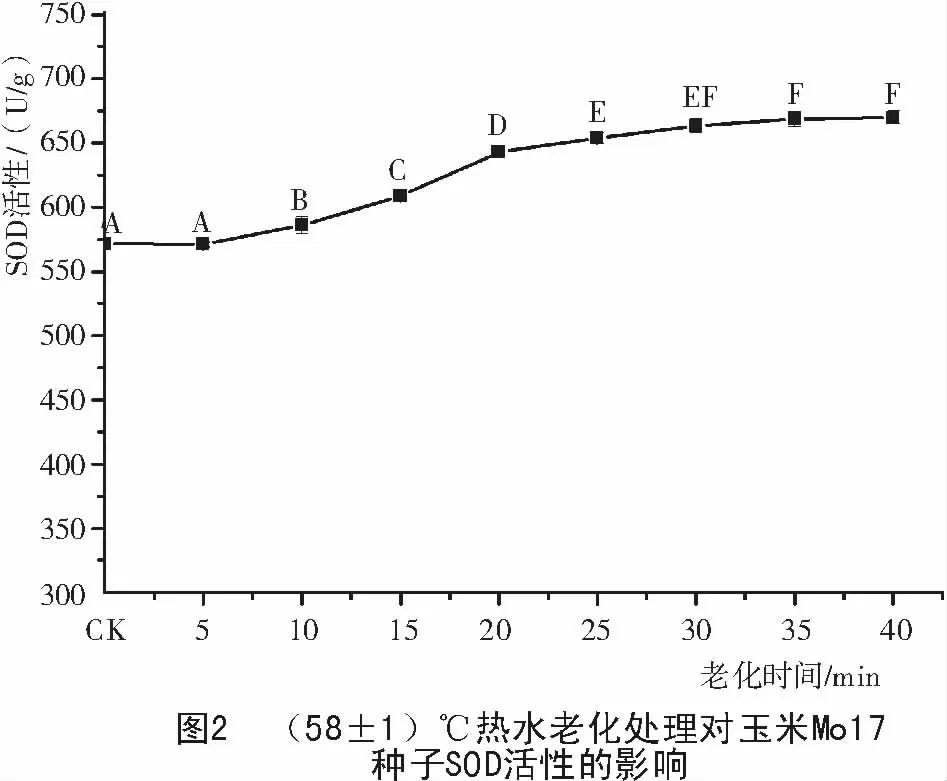
從2種玉米種子老化過程中SOD的升降變化幅度可以看出,478種子比Mo17種子耐老化性強,而且種子活力較高,說明不同品種自身的抗氧化調節能力不同,種子活力不同。
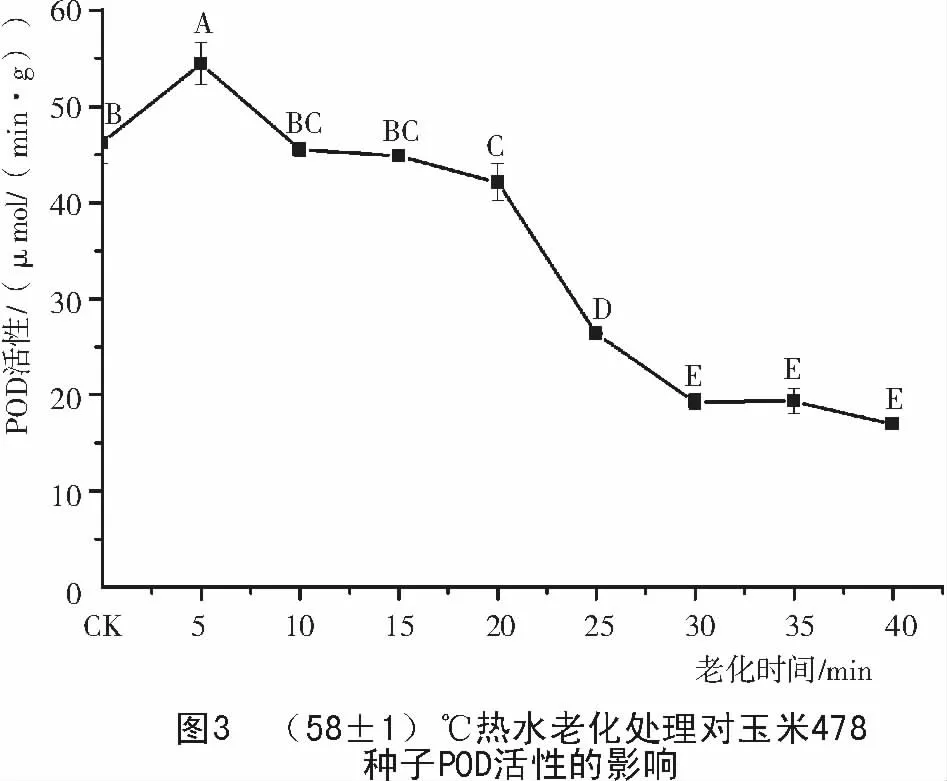
2.2 玉米種子老化過程中POD 活性的變化
由圖3可知,玉米478種子POD活性隨老化處理時間的延長而下降,在老化處理20 min內,POD活性變化不大,只在5 min時,比對照有所升高,這是由于短時的熱水處理刺激種子本身所做出的應急反應;在20~30 min期間,POD活性迅速下降;30 min之后,POD活性降到一個較低的水平,基本保持不變。老化20 min的478玉米種子發芽率仍在80%左右[7],此時的POD活性還較高,隨后POD活性急劇下降,到30 min后發芽率降至50%左右[7],種子活力與POD活性的密切相關,表明了POD活性可作為一個檢測指標顯示種子活力的高低及喪失的特性。
由圖4可知,老化的玉米Mo17種子POD活性的變化趨勢類似于478,不同的是Mo17種子POD活性從老化的開始就一直平穩下降,而478種子的POD活性變化則是慢—快—慢。說明隨著老化加劇,細胞中POD活性清除過氧化物的能力降低,進一步對細胞造成嚴重損傷,導致種子活力發生不可逆轉的下降,直到死亡。
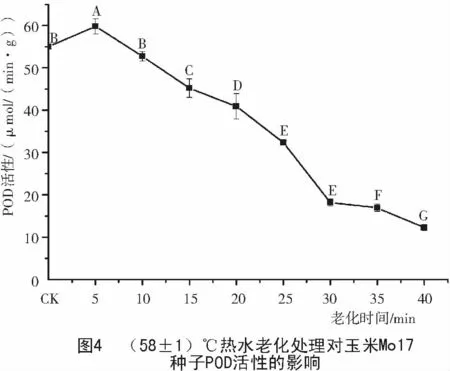
2.3 玉米種子老化過程中CAT 活性的變化
