香蕉泛素結合酶基因MaUCE2在非生物脅迫下的表達分析
2013-12-23 05:12:24王安邦金志強劉菊華賈彩紅張建斌苗紅霞徐碧玉
生物技術通報 2013年5期
關鍵詞:植物
王安邦 金志強 劉菊華 賈彩紅 張建斌 苗紅霞 徐碧玉,
(1. 海南大學農學院,海口 571101;2. 中國熱帶農業科學院海口實驗站 香蕉研究所,海口 571101;3. 中國熱帶農業科學院熱帶生物技術研究所,海口 571101)
植物固著生長的特性使其在生長發育過程中不可避免地與外界環境有著密切的關聯[1]。當面臨逆境脅迫時,植物必須能夠應對這些逆境才能存活[2-4]。逆境脅迫下,細胞產生各種異常的或受損的蛋白質,這些異常蛋白如果在細胞內積累過多,會影響細胞的各種代謝,從而破壞細胞結構和功能的完整性[5]。泛素-蛋白酶體途徑(ubiquitin-proteasome pathway,UPP)是目前已知的最重要、有高度選擇性的蛋白質降解途徑[6],它通過調節功能蛋白質的周轉(turnover)或降解不正常蛋白,實現對多種代謝過程的調節[7]。該途徑的關鍵酶主要包括泛素活化酶(ubiquitin-activating enzyme,E1)、泛素結合酶(ubiquitin-conjugating enzyme,E2)和泛素-蛋白連接 酶(ubiquitin-protein ligating enzyme,E3)[8]。其中E2是種類豐富的酶系,更是連接E1和E3的橋梁[9],對泛素化修飾的特異性和精確時空性起關鍵作用[10]。
植物E2蛋白基因(LeUBC1)首次從番茄cDNA文庫中分離,并且受熱激和重金屬誘導表達增強[11],說明E2蛋白基因在發生脅迫時可能參與細胞異常蛋白的降解過程。徐晨曦等[12]從檉柳中獲得E2基因,轉入煙草中異源表達發現該基因能明顯提高煙草的耐旱性。Cui等[13]發現擬南芥泛素結合酶UBC32定位在內質網上,參與異常蛋白質降解過程,可以提高擬南芥的耐鹽性。從馬鈴薯[14]和大豆中[15]也克隆到E2基因,轉入擬南芥后可提高擬南芥的抗鹽性和耐旱性。近年來,對E2基因功能研究盡管也取得了一定進展,但相對緩慢[16],而且關于香蕉泛素結合酶基因與植物的抗逆性關系的研究尚未見報道。
香蕉喜高溫,怕低溫,生長期對水分需求量大,因而溫度和水分是香蕉分布的主要限制因子。然而,由于全球異常氣候的增多,香蕉處于高溫、低溫、干旱、鹽堿、風害等逆境的時間也增多,這嚴重影響了香蕉產量并制約著香蕉產業的發展[17],因此香蕉抗逆性研究顯得尤為重要。
李羽佳等[18]從香蕉根系均一化全長cDNA 文庫中克隆了一個香蕉泛素結合酶基因,命名為MaUCE2,經生物信息學分析發現該基因有典型的保守催化結構域,內含有一個高度保守的半胱氨酸(cysteine)酶活性位點[7,16]。為了進一步研究該基因與非生物脅迫的關系,本研究利用實時定量PCR(qPCR)技術探究MaUCE2基因在干旱、鹽、低溫等非生物脅迫下的表達特性,旨在為研究E2基因生物學功能及其應用奠定基礎。
1 材料與方法
1.1 材料
供試材料為巴西蕉(Musa acuminata L. AAA group,‘Brazilian’),采自中國熱帶農業科學熱帶生物技術研究所香蕉種植園。
1.2 方法
1.2.1 香蕉MaUCE2熒光定量表達分析 采用李燕強等[19]改良的CTAB法提取香蕉總RNA。每個樣品取1 μL檢測RNA質量和濃度后,利用M-MLV逆轉錄酶(TaKaRa公司)合成cDNA鏈。
選用NCBI上已登錄的香蕉MaActin 片段為內參,引物序列根據已登錄序列進行設計,MaActin 1(5'-CGAGGCTCAATCAAAGA-3')和MaActin 2(5'-ACCAGCAAGGTCCAAAC-3')。MaUCE2 引物序列為MaUCE2-1(5'-CAAGACCGACAGAGCCAAGT-3')和MaUCE2-2 (5'-CCTTAGGTGGTTTGAATGG-3')。使用儀器為Mx3000P(Stratagene,USA),PCR反應體系為25 μL,PCR 產物的長度為232 bp。熒光定量PCR 的反應程序如下:95℃預變性30 s;95℃變性7 s,55℃退火15 s,72℃延伸20 s,循環40次。每個反應重復3 次。
1.2.2 MaUCE2在不同外源脅迫下的表達特性分析
