毛梾種子低溫層積過程中內源激素的變化及其與發芽的關系
2014-03-12 07:24:27李冬林張亞楠金雅琴夏重立
經濟林研究 2014年3期
李冬林,張亞楠,金雅琴,夏重立
(1.江蘇省林業科學研究院,江蘇 南京 211153;2.金陵科技學院園藝學院,江蘇 南京 210038)
毛梾Cornus walteri為我國傳統的木本油料植物,隸屬山茱萸科梾木屬Cornus。其自然分布于我國遼寧南部和華北、華東、中南、西北、西南等地區[1]。毛梾果實含油率高達31.8%~41.3%,出油率為25%~30%[2],其油脂不僅可供食用,還可用作機械和鐘表的潤滑油,也是制皂和油漆的好原料;其葉質地柔軟,富含營養,制成葉粉,各種畜禽均可利用,是一種良好的木本飼料植物;木材堅硬、紋理細致,是一種良質木材,其葉和樹皮可用以提取拷膠,還含有單寧物質,可用作染料;亦是綠化和固土樹種和蜜源植物。
毛梾最常用的育苗方法是播種育苗,但是毛梾種實具有明顯的休眠習性,種皮堅硬,未經處理的種子當年幾乎不能發芽,這給生產中實生苗的培育帶來了一定的難度,并成為毛梾天然更新和引種栽培的主要制約因子。近年來,有關梾木屬種子破除休眠的方法已有研究報道[3-4],但相關機制尚不清楚。為此,對毛梾種子低溫層積過程中各種內源激素的含量進行了測定,分析了內源激素與種子發芽的關系,以期為揭示毛梾種子休眠的內在機理、促進其引種與繁育而提供理論依據。
1 材料與方法
1.1 材料及處理
供試種實采集于江蘇南京中山陵園。種實于2011年10月采集,經揉搓、脫肉、沖洗、晾干,獲得純凈的種實5 kg。經測定,種實圓球型,內果皮淺肉紅色,直徑4.31~5.18 mm,百粒質量為 4.336 4~ 4.430 0 g。
試驗于2011年10月至2012年5月在江蘇省林業科學研究院實驗室進行。種實經脫肉后采用混沙濕藏處理,將種實和濕沙(含水量40%~45%)按照1∶3的體積比混合后置于0~5 ℃的冷庫中。濕沙為新鮮河沙,并經高溫滅菌處理。2011年10月至2012年6月,每月中旬取樣1次,重復3次,共取樣7次。取樣時將種實自濕沙中篩出,用自來水沖洗,并用吸水紙吸干水分,篩揀,獲得純凈種實(含肉紅色內果皮,以下稱為種子)待測。
1.2 內源激素的測定
按照四分法從待測種實中取出部分樣品,將硬果皮敲碎獲得白色測試樣品(含胚、胚乳及種皮)0.5~1.0 g。液氮速凍后,加入4 mL 的提取液(80%冷甲醇)冰浴研磨至勻漿,轉入10 mL的試管中。搖勻后在4 ℃下放置4 h,3 500 r·min-1離心15 min,取上清液。沉淀中加1 mL的提取液,攪勻,置于4 ℃下再提取1 h,然后離心,合并上清液并記錄體積,棄去殘渣。上清液過C18 固相萃取柱純化。測定方法采用酶聯免疫法(ELISA)[5],所用試劑盒由中國農業大學農學院提供。所有樣品均重復測定3次,取平均值。應用Excel作圖并作統計分析。
1.3 發芽試驗與結果測定
將篩揀后的部分純凈種實置于培養箱中作發芽試驗。將種子均勻播種于盛有濕沙的容器盒中,每盒100粒。播種后覆沙2~3 ㎝,均勻淋水,保證沙床的平整和濕潤,然后蓋膜以保濕。沙床溫度保持在25~30 ℃之間。重復試驗3次。播種后每天觀察種子發芽情況,以種子出土作為發芽標準,統計發芽勢及發芽率。
