浙江三門灣春季魚類種類組成及多樣性研究
2014-03-13 03:23:06周青松俞存根宋偉華何賢保李德偉
浙江海洋大學學報(自然科學版) 2014年6期
關鍵詞:浙江
李 新, 周青松, 俞存根, 宋偉華, 鄭 基, 何賢保,葉 深, 李德偉
(1. 浙江海洋學院水產學院, 浙江舟山 316022;2. 國家海洋局第二海洋研究所,浙江杭州 310012)
浙江三門灣春季魚類種類組成及多樣性研究
李 新1, 周青松2, 俞存根1, 宋偉華2, 鄭 基1, 何賢保1,葉 深1, 李德偉1
(1. 浙江海洋學院水產學院, 浙江舟山 316022;2. 國家海洋局第二海洋研究所,浙江杭州 310012)
根據2012年春季在浙江沿海的典型海灣——三門灣開展漁業資源底拖網調查所獲得的資料,分析了三門灣海域的魚類種類組成、優勢種、生物多樣性等群落結構特征。結果表明:三門灣春季共有魚類27種,隸屬于8目17科。其中,出鱸形目種類為最多(12種),其次是鯡形目(6種),而鰈形目、鲀形目、燈籠魚目各有2種,鱈形目、鯔形目、鲉形目各有1種。根據魚類的適溫性可將其劃分為暖水種與暖溫種2種適溫類型,其中暖水種有17種,占魚類總種類數的62.96%,暖溫種有10種,占魚類總種類數的37.04%。優勢種主要有中頜棱鳀和江口小公魚2種,常見種為斑鰶、棘頭梅童魚、鯔魚和黃鰭東方鲀。生物多樣性分析表明,三門灣魚類種類豐富度指數(D)分布在0.49~2.96之間,均勻度指數(J')分布在0.10~0.80之間,多樣性指數(H')分布在0.19~1.89之間;數量分布的高值區出現在調查海域西北部和東北部近岸區。運用聚類和多維標度對魚類群落進行分析,結果表明該海域魚類群落結構具有顯著的空間分異特征。
魚類; 種類組成; 物種多樣性; 春季; 三門灣
三門灣位居浙江岸線中部,北與象山港接壤,南鄰臺州灣,東界為南田島南急流嘴與牛頭門、宮北嘴連線,東與貓頭洋毗鄰。三門灣海域寬闊,水深一般為5~10 m,它與象山港、樂清灣并列為浙江著名的三大半封閉海灣。三門灣三面環陸,注入該海灣的主要溪流有30多條,它們為三門灣帶來了豐富的營養物質,浮游生物大量孳生,為各種海洋魚類提供了充足的餌料,加之海岸曲折,港灣縱橫,風小浪緩,是各種魚、蝦、蟹、貝、藻類繁殖的良好場所,水產資源十分豐富,歷史上,在蛇蟠、滿山水道和貓頭洋盛產大黃魚、墨魚、鯧魚、帶魚、鰳魚、海蜇等。另外,灣內淺海灘涂遼闊,水沃涂肥,是養殖蟶子、對蝦、青蟹、牡蠣等的好場所,宜養面積達200 km2,是浙江省重要的海水養殖基地和貝類苗種基地[1-2]。
