多金屬富集植物李氏禾根系對銅吸收機理的研究
2014-02-22 09:17:36田靜張學洪陳俊劉杰
生態環境學報 2014年7期
關鍵詞:植物
田靜,張學洪,陳俊,劉杰
多金屬富集植物李氏禾根系對銅吸收機理的研究
田靜,張學洪*,陳俊,劉杰
桂林理工大學環境科學與工程學院,廣西 桂林 541004
李氏禾(Leersia hexandra Swartz)是一種多金屬富集植物,通過研究其根系吸收銅的途徑和方式,有助于解釋李氏禾對銅的富集現象,為開發李氏禾修復銅污染土壤的植物修復技術提供科學依據。本文在溫室水培條件下,利用鈣離子通道抑制劑(LaCl3)和鉀離子通道抑制劑(TEA),研究了李氏禾根系銅吸收與鈣、鉀離子通道的關系;同時,在研究ATP酶抑制劑、解偶聯劑和低溫作用對銅吸收影響的基礎上,探討了李氏禾根系銅吸收與能量代謝的關系。結果表明,在1 mmol·L-1鈣離子通道抑制劑的作用下,李氏禾根對銅的吸收明顯被抑制了(p<0.05)。處理48 h后,李氏禾根中銅的濃度較對照下降了39.2%。這說明李氏禾根吸收銅與鈣離子通道密切相關。在5 mmol·L-1鉀離子通道抑制劑的作用下,李氏禾根對銅的吸收與對照沒用顯著差異。這表明,李氏禾根系對銅的吸收可能不是通過鉀離子通道進行的。ATP酶抑制劑釩酸鈉(Na3VO4)顯著地抑制李氏禾根對銅的吸收(p<0.05)。25 μmol·L-1和50 μmol·L-1Na3VO4處理48 h后,李氏禾根中銅的濃度較對照分別下降了26.2%和31.0%。由此,推測李氏禾根系對銅的吸收是一個消耗能量的過程。該結果與解偶聯劑抑制實驗結果相一致。在25 μmol·L-1和50 μmol·L-1解偶聯劑2,4-二硝基苯酚(DNP)作用下,李氏禾根系中的銅濃度較對照分別下降了25.8%和42.7%。低溫處理對李氏禾根吸收銅的抑制作用較解偶聯劑和ATP酶抑制劑更為明顯。2 ℃下暴露48 h,李氏禾根中銅的濃度較對照(25 ℃)下降了60.1%。這進一步證明了,李氏禾根系對銅的吸收存在消耗能量的主動過程。
李氏禾;銅;吸收;抑制劑;主動運輸
銅是生物必須的營養元素(BURKHEAD等,2009),適量的銅對人和動植物是有益的,但過量的銅則會對人和動植物的生長發育造成危害(倪才英等,2003)。在對環境污染較嚴重的Pb、Hg、Cd、Cu、As、Ni等重金屬中,Cu的排放量較大,每年約為939000 t(SINGH等,2003)。因此,銅污染是亟待解決的環境問題。