由圖5,6可知,老化處理后的玉米478和Mo17種子,CAT活性都隨老化時間的延長而緩慢下降,但2個玉米種子對照的CAT活性差別較大,Mo17對照的CAT活性只有478對照的1/3。478種子老化5~40 min,CAT酶活性下降了54%,而Mo17只下降了27%。說明老化可使種子中CAT活性降低,清除過氧化氫的能力減弱,種子活力下降,不同品種耐老化能力不同。
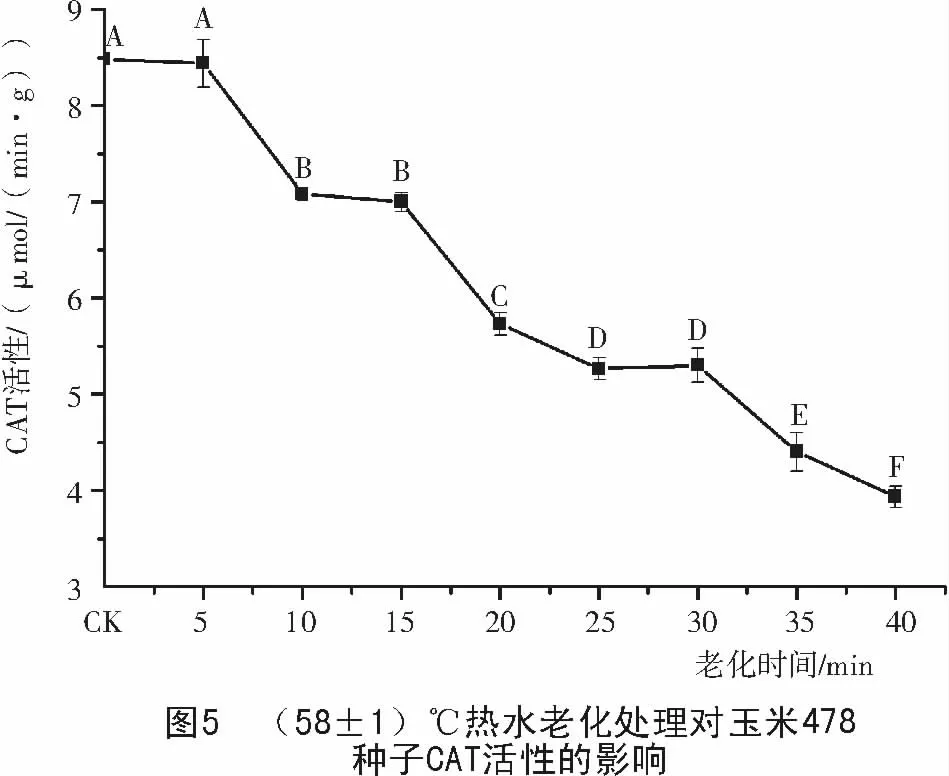
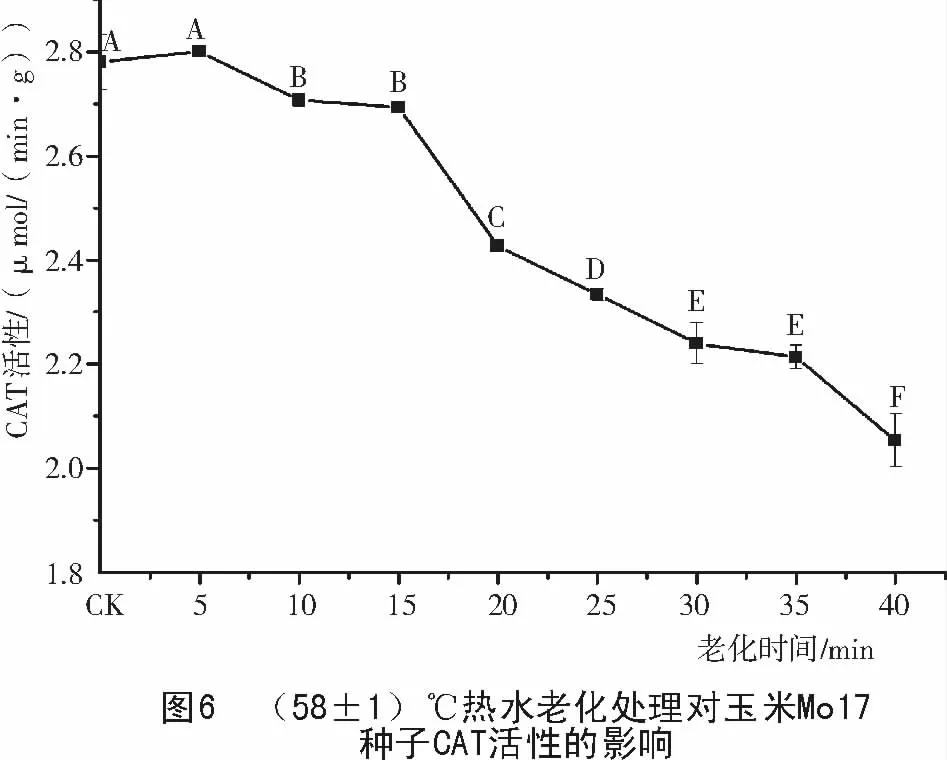
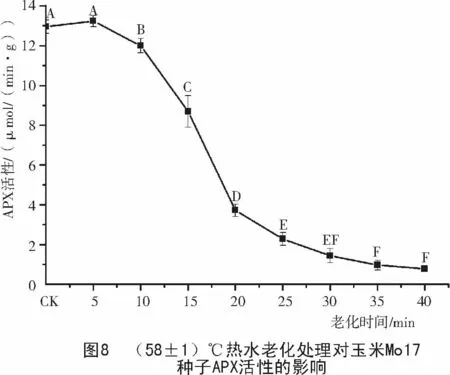
2.4 玉米種子老化過程中APX 活性的變化
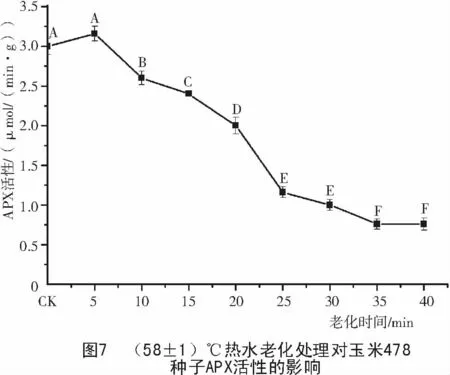
由圖7,8可知,老化處理后,478和Mo17種子的APX活性隨老化時間的延長呈下降的趨勢,2個玉米種子的APX活性差別較大。Mo17種子的APX活性是478種子的4倍,在老化20 min之前,Mo17的APX活性下降幅度遠遠大于478,25 min后二者的APX活性表現相差不大,都降到了較低的水平。
3 討論
植物在正常的生理代謝過程中,O2·-不斷產生同時又在不斷被清除,維持了一個生理性低水平和相對恒定的濃度[8],由此保證了植物的正常生長和發育。種子萌發過程中,O2·-產生速率上升的同時,SOD活性也逐漸增加。SOD活性受到O2·-產生速率的誘導,這種內源SOD活性的上升反映了植物自身的一種保護機制,而這種SOD活性的增加使O2·-限制在一定水平內,使O2·-不會對植物本身造成傷害[8-9]。種子在老化過程中細胞內會產生大量的O2·-,SOD的表達合成升高。過多的活性氧可以與生物大分子如蛋白質、核酸等發生反應,改變其原來的生理功能,引起植物體傷害甚至死亡。因此,在植物細胞中保持正常的生理代謝,抗氧化酶類SOD,POD,CAT和APX的作用十分明顯[10-12]。