1.4 數據處理
采用SAS612 進行數據統計與分析。采用方差分析方法估算不同處理間的差異顯著性(α=0.05),試驗結果均為平均值±標準差。
2 結果與分析
2.1 低溫層積過程中種子內源激素含量的變化規律
2.1.1 IAA含量的變化規律
低溫層積處理過程中毛梾種子IAA含量的變化規律不明顯(圖1)。未經層積處理的種子其IAA含量最高(81.861 ng·g-1),低溫層積處理后的種子其IAA含量降低為81.861 ng·g-1,但層積一段時間(30 d)后其IAA含量開始下降,并于層積120 d時上升到極值(65.751 ng·g-1),但層積處理150 d后其IAA含量又降至試驗期間的最低值(31.141 ng·g-1)。

圖1 低溫層積過程中毛梾種子IAA含量的變化Fig. 1 Change of IAA content in C. walteri seeds during cold stratification
2.1.2 ABA含量的變化規律
ABA含量變化明顯,整個處理期間呈現出顯著的下降趨勢(如圖2 A所示)。層積處理開始前,種子中的ABA含量最高(329.406 ng·g-1);層積處理后,隨著處理時間的延長,ABA含量一直在下降,并在層積處理150 d后降到極值(235.933 ng·g-1),這時ABA的含量是層積處理開始前的0.716倍。盡管在層積處理末期(180 d后)ABA含量有所回升,但ABA含量的變化趨勢是隨著層積處理時間的延長而下降的。
2.1.3 GA1/3含量的變化規律
GA1/3含量的變化規律如圖2 B所示。在層積處理初期(0~30 d)GA1/3含量緩慢增加,并在層積處理30 d時出現極值(5.734 ng·g-1)。但在層積處理30 d以后GA1/3含量又開始下降,這種下降的趨勢一直持續到試驗結束(180 d)。從GA1/3含量的總體變化趨勢來看,盡管是下降的,但變化一直很平穩。
2.1.4 ZRs與iPAs含量的變化規律
有關文獻[6]報道,細胞分裂素(CTK)具有促進細胞分裂、抑制器官老化、誘導花芽分化及延緩葉片衰老的作用。毛梾種子細胞分裂素ZRs含量的變化規律見圖2 C。從總體趨勢來看,其變化幅度很小。在層積處理開始前,毛梾種子中ZRS的含量為11.088 ng·g-1;層積90 d后,ZRS含量有所下降,并出現了低谷(10.019 ng·g-1),隨后又開始小幅升高;在層積150 d后,ZRS的含量為11.001 ng·g-1,隨后又出現小幅的下降。
iPAs含量的變幅相對較大(如圖2 D所示)。在整個層積處理期內,出現了2次上升2次下降的變化過程,其變化曲線略呈“M”形。在層積處理開始時,iPAs含量(3.553 ng·g-1)為整個層積處理期內的最低值;隨后開始上升,并于層積處理30 d后升至極值(5.937 ng·g-1),隨后開始下降;層積處理90 d時,iPAs含量降為4.557 ng·g-1;層積處理120 d后,再次升至最大值(7.124 ng·g-1);隨后開始降低,并一直持續到試驗結束。

圖2 低溫層積過程中毛梾種子ABA、GA1/3、ZRs和iPAs含量的變化Fig. 2 Changes of ABA, GA1/3, ZRs and iPAs contents in C. walteri seeds during cold stratification