植物修復技術(phytoremediation)被認為是修復銅等重金屬污染土壤和水體最經濟有效的方法,以廉價、高效、具有審美價值和長期應用等優點,受到廣泛的推崇(KUMER等,1995;ZHAO和MCGRATH,2009)。該技術的理論核心是通過超富集植物或富集植物對重金屬的吸收和積累,從而去除土壤或水體中的重金屬。因此,超富集/富集植物作為植物修復的基礎和核心載體成為研究的熱點。
李氏禾(Leersia hexandra Swartz)是國內首次報道的鉻超富集植物(張學洪等,2006)。已有研究表明,李氏禾對銅也表現出較強的富集能力(張學洪等,2008)。目前,人們對富集植物吸收轉運金屬離子的機理認識比較有限,主要的研究成果都來源于Cd、As、Zn等幾種重金屬(WEBER等,2004;VERBRUGGEN等,2009;ZHAO等,2009),關于富集植物吸收銅的機理研究相對較少。因此,李氏禾根對Cu的吸收途徑和方式會成為值得研究的科學問題,有助于解釋李氏禾對銅的富集現象,同時也為開發李氏禾修復鉻、銅復合污染環境的植物修復技術提供科學依據。
本文通過水培實驗研究了離子通道抑制劑、ATP酶抑制劑、解偶聯劑和低溫對鉻超富集植物李氏禾根系吸收銅的影響,從能量抑制和離子拮抗的角度探討了李氏禾根系吸收銅的機理,為揭示富集/超富集植物吸收銅的生物學機制提供線索。
1 材料與方法
1.1植物培養
水培試驗的李氏禾采自未受重金屬污染的桂林市雁山鎮田邊。選取生長一致的李氏禾幼苗,用
去離子水洗凈,放入含有1/2強度的改良Hoagland營養液的1.5 L塑料盆中,于人工氣候箱(LRH-550-GSI)中培養,控制植物生長條件為:光照周期12 h,25 ℃白天/20 ℃晚上,相對濕度75%,光照強度7000 Lx。每天加1次超純水來保持培養液的體積,每3 d更換1次營養液。培養10 d后用于以下各種試驗處理。
1.2離子通道抑制劑對Cu吸收的影響
實驗前,將李氏禾幼苗轉移至pH為5.5的250 μmol·L-1CaCl2溶液中預培養。12 h后,對李氏禾幼苗進行如下3組處理:(1)0.3 mmol·L-1CuSO4(對照);(2)1 mmol·L-1鈣離子通道抑制劑(LaCl3)+0.3 mmol·L-1CuSO4;(3)5 mmol·L-1鉀離子通道抑制劑(TEA)+0.3 mmol·L-1CuSO4。所有處理的pH值均調至5.5左右,分別于處理后的1、2、4、8、12、24和48 h收獲植物,每個時間點設3個重復。
1.3代謝抑制劑對Cu吸收的影響