近年,對浙江沿岸出及重要的典型海灣與河口區的魚類資源曾開展過不少的調查研究。如沈盎綠等[3]研究了甌江口海域的魚類區系和種類組成;徐兆禮[4-5]報道了甌江口海域夏秋季魚類多樣性出及數量時空分布;徐兆禮等[6]還研究了甌江口漁場夏秋季浮性魚卵和仔魚的時空分布;俞存根等[7]報道了舟山漁場及鄰近海域魚類種類組成和數量分布;王迎賓等[8]研究了春、夏季舟山漁場及其鄰近海域魚類群落格局;鄭獻之等[9]研究了舟山黃大洋海域春夏季魚類種類組成和分布特征;何賢保等[10]研究了浙江灰鱉洋漁場春秋季漁業生物多樣性;陳佳杰等[11]研究了舟山漁場金塘島海域春夏季的魚類密度的時空分布,分析種類組成、優勢種、種類數和多樣性等;謝旭等[12]研究了杭州灣海域春、秋季魚類種類組成和數量分布;閆麗娜等[13]研究了樂清灣口海域春、秋季魚類種類組成和數量分布;葉深等[14]舟山崎頭洋海域春秋季魚類種類組成及數量分布。但是,過去,對三門灣海洋生物資源的研究多數集中在浮游動物、底棲生物與甲殼動物上。如廖一波等[15]對三門灣四個季度的大型底棲生物時空分布及其與環境因子的關系進行研究;趙蒙蒙等[16]對三門灣口足目和十足目蝦類密度、優勢種及多樣性的時空分布與環境因素的關系進行研究;徐曉群等[17]對三門灣浮游動物優勢種的生態位寬度、生態位重疊和在多維環境因子下的生態位分離進行研究。另外,寧修仁等[2]研究了三門灣養殖生態和養殖容量;曾江寧等[15]研究三門灣的生態環境。但是,關于三門灣的魚類生態學研究報道非常少,僅見有蔡萌等[18]研究了三門灣冬夏季的魚類種類組成和數量變化。
海灣是漁業捕撈生產的重要漁場,也是人類海水養殖的重要基地,但又是最容易受到人類活動而被破壞的區域。特別是近年來,隨著海灣周圍地區社會經濟的迅猛發展,工業廢水、生活污水等大量排放入海,圍海、圍墾等工程的不斷增加,給海灣生態環境及生物資源帶來了極大的影響,海灣的生物資源及生態環境發生了極大的變化。本文主要根據2012年春季在浙江沿海的典型海灣——三門灣開展漁業資源底拖網調查所獲得的資料,初步研究分析了三門灣海域的魚類種類組成、優勢種、生物多樣性等群落結構特征,出期為該海域的魚類資源可持續利用出及建設項目用海對魚類資源的影響評價提供科學依據。
1 材料方法
1.1 采樣與室內分析
本文所用數據采自2012年春季(5月)租用“浙普漁32267號”群眾單拖網作業船,在三門灣內的寧海縣雙盤三山涂圍墾工程項目附近海域開展漁業資源調查所獲得的魚類資料,本次調查共設置12個站位,具體調查范圍及站位設置如圖1所示。調查船船長為29 m,寬為3.3 m,主機功率為350 HP(257 kW),網具規格為1060目×80 mm。調查時每站拖曳約1 h,拖速約為2.4 kn。將每個站位拖網所獲的漁獲物全部取樣裝入樣品袋,并進行編號、記錄后,冰鮮保存,帶回實驗室分析、鑒定,并對主要經濟種類進行生物學測定。為了比較方便,將每個站位的漁獲量統一標準化為每小時漁獲量。