2.2 低溫層積處理過程中毛梾種子中5種內源激素含量比值的變化規律
許多研究結果表明,不同植物激素對植物生理活動的影響不是孤立地發生作用的,而是既相互制約又相互促進的[7-8]。毛梾種子中IAA、GA1/3、ZRs、iPAs的平均值及其與ABA的比值見圖3。
IAA/ABA的變化趨勢與IAA含量的變化趨勢基本相似(圖3 A)。層積處理開始時IAA與ABA的比值最大,出現了最大比值(0.248),隨后下降并出現低谷(0.141)。層積處理30 d后,IAA/ABA開始上升,一直延續到120 d,這時的比值增加到0.231,隨后又重新出現了短時的下降趨勢(處理后150 d,IAA/ABA為0.132)。相對而言,GA1/3/ABA比值在整個層積處理期內波動較小。層積處理初期GA1/3/ABA的比值很低(0.015 7),但隨著層積處理時間的延長而迅速增加;在層積處理30 d時測得的GA1/3與ABA的比值為0.019 3;其后隨著層積處理時間的延長其比值一直很平穩地變化著(如圖3 B所示)。
ZRs/ABA比值的變化趨勢如圖3 C所示。由圖3 C可知,在層積處理前期ZRs/ABA比值的變化平穩。低溫層積處理120 d后,其比值開始增加,并于層積處理150 d后上升到極值(0.046 6)。而iPAs/ABA的變化趨勢與iPAs含量的變化趨勢類似,在整個試驗處理期間出現了2次上升和2次下降的變化過程(圖3 D)。在層積處理開始時,iPAs/ABA為0.010 8,此為整個處理期內的最低值;隨后開始上升,層積處理30 d后升至0.020 0,隨后開始下降,在層積處理90 d時,其比值降為0.016;在層積處理150 d后其比值又升為整個試驗期間的最大值(0.027 8);隨后又有所降低,并一直持續到試驗結束。

圖3 低溫層積過程中毛梾種子中的IAA、GA1/3、ZRs、 iPAs和ABA比值的變化Fig. 3 Changes of ratios of IAA, GA1/3, ZRs, iPAs to ABA in C. walteri seeds during cold stratification
2.3 不同低溫層積處理種子的各發芽指標值
發芽試驗結果表明,不同層積處理時間對種子發芽率與發芽勢均有顯著影響(表1)。方差分析結果表明,不同層積處理時間下毛梾種子的發芽率與發芽勢的差異均極顯著(P<0.01)。未經層積處理的干藏種子其發芽率與發芽勢均為0,說明干藏種子存在嚴重的發芽障礙。從表1中還可以看出,層積處理30 d后,毛梾種子的發芽率與發芽勢均不足10%,可見,短期的沙藏層積處理尚不能解除種子的深休眠。隨著層積處理時間的延長,種子的發芽率與發芽勢均明顯上升。低溫層積處理120與150 d后種子發芽率與發芽勢的變化均不顯著,以層積處理180 d的發芽率與發芽勢分別為41.34%和36.13%,均為最大值。經過層積處理的種子,種子休眠逐步被解除,其發芽率與發芽勢均明顯增加。可見,低溫層積處理對解除毛梾種子的休眠十分奏效。

表1 不同層積處理時間種子各發芽指標值?Table 1 Germination indexes of seeds at different durations of stratification treatment