實驗前,將李氏禾幼苗轉移至pH為5.5的250 μmol·L-1CaCl2溶液中預培養。12 h后,對李氏禾幼苗進行如下5組處理:(1)0.3 mmol·L-1CuSO4(對照);(2)25 μmol·L-1ATP酶抑制劑(Na3VO4)+0.3 mmol·L-1CuSO4;(3)50 μmol·L-1Na3VO4+0.3 mmol·L-1CuSO4;(4)25 μmol·L-1解偶聯劑2,4-二硝基苯酚(DNP)+0.3 mmol·L-1CuSO4;(5)50 μmol·L-1DNP+0.3 mmol·L-1CuSO4。所有處理的pH值均調至5.5左右,分別于處理后的1、2、4、8、12、24和48 h收獲植物,每個時間點設3個重復。
1.4低溫對Cu吸收的影響
實驗前,將李氏禾幼苗轉移至pH為5.5的250 μmol·L-1CaCl2溶液中預培養。12 h后,加入含有0.3 mmol·L-1CuSO4的培養液,然后將處理后的植物立刻放進2 ℃的人工氣候箱中培養,分別于處理后的1、2、4、8、12、24和48 h收獲植物,每個時間點設3個重復,以1.2中的對照作為正常溫度(25 ℃)的對照。
1.5樣品分析方法
將植物的根放入含有10 mmol·L-1EDTA的超聲波中清洗3次,每次清洗10 min,去除吸附在植物根表面的金屬離子(LEITA等,1991)。清洗后的根用去離子水洗3次,再用吸水紙把表面的水吸干。將新鮮的樣品放入105 ℃烘箱內殺青30 min,然后80 ℃烘干至恒重,再磨碎。
磨碎的樣品用濃HNO3和H2O2進行消解,銅的含量用火焰原子吸收分光光度法測定。
1.6數據處理
本研究的實驗數據采用三個平行樣測定值的算數平均值±標準差(SD)表示,數據采用SPSS10.0軟件中的單因素方差(ANOVA)進行統計分析,用最小顯著差數法(LSD)對數據進行顯著性檢驗(p<0.05)。采用Origin 8進行作圖。
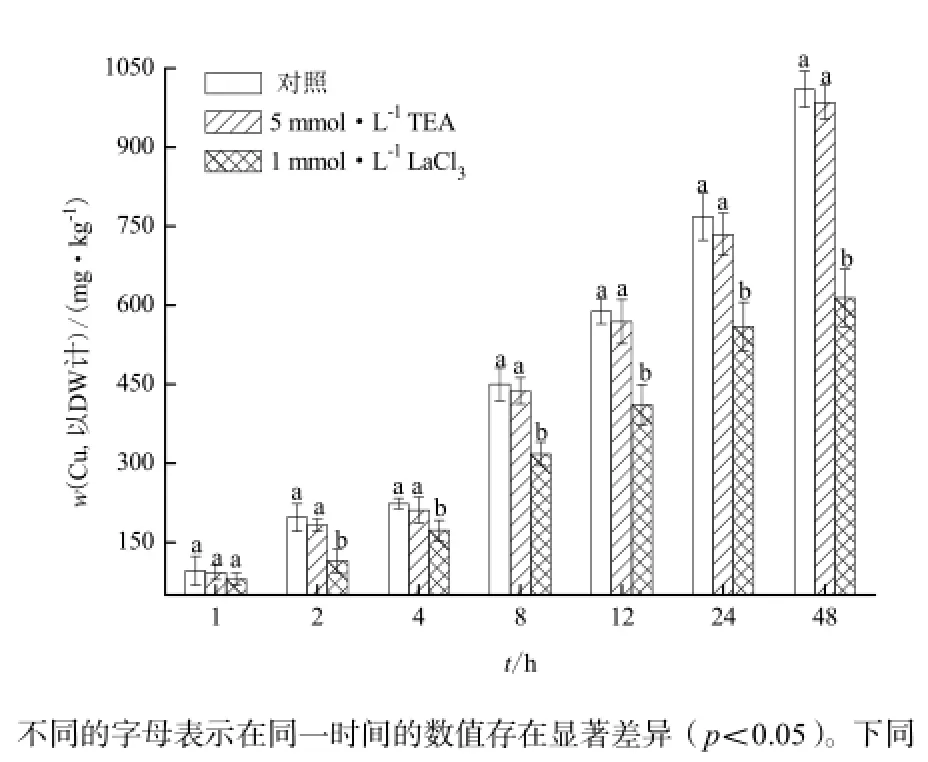
圖1 鉀鈣離子通道抑制劑對李氏禾根吸收Cu的影響Fig.1 Effects of TEA and LaCl3on Cu uptake by roots of L. hexandra
2 結果與分析
2.1離子通道抑制劑對李氏禾根吸收Cu的影響
在離子通道抑制劑的作用下,李氏禾根吸收Cu(0.3 mmol·L-1)的動態變化過程如圖1所示。在1 mmol·L-1鈣離子通道抑制劑(LaCl3)的作用下,李氏禾根對Cu的吸收明顯被抑制(p<0.05,圖1)。處理48 h后,李氏禾根中Cu的濃度較對照下降了39.2%。表明李氏禾對Cu的吸收與鈣離子通道密切相關。然而,在5 mmol·L-1鉀離子通道抑制劑(TEA)的作用下,李氏禾根對Cu的吸收并沒有明顯被抑制,這說明李氏禾對Cu的吸收可能不通過鉀離子通道進行。
