陜西楊凌地區(qū)TYLCV病毒生物信息學(xué)分析研究
2014-01-12 03:25:58王玲慧李翠潘銅華劉婧儀崔霞李云洲王巧麗梁燕西北農(nóng)林科技大學(xué)園藝學(xué)院旱區(qū)作物逆境生物學(xué)國家重點(diǎn)實(shí)驗(yàn)室陜西楊凌712100
長江蔬菜 2014年4期
王玲慧,李翠,潘銅華,劉婧儀,崔霞,李云洲,王巧麗,梁燕(西北農(nóng)林科技大學(xué)園藝學(xué)院,旱區(qū)作物逆境生物學(xué)國家重點(diǎn)實(shí)驗(yàn)室,陜西楊凌,712100)
陜西楊凌地區(qū)TYLCV病毒生物信息學(xué)分析研究
王玲慧,李翠,潘銅華,劉婧儀,崔霞,李云洲,王巧麗,梁燕
(西北農(nóng)林科技大學(xué)園藝學(xué)院,旱區(qū)作物逆境生物學(xué)國家重點(diǎn)實(shí)驗(yàn)室,陜西楊凌,712100)
采集楊凌五泉、揉谷和李臺(tái)3個(gè)番茄主產(chǎn)區(qū)表現(xiàn)矮化、黃化及曲葉癥狀的植株嫩葉,克隆番茄黃化曲葉病毒(TYLCV)基因全長并測序,依次得到病毒分離物TYLCV-SXYL2、TYLCV-SXYL3和TYLCV-SXYL4。通過多序列比對(duì)、系統(tǒng)發(fā)育樹構(gòu)建及蛋白質(zhì)結(jié)構(gòu)和理化性質(zhì)預(yù)測等生物信息學(xué)方法進(jìn)行基因組和蛋白質(zhì)的特征分析,結(jié)果表明,楊凌區(qū)3個(gè)TYLCV分離物之間全長核苷酸相似度為99.3%~99.4%,是不伴隨衛(wèi)星分子的單組分病毒,屬TYLCV-IS株系的不同分離物,全長為2 781 nt;與山東壽光病毒分離物TYLCV-SDSG親緣關(guān)系最近,相似度為99.6%,與陜西涇陽的分離物TYLCV-SX8相似度為99.1%,與以色列株系TYLCV-IS相似度達(dá)97.7%~97.8%;編碼6個(gè)蛋白質(zhì),其中CP、Rep、REn為跨膜蛋白,V2、TrAP、C4為胞內(nèi)蛋白,Rep和REn為穩(wěn)定蛋白,CP、V2、TrAP、C4為不穩(wěn)定蛋白。
番茄黃化曲葉病毒;楊凌;番茄;生物信息學(xué)分析
番茄(Solanum lycopersicum L.)是世界上最重要的蔬菜作物之一[1],而番茄黃化曲葉病(Tomato yellow leaf curl disease,TYLCD)為全球番茄生產(chǎn)中最具威脅的“植物癌癥”[2],1961年以色列最先鑒定的番茄黃化曲葉病毒(Tomato yellow leaf curl virus,TYLCV)是其主要病原。TYLCV為單鏈環(huán)狀DNA病毒,屬于雙生病毒科(Geminiviridea)菜豆金色花葉病毒屬(Geminiviridea),由煙粉虱(Bemisia tabaci)以持久性方式傳播[3,4],在寄主細(xì)胞核內(nèi)形成的雙鏈DNA復(fù)制中間體共編碼6個(gè)開放閱讀框。楊凌區(qū)是我國唯一的農(nóng)業(yè)高新技術(shù)產(chǎn)業(yè)示范區(qū),位于陜西省關(guān)中平原中部,2011年楊凌暴發(fā)的TYLCD已成為限制楊凌區(qū)番茄生產(chǎn)的主要因素,但目前未見有楊凌TYLCV全基因組分子特征及編碼蛋白生物信息學(xué)分析的研究報(bào)道。