2.4 低溫層積過程中毛梾種子內5種內源激素的含量及其比值與種子發芽率與發芽勢的相關性
低溫層積過程中毛梾種子中5種內源激素含量及其比值與種子各發芽指標的相關分析結果見表2。從表2中可以看出,IAA含量與IAA/ABA比值之間呈極顯著的正相關(p<0.01),但與iPA含量、ZRS/ABA比值、iPA/ABA比值之間均呈負相關,但其相關性不顯著(p>0.05);ABA含量與ZRS/ABA比值之間呈極顯著的負相關(p<0.01),與iPA/ABA比值間呈顯著的負相關(p<0.05);iPA含量與iPA/ABA比值之間呈極顯著的正相關(p<0.01),iPA/ABA與ZRS/ABA兩個比值間也呈顯著的正相關(p<0.05)。這一結果說明,毛梾種子中各種激素間存在一定的相關性。
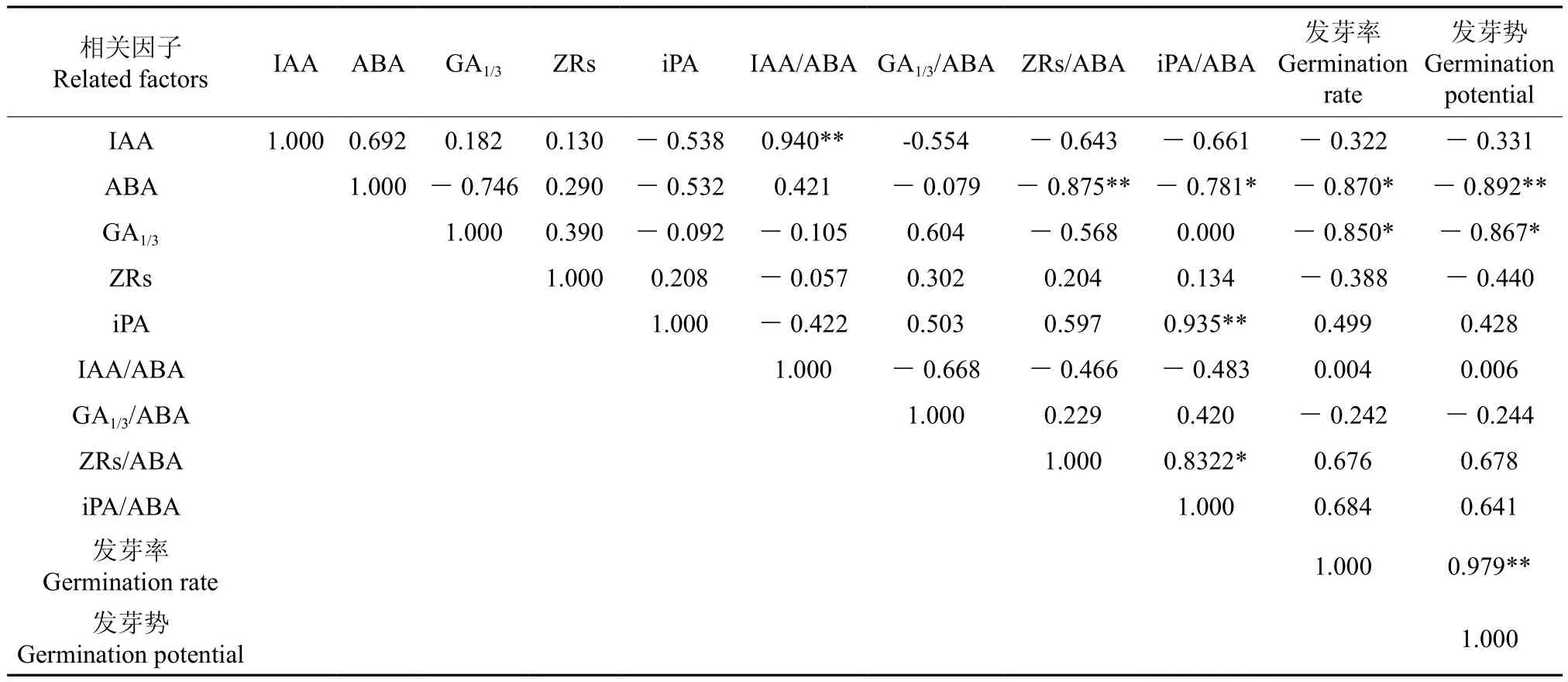
表2 5種內源激素含量及其比值與種子各發芽指標的相關性分析結果?Table 2 Relationship analysis of five kinds of endogenous hormones contents in seeds and their ratios and seeds germination indexes
從5種激素與種子各發芽指標的相關性來看,ABA含量與種子發芽率呈顯著負相關(p<0.05),與發芽勢呈極顯著的負相關(p<0.01),其相關系數分別為0.870、0.892。GA1/3含量與種子發芽率、發芽勢間均呈顯著的負相關(p<0.05),其相關系數分別為0.850、0.867。這一分析結果說明,種子內ABA含量與GA1/3含量對種子發芽的影響最為明顯。
3 結論與討論
在許多物種中,內源激素ABA涉及種子休眠狀態的誘導與維持[9-10],對種子內胚的發育、RNA和蛋白質合成都起到了非常重要的調節作用,被認為是控制種子休眠的主要因素[11],可能會降低胚的吸水能力和滲透勢,阻礙胚根的伸長,抑制種子的萌發[12]。本研究結果表明,在整個低溫層積處理過程中,毛梾種子內ABA含量隨著時間的變化總體上是減少的,這與肉蓯蓉Cistanche deserticola[13-14]、珙桐Davidia involucrata[15]、紫椴Tilia amurensis[16]、 青 錢 柳Cyclocarya paliurus[17]等深休眠植物種子的試驗結果基本相似。隨著低溫層積處理時間的延長,休眠種子中的ABA含量下降,這也許是種子萌發前的啟動信號,對于維持種子中內涵物的生理反應,逐漸解除休眠起到了關鍵作用。
有關IAA在解除種子休眠方面的作用有不同的研究報道。一種研究結果表明,在層積處理中,休眠種子中的IAA含量上升,這對休眠的解除起到了一定的促進作用。如Kochankov等人[18]認為,IAA參與了休眠調節;楊萬霞等人[17]研究認為,青錢柳種子在層積處理中其IAA含量總體呈上升趨勢,這對解除其休眠有利。另有一些研究結果表明,種子休眠與IAA含量的關系不大,過高的IAA含量甚至會起負面作用。