2.2代謝抑制劑對李氏禾根吸收Cu的影響
在ATP酶抑制劑(Na3VO4)的作用下,李氏禾根吸收Cu的動態變化過程如圖2所示。25 μmol·L-1和50 μmol·L-1Na3VO4均能顯著抑制李氏禾根對Cu的吸收(p<0.05)。25 μmol·L-1和50 μmol·L-1Na3VO4處理48 h后,李氏禾根中Cu的濃度較對照分別下降了26.2%和31.0%。在同一Cu濃度處理下,Na3VO4對李氏禾根吸收Cu的抑制作用并不隨Na3VO4濃度的增加而增加。這說明在48 h內,25 μmol·L-1和50 μmol·L-1Na3VO4對李氏禾根吸收Cu的抑制作用沒有顯著差異(圖2)。
在解偶聯劑2,4-二硝基苯酚(DNP)的作用下,李氏禾根吸收Cu的動態變化過程如圖3所示。25 μmol·L-1和50 μmol·L-1DNP均能顯著抑制李氏禾根對Cu的吸收(p<0.05)。25 μmol·L-1和50 μmol·L-1DNP處理48 h后,李氏禾根中Cu的濃度較對照分
別下降了25.8%和42.7%。與Na3VO4作用下不同的是,在同一Cu濃度處理下,DNP對李氏禾根吸收Cu的抑制作用會隨著DNP濃度的增加而增加。在48 h內,50 μmol·L-1DNP對李氏禾根吸收Cu的抑制作用顯著大于25 μmol·L-1DNP(p<0.05)。但是在2h之前,DNP濃度對李氏禾根吸收Cu的抑制作用的影響并不明顯。可能的原因是DNP對李氏禾根的代謝抑制具有時間延遲。
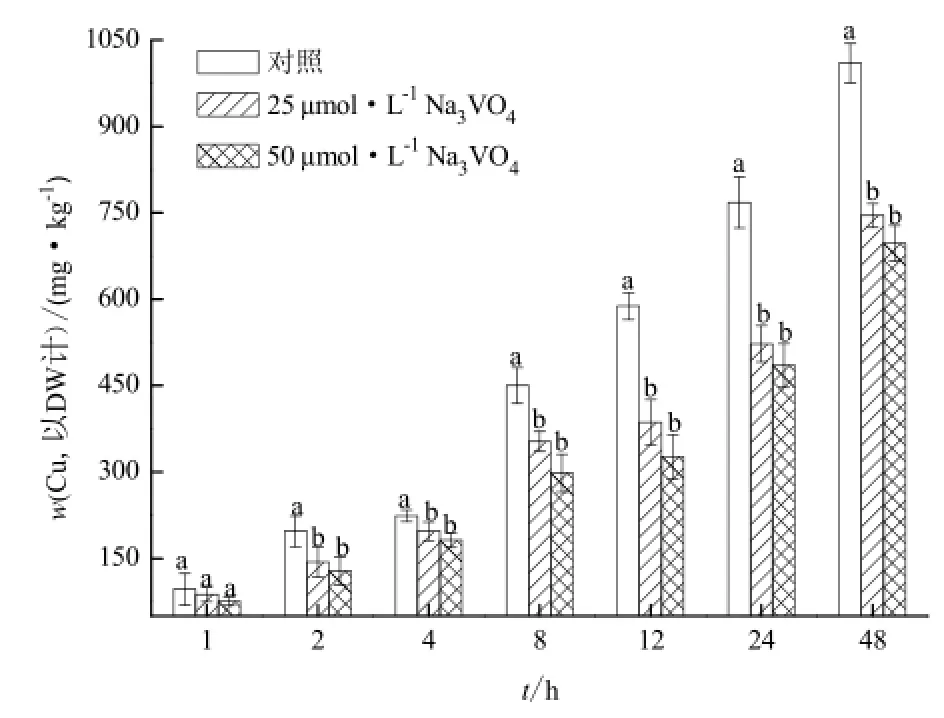
圖2 Na3VO4對李氏禾根吸收Cu的影響Fig.2 Effects of Na3VO4on Cu uptake by roots of L. hexandra
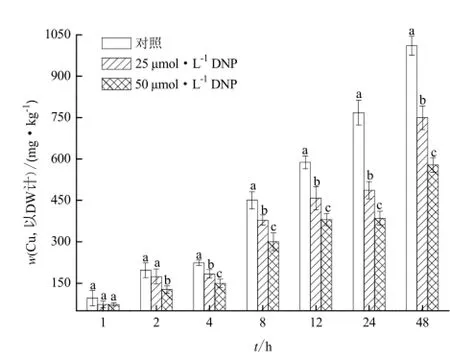
圖3 DNP對李氏禾根吸收Cu的影響Fig.3 Effects of DNP on Cu uptake by roots of L. hexandra
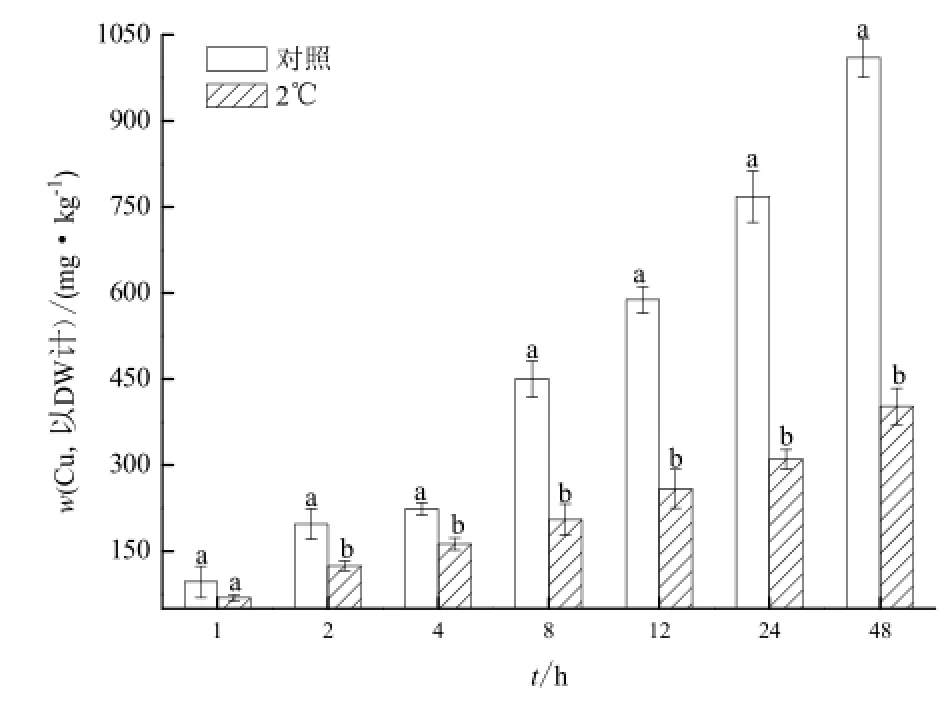
圖4 低溫對李氏禾根吸收Cu的影響Fig.4 Effects of low temperature on Cu uptake by roots of L. hexandra