1.2.2.1 香蕉植株的干旱脅迫處理 參照張麗麗等[20]香蕉干旱處理的方法,選取長勢一致、生長健壯的香蕉組培苗,分為4組,每組3株,測定每組各盆中土壤水分含量并保持一致。第1組不做任何處理,作為對照,使土壤相對含水量保持在75%-80%;第2組低度失水干旱脅迫,使土壤相對含水量保持在55%-60%;第3組中度失水干旱脅迫,使土壤相對含水量保持在45%-50%;第4組高度失水干旱脅迫,使土壤相對含水量保持在30%-35%。每組香蕉組培苗置于人工恒溫氣候箱中培養,測定各盆中土壤水分含量,當達到脅迫程度后,立即取每株組培苗根部液氮速凍,提取RNA。
1.2.2.2 香蕉植株的鹽脅迫處理 選取長勢一致、生長健壯的香蕉組培苗進行水培養,在培養液(1/2Hoagland培養液)中加入200 mmol/L NaCl進行鹽脅迫處理。分別于0、2、4、6和8 h分別觀察和取樣,將株苗根部立即用液氮速凍,提取RNA。
1.2.2.3 香蕉植株的低溫脅迫處理 選取長勢一致、生長健壯的香蕉組培苗,分為5組,每組3株,置于人工恒溫氣候箱中培養。第1組溫度設為28℃,作為對照,第2組到第5組分別置于15℃、10℃、7℃、5℃溫度下2 h進行低溫脅迫,當達到脅迫時間后,液氮速凍,提取RNA。
2 結果
如圖1所示,通過實時定量PCR分析發現,干旱處理可以誘導MaUCE2的表達,其表達量隨著干旱程度的加重而逐步上升。在高度干旱脅迫下處理下MaUCE2的表達量最高,為2.32,分別為低度干旱脅迫和中度干旱脅迫時期誘導的表達量的2.11倍和1.25倍。
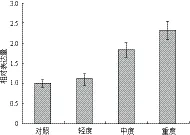
圖1 不同程度水分脅迫下MaUCE2的表達
如圖2所示,高鹽脅迫處理下的MaUCE2的表達量隨NaCl處理時間的增加而略有升高,在脅迫6 h時最高,為1.26。處理2 h、4 h和8 h誘導MaUCE2的表達量分別為1.11、1.13和0.98,與對照差異不大。
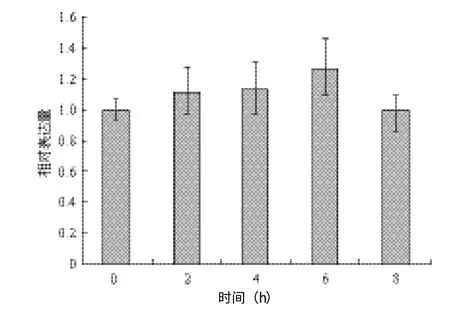
圖2 NaCl脅迫不同時間下MaUCE2的表達
如圖3所示,低溫脅迫可以誘導MaUCE2的表達,其表達量隨著溫度的降低而逐步升高,當溫度降低到5℃時,MaUCE2的表達量最高,為4.08。溫度為15℃和10℃時誘導MaUCE2的表達量為1.26和1.18,與對照差異不大。
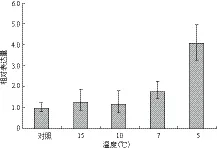
圖3 不同程度低溫脅迫下MaUCE2的表達
3 討論
逆境脅迫下,細胞產生各種異常的或受損的蛋白質,這些異常蛋白如果在細胞內積累過多,會影響細胞的各種代謝,從而破壞細胞結構和功能的完整性,而這些異常蛋白絕大部分是通過泛素-蛋白酶體途徑被降解,因此,泛素-蛋白酶體途徑中關鍵酶的大量表達與植物抗逆性的獲得有著密切的關系,而泛素結合酶E2在E1和E3之間穿梭往返運行,具有結合并轉移泛素到靶蛋白的作用,對泛素化修飾的特異性和精確時空性起關鍵作用。
干旱脅迫是導致導致作物減產的最主要原因[1,21],因此,有關植物應對干旱脅迫反應的研究顯得極為迫切和重要。本研究結果顯示干旱脅迫后MaUCE2表達量顯著提高,說明干旱處理可以誘導MaUCE2的表達。該結果與前人[12,14,15]研究結果相一致,而且本研究發現MaUCE2的表達量在高度干旱脅迫下表達量最高,這可能與高度干旱脅迫下細胞異常蛋白迅速增加有關。
Zhou等[15]克隆了馬鈴薯泛素結合酶基因GmUBC2,異源表達發現該基因能明顯提高擬南芥的耐鹽性。在鹽脅迫下該基因在擬南芥根中表達與對照差異不明顯。本研究結果顯示鹽脅迫后MaUCE2表達量略有上升,最高時是對照的1.26倍,然后基本恢復到正常水平,與對照差異也不明顯。
低溫脅迫下,MaUCE2表達量在溫度為15℃、10℃時上升不明顯,而溫度低到7℃和5℃時該基因表達量迅速上升,在5℃時表達量最大,是對照的4.08倍。這可以與植物適宜生長溫度的范圍有關,有研究表明當溫度小于8℃時香蕉苗開始受到寒害[22],因此當溫度低于8℃后細胞產生各種異常的或受損的蛋白質增多,導致MaUCE2表達量迅速增高。
目前大多數植物的泛素結合酶蛋白功能的研究還處于起步階段,因此從不同植物中克隆及研究泛素結合酶功能顯得尤為重要。本研究初步驗證了香蕉泛素結合酶基因MaUCE2在植物抗逆過程中可能起著重要作用,但MaUCE2在植物抗逆過程中具體的功能和機制尚需深入研究。
4 結論
在鹽、低溫和干旱脅迫下,香蕉泛素結合酶基因MaUCE2表達量發生明顯變化,表明香蕉中的泛素結合酶基因具有響應非生物脅迫能力,可能在香蕉適應逆境中發揮重要作用。
[1] 寧約瑟, 王國梁, 謝旗.泛素連接酶E3介導的植物干旱脅迫反應[J]. 植物學報, 2011, 46 (6):606-616.