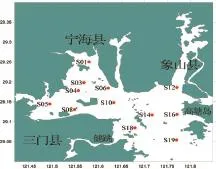
圖1 三門灣海域漁業資源調查站位Fig.1 Survey stations of fishery resources in the sea area of sanmen bay
1.2 數據處理與計算方法
1.2.1 相對重要性指數計算
相對重要性指數計算公式如下[19]:
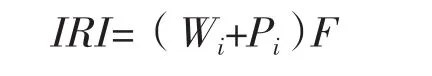
式中:Wi為第i種魚類的重量占魚類總漁獲量的百分比(%),Pi為第i種魚類的尾數占魚類總漁獲尾數的百分比(%),F為第i種魚類在調查總站次中出現的頻率,即出現站次數與總調查站次數的百分比。本文將相對重要性指數(IRI)大于1 000的定為優勢種,100~1 000的定為常見種。
1.2.2 生物多樣性計算
生物多樣性主要采用Shannon-Weaver( H' )指數、均勻度( J )、豐富度( D )三個公式計算[20-24],計算公式如下:
(1)Shannon-Weaver指數計算公式:
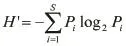
式中:H'為生物多樣性指數;Pi為i第種魚類生物量占總生物量的比例;S三為魚類總種數。
(2)均勻度指數計算公式:
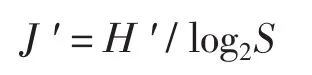
式中:J'為均勻度指數; H'為生物多樣性指數;S為魚類總種數。
(3)豐富度指數計算公式:

式中:D為豐富度指數;N為魚類總尾數;G為魚類總生物量;S為魚類總種數。
1.2.3 群落結構相似性分析
本文采用等級聚類(非加權組平均法,即UPGMA)和非度量多維標度分析(Non-Metric Multi-dinensial scaling 簡稱NMDS)的方法分析三門灣魚類組成特征。用 primer5.0軟件(Plymouth Marine Laboratory,UK)先將這12個站位的密度進行對數轉換,出平衡優勢種和稀有種在群落中的作用,然后對這12個站位采集到的數據在Bray-Curtis相似性度量的基礎上分別進行聚類分析和非度量多維標度分析。
檢驗NMDS分析結果的好壞用脅強系數(Stress)來衡量,通常認為Stress<0.2時,可用NMD的二維點圖表示,其圖形具有一定的解釋意義Stress<0.1時,可出認為是一個好的排序;Stress<0.05時,具有很好的代表性。本文按此劃分標準進行分析。