如李金克等人[19]研究認為,高濃度的IAA對紅松Pinus koraiensis種子發芽不利;高紅兵等人[20]在對東北紅豆杉Taxus cuspidata的研究中發現,種子層積過程中少量IAA的增加是由胚生長而引起的,IAA與其休眠和萌發的關系不大;黃丹等人[21]在研究美國桂花Osmanthus americanus時也發現,美國桂花種子在層積過程中其IAA含量的變化規律不明顯。本文的研究結果表明,低溫層積處理前期毛梾種子中的IAA含量是增加的,但其達到一定高度后又有所下降。因此認為,作為促進劑的IAA在解除毛梾種子休眠過程中起到了一定的作用,但其作用規律及機理尚待進一步研究。
目前的研究結果表明,用GA處理種子可有效促進種子的萌發[22]。洑香香等人[23]的研究結果表明,GA3浸泡結合層積處理可有效解除山茱萸Cornus officinalis種子休眠。王寧等人[24]的研究結果表明,用400 mg·L-1GA浸泡冬青Ilex purpurea種子48 h可顯著提高GA、CTK和IAA含量,顯著降低ABA含量,從而促進種子萌發。因此,林木種子的休眠與萌發可能取決于內源生長抑制劑和生長激素劑之間的平衡力。Wareing與 Saunders[25]研究認為,GA1/3通過與ABA和環境因子的復雜作用來調節休眠的釋放和種子的萌發。但Bewley[26]認為,GA1/3并不參與對休眠本身的調控,而對促進和維持發芽起作用,能拮抗ABA。試驗中發現,隨著層積處理時間的延長,毛梾種子中的GA1/3與ABA含量間呈負相關性,盡管相關性不十分顯著,但也表明了兩者間存在一定的拮抗關系。而ZRs/ABA比值的變化基本上呈現出緩慢增加的趨勢,因此認為,ZRs在打破毛梾種子休眠、促進種子萌發上起到了一定的作用,這種作用也可能是通過ABA和GA1/3的平衡和拮抗來調節控制的。
植物種子休眠及休眠的解除是十分復雜的生理現象,內源激素的變化及其相互關系是控制休眠的內因,而外部環境條件的變化是其外因[27]。低溫層積處理目前仍是解除種子深休眠的主要手段。發芽試驗結果表明,種子經過層積處理后,其發芽率與發芽勢均明顯上升,并隨處理時間的延長而增加,而未經低溫層積處理的種子其發芽率為0,說明低溫層積處理對毛梾種子深休眠的解除效果十分明顯。但是,對于不同植物的種子及不同的處理方式,以層積處理解除休眠所需的時間是不同的。例如銀杏Ginkgo biloba因其存在生理休眠,需要低溫層積處理90 d才能解除其休眠,而美國山核桃Caraya illinoensis則需要低溫層積處理30~150 d才能順利發芽[28]。本試驗結果說明,低溫層積120與150 d種子發芽率及發芽勢的變化均不顯著,而層積處理180 d后毛梾種子的發芽率與發芽勢均明顯上升,分別達到41.34%和36.13%。由此推斷,要有效解除毛梾種子的深休眠,至少要低溫層積處理150 d以上。本研究結果可為此類植物實生苗培育過程中的種子播種前的預處理提供參考依據。
[1] 《中國樹木志》編輯委員會.中國樹木志[M].第2卷.北京:中國林業出版社,1985:1686-1687.
[2] 柳 鎏,孫醉君.中國重要經濟樹種[M].南京:江蘇科學技術出版社,1986:148-150.
[3] 康永祥,贠玉潔,趙寶鑫,等.毛梾種子萌發特性及幼苗生長規律研究[J].西北林學院學報,2012,27(3):62-67.