本試驗(yàn)對(duì)楊凌區(qū)五泉、揉谷和李臺(tái)的番茄主產(chǎn)區(qū)進(jìn)行調(diào)查采樣,對(duì)病毒分離物進(jìn)行全長克隆、測序和變異分析,并對(duì)病毒編碼蛋白質(zhì)的氨基酸序列和蛋白質(zhì)跨膜結(jié)構(gòu)、不穩(wěn)定系數(shù)等性質(zhì)進(jìn)行了比較分析,旨在進(jìn)一步明確陜西楊凌番茄黃化曲葉病毒基因組結(jié)構(gòu)和進(jìn)化起源,為進(jìn)一步開展番茄黃化曲葉病的防控工作、番茄分子抗病育種、病毒變異進(jìn)化和病毒編碼蛋白質(zhì)結(jié)構(gòu)功能研究提供科學(xué)依據(jù)。
1 材料與方法
1.1 試驗(yàn)材料與試劑
①毒源樣品 在陜西省楊凌區(qū)五泉、揉谷和李臺(tái)的番茄主產(chǎn)區(qū)采集表現(xiàn)生長遲滯矮化、上部葉片黃化、葉片邊緣向上卷曲、褶皺簇狀、后期不能正常開花結(jié)果癥狀的番茄植株嫩葉各1份,常溫下用采樣袋密封帶回,液氮速凍后置于-80℃保存。
②試劑 Pfu DNA Polymerase購自Fermenents公司,瓊脂糖凝膠DNA回收試劑盒(離心柱型)購自Bioteke公司,質(zhì)粒微量提取試劑盒購自BIOMIGA公司,PrimeSTAR Max DNA Polymerase、加A尾試劑盒A-Tailing Kit、克隆載體PMD18-T和DH5α感受態(tài)大腸桿菌購自TaKaRa公司,試驗(yàn)所用引物由生工生物工程(上海)股份有限公司合成,序列測定由南京金斯瑞生物科技有限公司完成。
1.2 試驗(yàn)方法
①植物總DNA的提取和病毒分子鑒定 DNA的提取用CTAB法,得到的樣品DNA溶液于-20℃保存?zhèn)溆谩YLCV鑒定用李常保等[5]設(shè)計(jì)的TYLCV特異引物TYLCV-F/TYLCV-R;陽性對(duì)照TYLCV-SH2由浙江大學(xué)周雪平教授饋贈(zèng)。DNA-B組分鑒定用雙生病毒DNA-B組分鑒定通用引物PCRc1/PBLv2040[6];衛(wèi)星DNA β鑒定用雙生病毒衛(wèi)星DNA β鑒定通用引物Beta01/Beta02[7],引物序列和目標(biāo)條帶大小見表1。

表1 引物序列表及預(yù)期擴(kuò)增條帶
②分子克隆和測序 用TYLCV特異引物TYLCV-F/TYLCV-R,Pfu DNA Polymerase擴(kuò)增部分片段(約500 bp)后回收,用A-tailing Kit加A尾后克隆至PMD18-T載體,轉(zhuǎn)化入DH5α感受態(tài)大腸桿菌,經(jīng)氨芐抗性篩選、菌液PCR鑒定及提取質(zhì)粒酶切驗(yàn)證后用于測序。依據(jù)測得的519 bp序列用Primer Premier軟件(Version 5.0,Premier,Canada)設(shè)計(jì)相鄰背向引物SX-F/SX-R,用PrimeSTAR Max DNA Polymerase擴(kuò)增病毒基因組全長,經(jīng)克隆并測序得到基因組全序列。