2.3低溫對李氏禾根吸收Cu的影響
在2 ℃時,李氏禾根吸收Cu的動態變化過程如圖4所示。2 h后,2 ℃處理后的李氏禾根對培養液中Cu的吸收均受到顯著地抑制。8 h時,2 ℃處理的李氏禾根中Cu的濃度較對照(25 ℃)下降了54.5%。在48 h時抑制作用最為明顯,李氏禾根中Cu的濃度較對照(25 ℃)下降了60.1%。這表明李氏禾根對Cu的吸收與植物的代謝水平有關。當代謝水平下降時,李氏禾根對Cu的吸收明顯下降。
3 討論
離子通道是細胞膜上的蛋白質孔道,通過通道門的快速啟閉來控制離子穿過等離子體膜的擴散,是選擇性的離子過濾器(DUTERTRE和LEWIS,2010)。Ca2+和K+通道是兩個重要的陽離子通道,分別可通過LaCl3和TEA進行抑制。LaCl3是一種非特異性的Ca2+通道抑制劑,能夠抑制質膜上ATP酶的活性,并且阻礙細胞內外離子交換(BONDGAARD和BJERREGAARD,2005)。本實驗中,LaCl3顯著抑制了李氏禾根系對銅的吸收,而TEA對李氏禾根系吸收銅沒有抑制作用,這表明李氏禾根系吸收銅與Ca2+通道密切相關,但是可能不通過K+通道。施積炎(施積炎等,2004)等發現海州香薷根對銅的吸收與鈣離子通道密切相關,而鴨跖草根對銅的吸收則可能不通過鈣離子通道。由此可見,重金屬進入植物體內的離子通道的類型不僅與重金屬的種類有關,而且還與植物的種類有密切關系。
P型-ATP酶起著維持細胞內外的離子平衡的作用,是通用的離子泵。微量的釩酸鈉(Na3VO4,μmol級)即可抑制各種P型-ATP酶的活性(WILLIAMS等,2000)。Tabata(TABATA等,1997)等首次報道擬南芥中存在PAA1(一種P型-ATP酶),并參與轉運Cu進入葉綠體。Shikanai(SHIKANAI等,2003)等通過給Cu/Zn過氧化物歧化酶和內囊體腔蛋白質體藍素提供輔酶,證明HMA6(PAA1)負責轉運銅跨膜進入葉綠體。Southron(SOUTHRON等,2004)等在歐洲油菜中發現了銅的轉運體RAN1。Elizabeth(ELIZABETH和MARY,2006)等發現有三種P1B-ATP酶參與銅跨膜轉運到葉綠體中。本實驗中,在0.3 mmol·L-1Cu處理下,25 μmol·L-1和50 μmol·L-1Na3VO4均能顯著抑制李氏禾根對銅的吸收(圖2),這表明李氏禾內存在轉運銅的P型-ATP酶,銅的吸收與P型-ATP酶的活性有密切關系,即李氏禾根對銅可能存在主動吸收的作用。