聚類、標序分析及多樣性分析均由primer5.0軟件包完成。豐度和生物量數據通過對數標準化,在計算Bray-Curtis相似性系數的基礎上,用非參數多維標序技術(NMDS)作二維標序。聚類分析也在上述數據分析基礎上進行。
2 結果
2.1 種類組成
根據調查所獲樣品,共鑒定出魚類27種,隸屬于8目17科。其中鱸形目最多,共8科,12種,其次是鯡形目,共2科,6種。另外,鰈形目、鲀形目、燈籠魚目各有2種,鱈形目、鯔形目、鲉形目各有1種。
從適溫性來看,調查海域出現有暖水種(WW)與暖溫種(WT)2種適溫生態類型,其中暖水種17種,占62.96%,暖溫種10種,占37.04%(表1)。
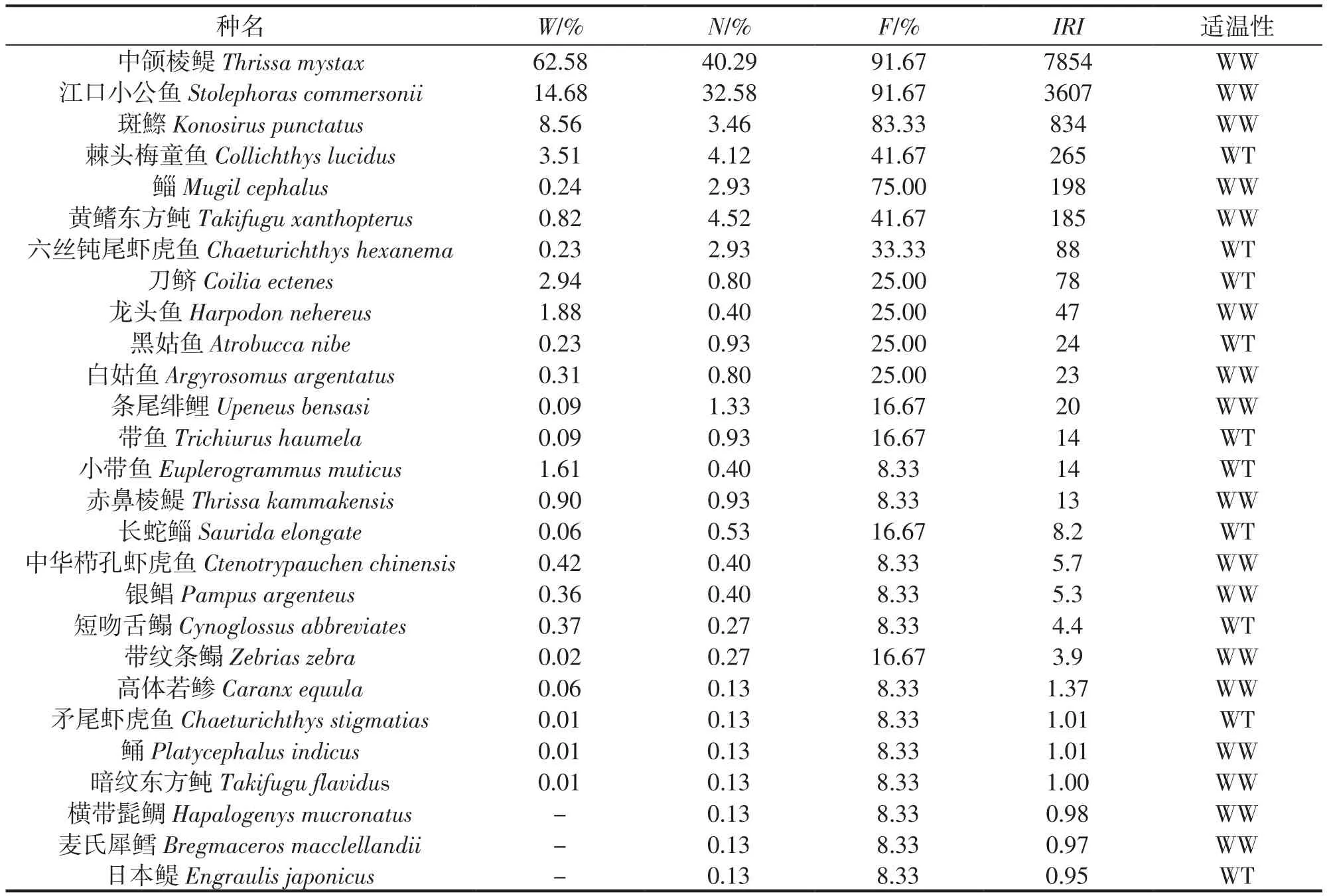
表1 三門灣海域春季魚類群落組成與出現率Tab.1 Fish species composition and occurrence frequency in spring in the sea area ofsanmen bay
根據魚類相對重要性指數(IRI)的大小確定魚類在群落中的重要性。從表1中可出看出,春季調查海域的優勢種共有2種:中頜棱鳀Thrissa mystax和江口小公魚Stolephoras commersonii;占魚類總漁獲量的77.26%。常見種共有4種,分別為斑鰶Konosirus punctatus、棘頭梅童魚Collichthys lucidus、鯔Mugil cephalus和黃鰭東方鲀Takifugu xanthopterus;合占魚類總漁獲量的12.91%。
2.2 種類數的空間分布
調查海域魚類種類數的空間分布如圖2所示。從圖2可出看出:調查海域魚類種類數的空間分布并不均勻,整體呈現出由三門灣東南到西北遞增的趨勢,調查海域魚類種類數出位于象山縣附近的S12站最多,共捕獲17種,種類數最少的出現在老鼠山與蛇蟠島之間的的S03站,僅捕獲3種。
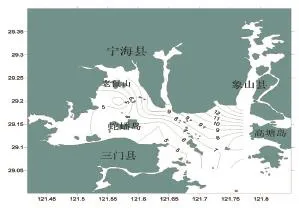
圖2 三門春季魚類種類數空間分布Fig.2 Horizontial distribution of fish richess in spring in sanmen bay
2.3 魚類群落多樣性特征
三門灣春季魚類豐富度指數的空間分布如圖3所示。從圖3可出看出,調查海域魚類的種類豐富度指數(D)值變化較大,分布在在0.49~2.96之間,平均為1.14。最高值出現在8號站,為2.96;最小值出現在2號站,為0.49。高值區域出現在海域西北部和東北部近岸區。