③序列比對(duì)分析和進(jìn)化樹的構(gòu)建 通過NCBI Blast(Basic local alignment search tool)尋找TYLCV同源序列信息。用 DNAStar Package的 MEGA(Version 5.05,Madison,Wis,USA)軟 件[8]中 的MUSCLE算法進(jìn)行多序列比對(duì)分析后,用鄰近相連法(Neighbor-joining)構(gòu)建基于病毒DNA-A核苷酸全序列的系統(tǒng)進(jìn)化樹。用DNAMAN(Version 5.2.2,Lynnon Biosoft,Quebec Canada) 軟件構(gòu)建不同TYLCV分離物的同源矩陣。
④病毒基因組結(jié)構(gòu)分析 病毒開放閱讀框的查找利用NCBI的ORF Finder(Open Reading Frame Finder,http://www.ncbi.nlm.nih.gov/gorf/gorf.html)在線工具。病毒基因組結(jié)構(gòu)圖利用DNAStar Package中的SeqBuilder軟件繪制。
⑤蛋白質(zhì)跨膜結(jié)構(gòu)和理化性質(zhì)的預(yù)測及分析 病毒蛋白質(zhì)的跨膜結(jié)構(gòu)利用TMpred(http:// www.ch.embnet.org/software/TMPRED_form.html)在線工具預(yù)測。蛋白質(zhì)理化性質(zhì)利用 ExPASy的ProtParam(http://web.expasy.org/protparam/)在線工具進(jìn)行分析。
2 結(jié)果與分析
2.1 采集樣品DNA病毒全長克隆和聚類分析
①TYLCV病毒分子鑒定 通過用TYLCV特異引物TYLCV-F/TYLCV-R對(duì)樣品DNA進(jìn)行PCR鑒定,結(jié)果顯示,3個(gè)樣品TYLCV-SXYL2、SXYL3和SXYL4都為帶毒植株(圖1),并克隆測序得到519 bp序列。以雙生病毒DNA-B組分通用引物PCRc1/PBLv2040進(jìn)行PCR擴(kuò)增,未得到預(yù)期500~650 bp大小的條帶,以雙生病毒衛(wèi)星DNA β鑒定通用引物Beta01/Beta02進(jìn)行PCR反應(yīng),也未得到預(yù)期1 200~1 400 bp大小的條帶。結(jié)果表明,陜西楊凌地區(qū)侵染番茄的TYLCV為不含DNA-B且不伴隨衛(wèi)星DNA β的單組分病毒,只含DNA-A。

圖1 利用TYLCV-F/TYLCV-R進(jìn)行病毒分子鑒定
②DNA-A全長的同源矩陣 選擇NCBI登錄的國內(nèi)外不同地區(qū)分離的19個(gè)TYLCV分離物(表2),用DNAMAN多序列比對(duì)后構(gòu)建同源矩陣(表3)。發(fā)現(xiàn)楊凌區(qū) 3個(gè)分離物 TYLCV-SXYL2、SXYL3、SXYL4核苷酸序列全長之間相似度為99.3%~99.4%,且和TYLCV-IS相似度為97.7%~97.8%。楊凌區(qū)的3個(gè)病毒分離物與山東壽光分離物TYLCV-SDSG的相似度都為99.6%,與陜西涇陽的分離物TYLCV-SX8的相似度都為99.1%。

表2 用于序列比對(duì)和系統(tǒng)進(jìn)化樹構(gòu)建的番茄黃化曲葉病毒
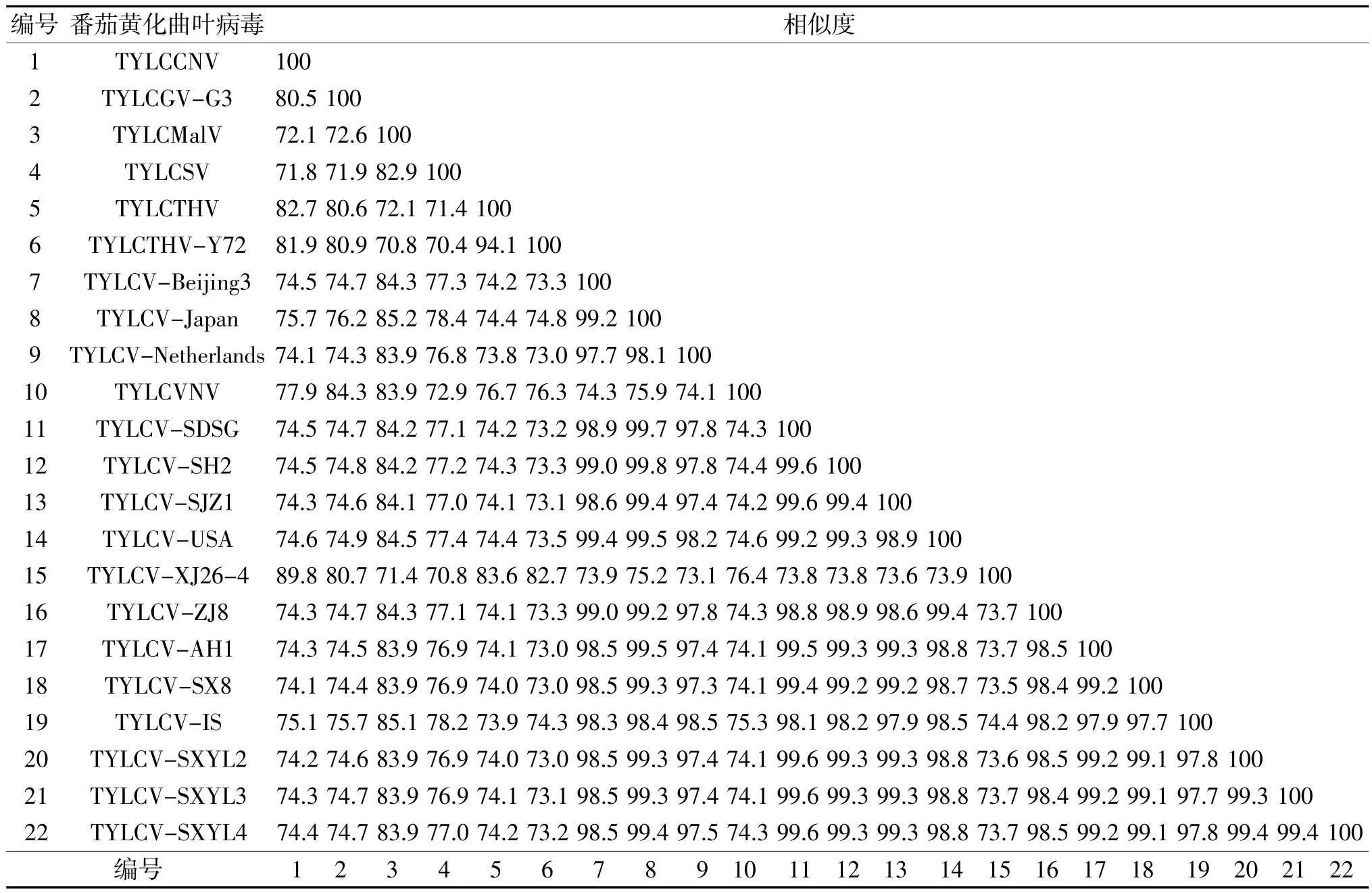
表3 基于DNA-A核苷酸全序列構(gòu)建的同源矩陣