通過對玉米自交系Mo17,478種子人工老化過程中SOD,POD,CAT,APX活性變化規律進行研究,揭示了不同品種的活力不同,不同品種自身的抗氧化調節能力不同。老化使玉米種子細胞內POD,CAT和APX的活性降低,清除過氧化物的能力下降,造成H2O2等活性氧在種子內積累,導致細胞膜的膜脂質過氧化,進而對細胞造成嚴重損傷,導致種子活力發生不可逆轉的下降,直到死亡[13-15]。
本試驗結果表明,除SOD活性隨老化時間的延長不斷升高外,POD,CAT和APX的活性均隨老化時間的延長整體呈下降趨勢。因此,在玉米種質資源保存和商品玉米種子貯藏中,POD,CAT和APX的活性可作為檢測玉米種子活力高低的一個重要指標。
[1]張曉芳.玉米種質資源品質性狀的鑒定與評價[J].玉米科學,2006,14(1):18-20.
[2]馬俊華,劉建軍.玉米種子萌發初期生理特性的變化研究[J].中國農業科技導報,2011,13(4):99-103.
[3]Mccord JM,Fridovich I.Superoxidedismutase:An enzymic function for erythrocuprein(Hemocuprein)[J].J Bilo Chem,1969,224:6049-6055.
[4]McDonald MB.Seed deterioration physiology,repair and assessment[J].Seed Scienceand Technology,1999,27:172-177.