[4] 王曉光,李 蔚,劉先貴.光皮毛梾種子催芽技術研究[J].湖北林業科技,2007,(4):26-28.
[5] 李宗庭,周 燮.植物激素及其免疫檢測技術[M].南京:江蘇科學技術出版社,1996.
[6] 唐定臺,徐民新,馮永紅.石竹試管花的誘導及其影響因子的研究[J].園藝學報,1996,23(3):277-280.
[7] Khan A. Primary, preventive and permissive role of hormones in plant systems[J]. Bot Rev, 1975, 41:391-420.
[8] 曹幫華,蔡春菊.銀杏種子后熟生理與內源激素變化的研究[J].林業科學,2006,42(2):32-37.
[9] Leubner-Metzger G. Functions and regulation of b-1,3-glucanase during seed germination, dormancy release and after-ripening[J].Seed Sci Res, 2003, 13:17-34.
[10] Nambara E, Marion Poll A. ABA action and interactions in seeds[J].Trends Plant Sci, 2003,8:213-217.
[11] 韓明玉,張滿讓,田玉命.植物激素對幾種核果類種子休眠破除和幼苗生長的效應研究[J].西北植物學報, 2002, 22(6):1348-1354.
[12] 付婷婷,程紅焱,宋松泉.種子休眠的研究進展[J].植物學報,2009, 44(5): 629-641.
[13] 陳慶亮,王華磊,王志芬,等.低溫層積與外源GA3對肉蓯蓉種子萌發及其內源GA和ABA含量的影響[J].植物生理學通訊,2009,45(3):270-272.
[14] 牛東玲,宋玉霞,郭生虎.肉蓯蓉種子休眠與萌發特性的初步研究[J].種子,2006,25(2): 17-21.
[15] 雷濘菲,彭書明,牛 蓓.珍稀瀕危植物珙桐種子休眠萌發過程中內源激素的變化[J].廣西植物,2009,29(1):66-69.
[16] 楊立學,王海南,張 琳.低溫層積過程中紫椴種子內源激素含量的變化[J].經濟林研究,2012,30(2):15-18.
[17] 楊萬霞,方升佐.青錢柳種子綜合處理過程中內源激素的動態變化[J].南京林業大學學報:自然科學版, 2008, 32(5):85-88.
[18] Kochanko V G, Gitzesik M, Chojnowsk M,et al.Effect of temperature, growth regulators and other chemicals onEchinacea purpurea(L) Moench seed germination and seedling survival[J].Seed Sci & Technol, 1998,26: 547 - 554.
[19] 李金克,金幼菊,陳華君,等.紅松種子層積催芽處理和萌發過程中內源GA和IAA含量的變化[J].河北林果研究, 1997,12(3):203-208.
[20] 高紅兵,昊榜華,孫振良.東北紅豆杉層積過程中內源生長素和脫落酸含量的變化[J].吉林林學院學報, 1998, 14(4):187-189.
[21] 黃 丹,許岳香,胡海波.美國桂花種子低溫層積過程中內源激素的動態變化[J].林業科技開發,2010,24(1):53-56.
[22] 余朝霞,黃雪群. GA對林木種子萌發的調控研究進展[J].浙江林業科技,2003,23(1):73-77.
[23] 洑香香,周曉東,劉紅娜.山茱萸種子休眠機理與解除方法初探[J].中南林業科技大學學報,2013,33(4):7-13.
[24] 王 寧,梅海軍,袁美麗,等.赤霉素浸種和變溫層積過程對冬青種子激素含量的影響[J].河南農業大學學報, 2010, 44(5)524-527.
[25] Wareing PF, Saunders PF. Hormones and dormancy[J]. Annu Rev Plant Physiol, 2003, 22:261-288.
[26] Bewley JD. Seed germination and dormancy[J]. Plant Cell, 1997,9:1055-1066.
[27] 王友鳳,馬祥慶.林木種子萌發的生理生態學機理研究進展[J].世界林業研究,2007,20(4):19-23.
[28] 管康林.樹木種子休眠性狀的研究綜述[J].浙江林學院學報,1986,3(2):99-109.