③基于DNA-A全長的系統(tǒng)進(jìn)化樹 陜西楊凌3個(gè)TYLCV分離物同19個(gè)不同國家地區(qū)的TYLCV通過MEGA5.05軟件中MUSCLE算法多序列比對(duì)后,用鄰近相連 (Neighbor-Joining)法構(gòu)建系統(tǒng)進(jìn)化樹 (圖2)發(fā)現(xiàn),楊凌分離物和山東壽光分離物TYLCV-SDSG、河北石家莊分離物TYLCV-SJZ1、陜西涇陽分離物TYLCV-SX8、安徽分離物TYLCV-AH1、北京分離物TYLCV-Beijing3、美國分離物TYLCV-USA及浙江分離物TYLCV-ZJ8在同一小支上,且與山東壽光分離物TYLCV-SDSG親緣關(guān)系最近。上述分離物還與上海分離物TYLCVSH2[9]、日本分離物TYLCV-Janpan、荷蘭分離物TYLCV-Netherlands和以色列株系TYLCV-IS在同一大支上。意大利撒丁島分離物TYLCSV和西班牙馬加拉分離物TYLCMalV在同一支。廣東分離物TYLCGV-G3和越南分離物TYLCVNV在同一支。新疆分離物TYLCV-XJ26-4和廣西分離物中國番茄黃化曲葉病毒TYLCCNV為同一支。云南分離物TYLTHV-Y72和泰國分離物TYLCTHV在同一支。由此可知,我國番茄黃化曲葉病毒的分布和種類同樣有較大的復(fù)雜性和多樣性,而楊凌番茄黃化曲葉病毒分離物很可能由最早發(fā)現(xiàn)的TYLCV-IS發(fā)展而來,且與山東壽光分離物親緣關(guān)系最近,基于楊凌地區(qū)與山東壽光種苗交易頻繁的現(xiàn)狀,推測很可能是因?yàn)楦胁∮酌缃灰讕砹瞬《緜鞑ヅc蔓延。

圖2 基于DNA-A全長核苷酸序列相似性構(gòu)建的系統(tǒng)進(jìn)化樹
2.2 病毒DNA-A全長序列和基因組結(jié)構(gòu)
設(shè)計(jì)背向引物PCR擴(kuò)增并克隆得到病毒DNA-A全長。測序結(jié)果表明,TYLCV-SXYL2、SXYL3、SXYL4(GenBank登錄號(hào)依次為KC138545、KC138544、KC138543)全長都為2 781 nt,共編碼6個(gè)開放閱讀框(圖3),在病毒鏈上編碼外殼蛋白和AV2蛋白,在互補(bǔ)鏈上編碼復(fù)制相關(guān)蛋白、轉(zhuǎn)錄激活子、復(fù)制增強(qiáng)子和AC4蛋白。在AC1和AV2之間有313 nt的非編碼區(qū),也叫基因間隔區(qū)。
①基因間隔區(qū) 基因間隔區(qū) (Intergenic Region,IR)位于1~147 nt和2 616~2 781 nt,共含313個(gè)核苷酸,有調(diào)控病毒復(fù)制和轉(zhuǎn)錄起始必須的元件,含有病毒復(fù)制和轉(zhuǎn)錄所必需的結(jié)構(gòu)域以及莖環(huán)結(jié)構(gòu),莖環(huán)頂端有保守的九核苷酸TAATATT/AC序列。由于IR區(qū)相對(duì)于編碼區(qū)選擇壓小,也是病毒變異最活躍的區(qū)域。通過對(duì)TYLCV-SXYL2、SXYL3、SXYL4和TYLCV-IS的IR區(qū)核苷酸序列比較 (圖4)發(fā)現(xiàn),其特征序列中莖環(huán)頂端的九核苷酸TAATATT/AC(位于2 775~2 781 nt及1~2 nt)保守序列、TATA box和TATATA box,與TYLCV-IS一致,但CAAT box和5~8 nt短重復(fù)序列已表現(xiàn)出與TYLCV-IS有較大差異,其中短重復(fù)序列是Rep蛋白的結(jié)合位點(diǎn)。