[5]李忠光,李江鴻,杜朝昆,等.在單一提取系統中同時測定五種植物抗氧化酶[J].云南師范大學學報,2002,22(6):44-48.
[6]史偉,張世鮮,田甜.抗體酶研究新進展[J].內蒙古農業科技,2011(1):26-27.
[7]喬燕祥,高平平,馬俊華,等.2個玉米自交系在種子老化過程中的生理特性和種子活力變化的研究[J].作物學報,2003,29(1):123-127.
[8]李雪峰,鄒學校,劉志敏.辣椒種子人工老化及劣變的生理生化變化 [J].湖南農業大學學報:自然科學版,2005,31(3):265-268.
[9]傅愛根,王愛國,羅廣華.大豆萌發過程的活性氧代謝[J].熱帶亞熱帶植物學報,1997,5(4):32-38.
[10]谷建田,孔祥輝.老化前后濕干處理對番茄及菜花種子的效應[J].種子,1997(3):13-18.
[11]莊偉建.花生種子藥劑處理貯藏技術的生理基礎研究[J].花生學報,2002,31(2):1-6.
[12]孫春青,楊偉,戴忠良,等.人工老化處理對結球甘藍種子生理生化特性的影響[J].西北植物學報,2012,32(8):1615-1620.
[13]和紅云,田麗萍,薛琳.植物抗寒性生理生化研究進展[J].天津農業科學,2007,13(2):14-17.
[14]衣瑩,張玉龍,郭志富,等.冬小麥葉片對低溫脅迫的生理響應[J].華北農學報,2013,28(1):144-148.
[15]李健,趙宇,李錦錦.不同基因型玉米葉片衰老與活性氧代謝的關系及其調控[J].華北農學報,2011,26(1):131-135.