2,4-二硝基苯酚DNP是一種氧化磷酸化的化學解偶聯劑,由于它可以通過破壞線粒體內膜兩側的質子梯度從而抑制ATP生成,而且解偶聯劑還可以引起線粒體中ATP的大量水解(FELLE和BENTRUP,1977;TRIPATHI等,1995)。由于大部分的生命活動都需要耗能,因此線粒體內的產能過程對生命體極為重要(SCHRAUWEN等,2006)。Sergi(SERGI和DENNIS,2002)等認為Cu+是通過與兩種高親和力轉運體Ctr1、Ctr2和一種低親和力轉運體Fet4結合才從外界跨膜進入細胞內的。本實驗中,25 μmol·L-1和50 μmol·L-1DNP均能顯著抑制李氏禾根對Cu的吸收(圖3),這表明解偶聯劑的使用減少了植物的能量供應,而轉運蛋白通過構象變化來跨膜轉運離子是一個需要能量的過程。進一步說明李氏禾根吸收銅需要消耗能量,可能是一個主動吸收的過程。這一實驗結果與Na3VO4抑制吸收的結果一致。
為了排除代謝抑制劑本身對植物細胞通透性的破壞,從而對植物吸收銅造成影響,因此還分析了低溫(2 ℃)對李氏禾根吸收銅的影響。實驗結果表明,48 h后,低溫對李氏禾根吸收銅的抑制作用最為顯著。因為低溫能夠影響酶的活性從而抑制植物的總體代謝水平(RUELLAND和ZACHOWSKI,2010),因此同時抑制了Cu2+和Cu+的吸收。低溫實驗結果又進一步證實李氏禾根對銅的吸收可能存在主動吸收的機制。施積炎(施積炎等,2004)等研究也表明,海州香薷和鴨跖草對銅存在主動吸收。本研究結果顯示,Na3VO4、DNP和低溫均能顯著抑制李氏禾根對銅的吸收,表明李氏禾根對銅的吸收是一個主動吸收的過程。植物對金屬離子的主動吸收機制有助于將土壤或水體中更多的重金屬離子轉移到植物體內,有利于利用植物修復技術修復被重金屬污染的環境。
4 結論
鈣離子通道抑制劑(LaCl3)顯著抑制了李氏禾根系對銅的吸收,而鉀離子通道抑制劑(TEA)對李氏禾根系吸收銅沒有抑制作用,這表明李氏禾根系吸收銅與鈣離子通道密切相關,可能不通過鉀離子通道進行。
ATP酶抑制劑釩酸鈉(Na3VO4)、解偶聯劑2,4-二硝基苯酚(DNP)和低溫(2 ℃)處理均能顯著地抑制李氏禾根系對銅的吸收,這表明李氏禾根系吸收銅存在主動吸收機制。
BONDGAARD M, BJERREGAARD P. 2005. Association between cadmium and calcium uptake and distribution during the moult cycle of female shore crabs[J]. Aquatic Toxicology, 72: 17-28.