②編碼蛋白的結(jié)構(gòu)和性質(zhì)分析 為進(jìn)一步了解楊凌番茄黃化曲葉病毒分離物編碼蛋白質(zhì)氨基酸序列的變異特點(diǎn)和更好地進(jìn)行蛋白質(zhì)性質(zhì)分析,將TYLCV-SXYL4與19個(gè)其他地區(qū)分離物及TYLCV-SXYL2、SXYL3編碼的6個(gè)蛋白質(zhì)氨基酸序列相似度進(jìn)行比較分析 (表5),結(jié)果顯示,楊凌3個(gè)分離物和越南番茄黃化曲葉病毒TYLCVNV、以色列TYLCVIS轉(zhuǎn)錄激活子TrAP的氨基酸序列完全相同。TYLCV-SXYL3、SXYL4和山東壽光分離物TYLCV-SDSG及河北石家莊分離物TYLCV-SJZ1復(fù)制增強(qiáng)子REn的氨基酸序列完全相同,且與TYLCV-IS的REn氨基酸序列相比,第58位由纈氨酸(Valine,V)突變?yōu)楸彼幔ˋlanine,A)、第59位由甲硫氨酸 (Methionine,M) 突變?yōu)榱涟彼幔↙eucine,L)、第94位由天冬氨酸(Aspartic acid,D)突變?yōu)槔野彼幔═yrosine,Y)、第124位由谷氨酸(Glutamic acid,E)突變?yōu)楸彼帷YLCV-SXYL2、SXYL3、SXYL4和日本分離物TYLCV-Japan、山東壽光分離物TYLCV-SDSG、上海分離物TYLCV-SH2、河北石家莊分離物TYLCV-SJZ1的C4蛋白氨基酸序列完全相同,與TYLCV-IS的C4蛋白氨基酸序列相比,第64位由脯氨酸(Proline,P)突變?yōu)榻z氨酸(Ser,S)、第67位由甲硫氨酸突變?yōu)楫惲涟彼幔↖soleucine,I)、第84位由賴氨酸(Lysine,K)突變?yōu)榫彼幔ˋrginine,R)。

圖4 楊凌番茄黃化曲葉病毒與以色列番茄黃化曲葉病毒基因間隔區(qū)分子特征比較

表5 TYLCV-SXYL4與19個(gè)病毒分離物及TYLCV-SXYL2、SXYL3的蛋白質(zhì)氨基酸序列比較
通過TMpred在線工具分別對(duì)楊凌3個(gè)病毒分離物編碼的6個(gè)蛋白進(jìn)行跨膜結(jié)構(gòu)預(yù)測發(fā)現(xiàn),CP、Rep、REn存在跨膜結(jié)構(gòu)(圖5),而V2、TrAP、C4為胞內(nèi)蛋白;楊凌3個(gè)病毒分離物的CP、V2、Rep和REn氨基酸序列有1~2個(gè)位點(diǎn)的區(qū)別,沒有影響蛋白質(zhì)的跨膜結(jié)構(gòu)的預(yù)測結(jié)果。
利用ExPasy的ProtParam在線 工 具 對(duì) TYLCV-SXYL2、SXYL3、SXYL4和TYLCV-IS編碼蛋白的理論等電點(diǎn)、不穩(wěn)定系數(shù)和親水性平均系數(shù)進(jìn)行預(yù)測和比較分析(表6),發(fā)現(xiàn)楊凌番茄黃化曲葉病毒分離物和TYL-CV-IS編碼AV2蛋白和REn的理論等電點(diǎn)差異較大。按不穩(wěn)定系數(shù)<40為穩(wěn)定蛋白、不穩(wěn)定系數(shù)>40為不穩(wěn)定蛋白推測,CP、V2、TrAP、C4為不穩(wěn)定蛋白,Rep和REn為穩(wěn)定蛋白。