[2] Peleg Z, Blumwald E. Hormone balance and abiotic stress tolerance in crop plants[J]. Curr Opin Plant Biol, 2011, 14:290-295.
[3] Ahuja I, de Vos RCH, Bones AM, Hall RD. Plant molecular stress responses face climate change[J]. Trends Plant Sci, 2010, 15:664-674.
[4] Hirayama T, Shinozaki K. Research on plant abiotic stress responses in the post-genome era:past, present and future[J]. Plant J, 2010, 61:1041-1052.
[5] Smalle J, Kurepa J, Yang P et al. The pleiotropic role of the 26S proteasome subunit RPN10 in Arabidopsis growth and development supports a substrate-specific function in abscisic acid signaling[J]. Plant Cell, 2003, 15:965-980.
[6] 郭啟芳, 鄒琦, 王瑋. 植物泛素/26S蛋白酶體通路的生理功能和分子生物學[J]. 植物生理學通訊, 2004, 40(5):533-539.
[7] 黃海杰, 陳雄庭.植物泛素/26S蛋白酶體途徑研究進展[J].中國生物工程雜志, 2008, 28(7):127-132.
[8] Mukhopadhyay D, Riezman H. Proteasome-independent functions of ubiquitin in endocytosis and signaling[J]. Science, 2007, 315:201-205.
[9] Pickart CM. Mechanisms underlying ubiquitination[J]. Annu Rev Biochem, 2001, 70:503-533.
[10] 倪曉光, 趙平.泛素-蛋白媒體途徑的組成和功能[J]. 生理科學進展, 2006, 37(3):255-258.
[11] Feussner K, Feussner I, Leopold I, et al. Isolation of a cDNA coding for an ubiquitin-conjugating enzyme UBC1 of tomato-the first stressinduced UBC of higher plants[J]. FEBS Lett, 1997, 409(2):211-215.
[12] 徐晨曦, 姜靜, 劉甜甜, 等.檉柳泛素結合酶基因(E2s)的序列分析及功能驗證[J]. 東北林業大學學報, 2007, 35(11):1-4.
[13] Cui F, Liu LJ, Zhao QZ, et al. Arabidopsis ubiquitin conjugase UBC-32 is an ERAD component that functions in brassinosteroid-mediated salt stress tolerance[J]. Plant Cell, 2012, 24(1):233-244.
[14] Wan X, Mo A, Liu S, et al. Constitutive expression of a peanut ubiquitin-conjugating enzyme gene in Arabidopsis confers improved water-stress tolerance through regulation of stress-responsive gene expression[J]. J Biosci Bioeng, 2011,111(4):478-484.
[15] Zhou GA, Chang RZ, Qiu LJ. Overexpression of soybean ubiquitinconjugating enzyme gene GmUBC2 confers enhanced drought and salt tolerance through modulating abiotic stress-responsive gene expression in Arabidopsis[J]. Plant Mol Biol, 2010, 72(4-5):357-367.
[16] 王金利, 史勝青, 賈利強.植物泛素結合酶E 2功能研究進展[J]. 生物技術通報, 2010(4):7-8.
[17] 劉菊華, 鄧成菊, 金志強, 等.香蕉乙二醛酶基因MaGLO14的克隆及在非生物脅迫下的功能鑒定[J]. 中山大學學報:自然科學版, 2011, 50(5):87-92.
[18] 李羽佳, 金志強, 劉菊華, 等. 香蕉泛素結合酶基因的克隆及功能初步分析[J]. 熱帶作物學報, 2013.
[19] 李燕強, 金志強, 徐碧玉.香蕉果實RNA提取方法的改進和比較[J]. 福建熱作科技, 2005, 30(2):37-39.
[20] 張麗麗, 徐碧玉, 金志強, 等.香蕉谷胱甘肽過氧化物酶基因MaCPX的克隆和表達分析[J]. 園藝學報, 2012, 39(8):1471-1481.
[21] Luo LJ. Breeding for water-saving and drought-resistance rice (WDR) in China[J]. J Exp Bot, 2010, 61:3509-3517.
[22] 黃朝榮.南寧市香蕉冷害指標及防御措施探討[J]. 廣西農業科學, 1991(3):106-109.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