BURKHEAD J L, GOGOLIN K A, ABDEL-GHANY S E, et al. 2009. Copper homeostasis[J]. New Phytologist, 182 (4): 799-816.
DUTERTRE S, LEWIS R J. 2010. Use of venom peptides to probe ion channel structure and function[J]. Journal of Biological Chemistry, 285: 13315-13320.
ELIZABETH P C, MARY L G.. 2006. Put the metal to the petal: metal uptake and transport throughout plants[J]. Current Opinion in Plant Biology, 9(3): 322-330.
FELLE H, BENTRUP F W. 1977. A study of primary effect of the uncoupler carbonyl cyanide m-chlorophyl-hydrazone on membrane potential an conductance in Riccia fluitens[J]. Biochimicaet Biophysica Acta, 464: 179-187.
KUMER P B A N, DUSHENKOV V, RASKIN I. 1995. Phytoextration: The use of plants to remove heavy metals from soils[J]. Environmental Science & Technology, 29: 1232-12391.
LEITA L, CONTIN M, MAGGIONI A. 1991. Distribution of cadmium and induced Ca-bingding proteins in roots, stems and leaves of Phaseolus vulgaris [J]. Plant Science, 77: 139-147.
RUELLAND E, ZACHOWSKI A. 2010. How plants sense temperature[J]. Environmental and Experimental Botany, 69: 225-232.
SCHRAUWEN P, HOEKS J, HESSELLNK M K C. 2006. Putative function and Physiological relevance of the mitochondrial uncoupling protein-3: involvement in fatty acid metabolism?[J]. Progress in Lipid Research, 45(1): 17-41.
SERGI P, DENNIS J T. 2002. Molecular mechanisms of copper uptake and distribution[J]. Department of Biological Chemistry, 6: 171-180.
SHIKANAI T, MüLLER-MOULé P, MUNEKAGE Y, et al. 2003. A P-type ATPase of Arabidopsis, functions in copper transport in chloroplasts[J]. Plant Cell, 15: 1333-1346.
SINGH O V, LABANA S, PANDEY G., et al. 2003. Phytoremediation: an overview of metallic ion decontamination from soil[J]. Applied Microbiology and Biotechnology, 61(5): 405-412.
SOUTHRON J L, BASU U, TAYLOR G J. 2004. Complementation of Saccharomyces cerevisiae ccc2 mutant by a putative P1B-ATPase from Brassica napus supports a copper-transporting function[J]. FEBS Letters, 566: 218-222.
TABATA K, KASHIWAGI S, MORI H, et al. 1997. Cloning of a cDNA encoding a putative metal-transporting P-type ATPase from Arabidopsis thaliana[J]. Biochemical Biophysica Acta, 1326: 1-6.
TRIPATHI R D, RAI U N, GUPTA M, et al. 1995. Cadmium transport in submerged macrophyte Ceratophyllum demersum L. in presence of various metabolic inhibitors and calcium channel blockers[J]. Chemosphere, 31(7): 3783-3791.
VERBRUGGEN N, HERMANS C, SCHAT H. 2009. Molecular mechanisms of metal hyperaccumulation in plants[J]. New Phytol, 181(4): 759-776.
WEBER M, HARADA E, VESS C, et al. 2004. Comparative microarray analysis of Arabidopsis thaliana and Arabidopsis halleri roots identifies nicotianamine synthase, a ZIP transporter and other genes as potential metal hyperaccumulation factors[J]. The Plant Journal, 37(2): 269-281. WILLIAMS L E, PITTMAN J K, HALL J L. 2000. Emerging mechanisms
for heavy metal transport in plants[J]. Biochimica et Biophysica Acta, 1465: 104-126.
ZHAO F J, MA J F, MEHARG A A, et al. 2009. Arsenic uptake and metabolism in plants[J]. New Phytologist, 181(4): 777-794.
ZHAO F, MCGRATH S P. 2009. Biofortifcation and phytoremediation[J]. Current Opinion in Plant Biology, 12: 373-380.