圖5 楊凌番茄黃化曲葉病毒分離物TYLCVSXYL2的CP、Rep、REn跨膜結(jié)構(gòu)預(yù)測結(jié)果
3 討論與結(jié)論
為了進(jìn)一步明確引起楊凌地區(qū)不同番茄主產(chǎn)區(qū)番茄黃化曲葉病害的病原種類和分子特征,依據(jù)Begomovirus分類中同時(shí)滿足外殼蛋白氨基酸序列相似度>90%和核苷酸全長相似性>89%才可能為同一病毒的不同分離物的標(biāo)準(zhǔn)[10],本研究在楊凌區(qū)3個(gè)不同番茄主產(chǎn)區(qū)調(diào)查采樣并對(duì)全基因組進(jìn)行了測序和基因組結(jié)構(gòu)分析。本研究表明,在楊凌區(qū)五泉、揉谷和李臺(tái)的番茄主產(chǎn)區(qū)引起番茄黃化曲葉病的病原確為TYLCV-IS株系的不同分離物 (TYLCV-SXYL2、TYLCV-SXYL3、TYLCV-SXYL4),經(jīng)鑒定,這3個(gè)分離物都為單組分雙生病毒且不伴隨衛(wèi)星分子。李云洲等[11]克隆了楊凌地區(qū)西北農(nóng)林科技大學(xué)園藝實(shí)驗(yàn)場番茄黃化曲葉病毒外殼蛋白基因后發(fā)現(xiàn),其氨基酸序列與以色列株系TYLCV-IS相似度>90%。
陜西省涇陽縣2010年暴發(fā)番茄黃化曲葉病后[12],楊凌地區(qū)各大番茄主產(chǎn)區(qū)在2011年相繼發(fā)現(xiàn)番茄黃化曲葉病。但由系統(tǒng)進(jìn)化樹構(gòu)建及病毒編碼的6個(gè)蛋白氨基酸相似度比對(duì)結(jié)果發(fā)現(xiàn),楊凌與山東壽光分離物TYLCV-SXSG親緣關(guān)系比與地理位置最近的涇陽分離物TYLCV-SX8的近,病毒蛋白特別是TYLCV-SXYL3、SXYL4編碼的REn的氨基酸序列相似度與TYLCV-SDSG達(dá)100%,與涇陽TYLCV-SX8僅為97.8%,這說明了植物病毒的傳播流行不僅受地理位置、氣候環(huán)境的影響,人為因素也起著越來越重要的作用。
雖然TYLCV基因組小,編碼的蛋白數(shù)量有限,但其與寄主的互作及致病機(jī)理仍然不清楚[13]。本研究對(duì)不同TYLCV編碼的蛋白質(zhì)間的氨基酸相似度、蛋白質(zhì)的理化特性和跨膜結(jié)構(gòu)進(jìn)行了生物信息學(xué)分析及比較,表明CP、Rep、REn為跨膜蛋白,V2、TrAP、C4為胞內(nèi)蛋白,Rep和REn為穩(wěn)定蛋白,CP、V2、TrAP、C4為不穩(wěn)定蛋白。
[1]Foolad M R,Panthee D R.Marker-assisted selection in tomato breeding[J].Critical Reviews in Plant Sciences, 2012,31(2):93-123.
[2]Lefeuvre P,Martin D P,Harkins G,et al.The spread of tomato yellow leaf curl virus from the Middle East to the world[J].PLoS Pathogens,2010,6(10):e1001164.
[3]Pan H P,Chu D,Yan W Q,et al.Rapid spread of tomato yellow leaf curl virus in China is aided differentially by two invasive whiteflies[J].PLoS One,2012,7(4):e34817.
[4]Liu B M,Preisser E L,Chu D,et al.Multiple forms of vector manipulation by a plant-infecting virus:Bemata and tomato yellow leaf curl virus[J].Journal of virology,2013,87(9):4 929-4 937.
[5]李常保,柴敏,李季,等.北京番茄黃化曲葉病毒病的發(fā)生及分子檢測[J].中國蔬菜,2010(1):28-30.
[6]Rojas M R,Gilbertson R L,Maxwell D P.Use of degenerate primers in the polymerase chain reaction to detect whitefly-transmitted geminiviruses[J].Plant Disease,1993,77(4): 340-347.
[7]Briddon R W,Bull S E,Mansoor S,et al.Universal primers for the PCR-mediated amplification of DNA β[J].Molecular Biotechnology,2002,20(3):315-318.
[8]Tamura K,Peterson D,Peterson N,et al. MEGA5:molecularevolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods[J].Molecular biology and evolution,2011,28(10):2 731-2 739.
[9]Zhang Y P,Zhu W M,Cui H M,et al. Molecular identification and the complete nucleotide sequence of TYLCV isolate from Shanghai of China[J].Virus Genes,2008,36 (3):547-551.
[10]Fauquet C M,Bisaro D M,Briddon R W,et al.Revision of taxonomic criteria for species demarcation in the family Geminiviridea,and an updated list of begomovirus species[J]. Archives of Virology,2003,148(2):405-421. [11]Diaz‐Pendon J A,Truniger V,Nieto C,et al.Advancesin understanding recessive resistance to plant viruses [J].Molecular Plant Pathology,2004,5(3):223-233.
[12]阮濤,楊會(huì)房,楊水英,等.分離自陜西涇陽番茄的番茄黃化曲葉病毒(TYLCD)的分子特征[J].農(nóng)業(yè)生物技術(shù)學(xué)報(bào),2013,21(1):97-105.
[13]Verlaan M G,Hutton S F,Ibrahem R M,et al.The tomato yellow leaf curl yirus resistance genes Ty-1 and Ty-3 are allelic and code for DFDGD-Class RNA-dependent RNA polymerases[J].PLoS Genetics,2013,9 (3):e1003399.
Bioinformatic Analysis of TYLCV in Yangling of Shaanxi Province
WANG Linghui,LI Cui,PAN Tonghua,LIU Jingyi,CUI Xia,LI Yunzhou,WANG Qiaoli,LIANG Yan

We collected the leaves from tomato plants exhibiting stunting,yellowing and curling symptoms in three main tomato production areas Wuquan,Rougu and Litai of Yangling district,and through cloning and sequencing the tomato yellow leaf curl virus(TYLCV)genomes,we got three isolates TYLCV-SXYL2,TYLCV-SXYL3,TYLCV-SXYL4. Bioinformatic approaches including multiple sequence alignment,phylogenetic analysis,protein structure and characteristic prediction were used in genomic and protein analysis,the results showed that,the three isolates,belonged to different isolates of TYLCV-IS strain,were monopartite viruses without satellite,and their full length were 2 781 nt,with similarity degrees ranging from 99.3%to 99.4%.The three Yangling isolates showed closest phylogenetic relationship with TYLCVSDSG,with similarity degree of 99.6%,and the similarity degree was 99.1%to Jingyang isolate TYLCV-SX8 and 97.7%-97.8%to Isreal isolate TYLCV-IS.The results of protein structure prediction showed that they encoded six proteins,and CP,Rep,REn had transmembrane structure,while V2,TrAP,C4 were intracellular proteins.Rep and REn were stable proteins,while CP,V2,TrAP,C4 were unstable proteins.