倪才英, 陳英旭, 駱永明. 2003. 土壤-植物系統銅污染與修復的研究進展[J]. 浙江大學學報: 農業與生命科學版, 29(3): 237-243.
施積炎, 陳英旭, 田光明, 等. 2004. 海州香薷和鴨跖草銅吸收機理[J].植物營養與肥料學報, 10(6): 642-646.
張學洪, 陳俊, 李海翔, 等. 2008. 鉻超富集植物李氏禾對銅的富集特征研究[J]. 農業環境科學學報, 27(2): 521-524.
張學洪, 羅亞平, 黃海濤,等.2006.一種新發現的濕生鉻超積累植物——李氏禾(Leersia hexandra Swartz)[J]. 生態學報, 26(3): 950-953.
Mechanisms of Cu Uptake in a Multi-metal Accumulating Plant Leersia hexandra Swartz
TIAN Jing, ZHANG Xuehong*, CHEN Jun, LIU Jie
College of Environmental Science and Engineering, Guilin University of Technology, Guilin 541004, China
Leersia hexandra Swartz is a multi-metal accumulating plant. The study on the mechanisms of Cu uptake by roots of L. hexandra will help to understand how plants accumulate copper. Additionally, understanding Cu uptake mechanisms in plants is also valuable for phytoremediation of Cu-contaminated soil by L. hexandra. In the present work, a series of greenhouse hydroponic experiments were carried out to study the relationship of Cu2+uptake with Ca2+and K+channels using a calcium ion channel blocker (LaCl3) and a potassium ion channel blocker (TEA). Effects of ATPase inhibitor, uncoupler, and low temperature on Cu2+uptake by roots of L. hexandra were also studied, respectively. Based on these experiments, Cu uptake mechanisms in L. hexandra were discussed. The results showed that Ca2+channel blocker significantly limited the Cu uptake (p<0.05). After 48 h treatment with 1 mmol·L-1LaCl3, Cu concentration in the roots was decreased by 39.2% as compared to the control group, which indicated that Cu uptake by roots of L. hexandra was closely related to Ca2+channel. Conversely, K+channel blocker did not inhibit Cu uptake, suggesting that Cu2+did not pass through potassium ion channels. ATPase inhibitor (Na3VO4) significantly limited the Cu uptake (p<0.05). Cu concentrations in the roots exposed to 25 μmol·L-1and 50 μmol·L-1Na3VO4for 48 h were decreased by 26.2% and 31.0%, respectively. Therefore, the Cu uptake by the roots of L. hexandra may need energy consumption. This was supported by the observation that Cu uptake was significantly limited by an uncoupler, 2, 4-dinitrophenol (DNP). Cu concentrations in roots were decreased by 25.8% and 42.7% compared to control after 48 h exposure to 25 μmol·L-1and 50 μmol·L-1DNP. Effects of low temperature on Cu uptake were more pronounced than those of ATPase inhibitor and uncoupler. Cu concentration in roots was decreased by 60.1% at 2 ℃ for 48 h. The result confirmed that the Cu uptake by roots of L. hexandra was an active process dependent on energy consumption.
L. hexandra; Cu; uptake; inhibitors; active transport
X173
A
1674-5906(2014)07-1217-05
國家自然科學基金項目(41361085);“八桂學者”建設工程專項經費資助;廣西危險廢物處置產業化人才小高地基金
田靜(1988年生),女,碩士研究生,主要研究方向為重金屬污染及恢復生態學。E-mail: tianjing421@sina.com
*通訊作者:E-mail: zhangxuehong@x263.net
2014-03-03
田靜,張學洪,陳俊,劉杰. 多金屬富集植物李氏禾根系對銅吸收機理的研究[J]. 生態環境學報, 2014, 23(7): 1217-1221.
TIAN Jing, ZHANG Xuehong, CHEN Jun, LIU Jie. Mechanisms of Cu Uptake in a Multi-metal Accumulating Plant Leersia hexandra Swartz [J]. Ecology and Environmental Sciences, 2014, 23(7): 1217-1221.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