Tomato yellow leaf curl virus(TYLCV);Yangling;Tomato;Bioinformatic analysis
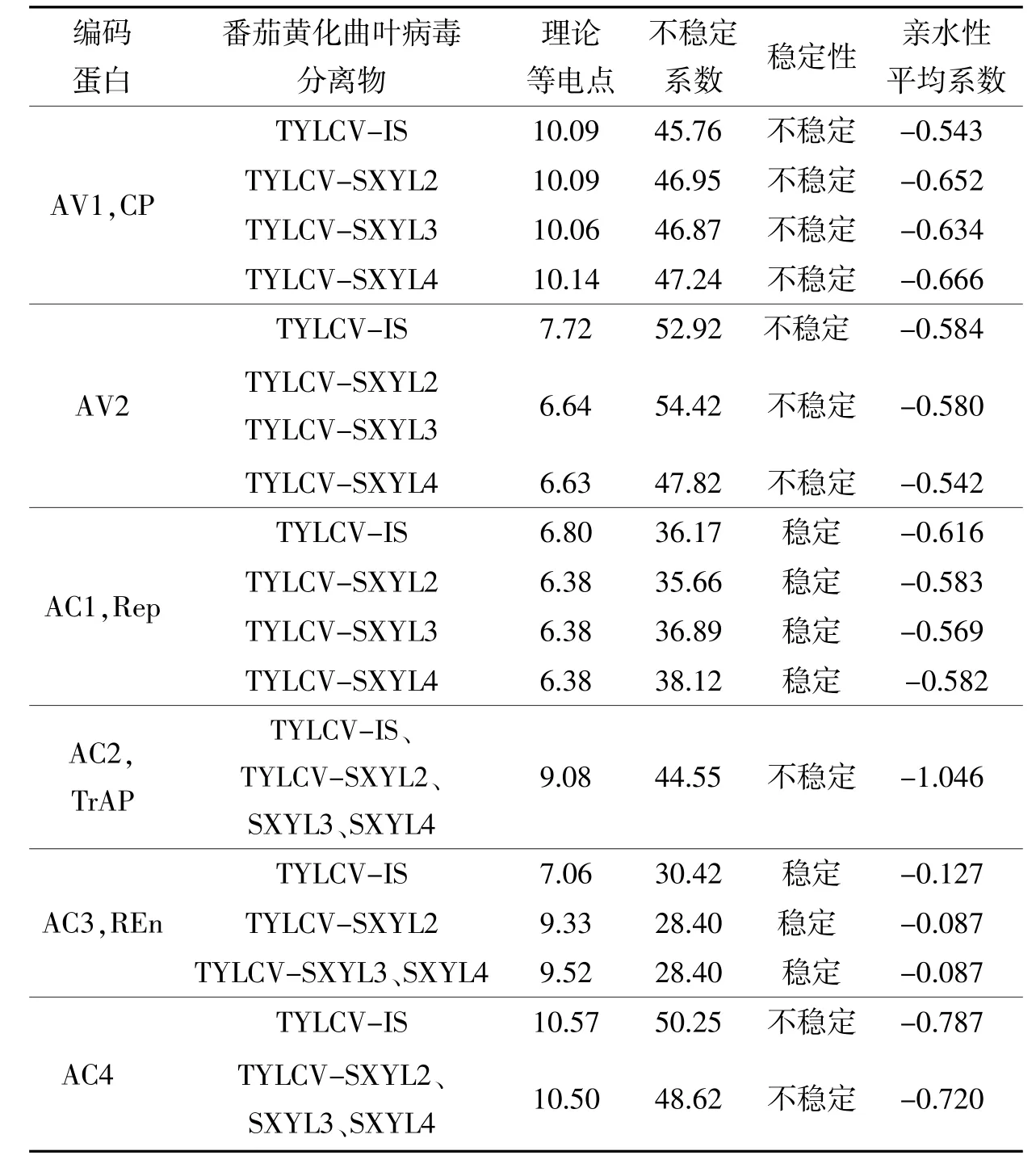
表6 楊凌番茄黃化曲葉病毒分離物與TYLCV-IS編碼蛋白的理化性質(zhì)比較
S436.412.1+1
:A
:1001-3547(2014)04-0054-07
10.3865/j.issn.1001-3547.2014.04.020
陜西省科技統(tǒng)籌創(chuàng)新工程計(jì)劃項(xiàng)目(2011KTCL02-03),陜西省“13115”科技創(chuàng)新工程重大科技專項(xiàng)(2009ZDKG-14)
王玲慧(1992-),女,碩士,主要從事園藝生物技術(shù)研究,
E-mail:sophia_wang@nwsuaf.edu.cn
梁燕(1963-),女,通信作者,教授,博士,博士生導(dǎo)師,主要從事番茄育種和蔬菜種質(zhì)資源研究,
E-mail:liangyan@nwsuaf.edu.cn
2013-11-26