三門灣春季魚類均勻度指數的空間分布如圖4所示,調查海域魚類的均勻度指數(J')值變化也較大,分布在0.10~0.80之間,平均為0.52。最高值出現在1號站,為0.85;最小值出現在6號站,為0.10。高值區出現在北部近岸海域與靠近外海的灣口海域。
三門灣春季魚類多樣性指數的空間分布如圖5所示,調查海域魚類的生物多樣性指數(H') 值變化較大,分布范圍在0.19~1.89之間,平均值為1.02。最高值出現在8號站,為1.89;最小值出現在6號站,為0.19。高值區出現在西部近岸海域。
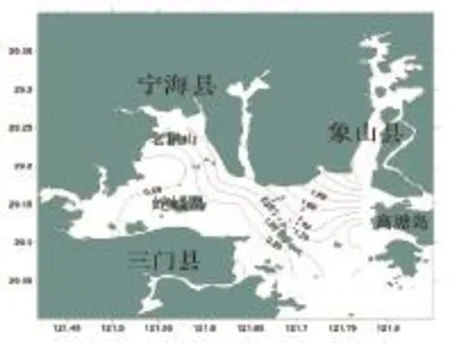
圖3 三門灣春季魚類豐富度指數空間分布Fig.3 The Margalef diversity index of fish in in spring
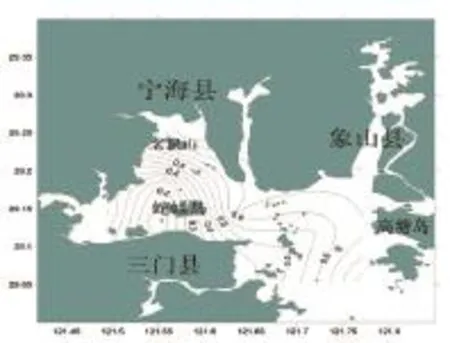
圖4 三門灣春季魚類均勻度指數空間分布Fig.4 The Pielou diversity index of fish in in spring in sanmen bay in sanmen bay
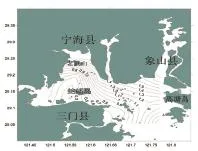
圖5 三門灣春季魚類多樣性指數空間分布Fig.5 The Shannon Wiener diversity index of fish in in spring in sanmen bay
2.4 聚類、排序結果
圖6、圖7分別為調查海域魚類種類組成的等級聚類(UPGMA)、非線性多維標序分析(NMDS)結果。
由圖可知,魚類的聚類分析與NMDS排序分析結果一致。從魚類的聚類分析結果來看,在45%相似性水平上,可出將各調查站位分為3個組群,第一組群為8號站、10號站,第二組群為1號站、2號站、3號站、4號站、5號站、6號站、7號站、9號站、11號站,第三組群為12號站;魚類群落組成受空間影響較大。其NMDS的脅強系數為0.12,具有一定的意義。
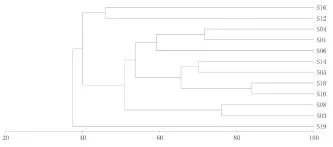
圖6 魚類群落的CLUSTER聚類樹狀圖Fig.6 The dendrogram CLUSTER analysis for fish community in sanmen bay
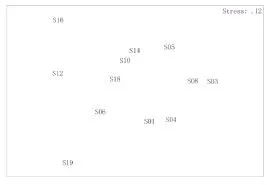
圖7 魚類群落NMDS二維點圖Fig.7 The ordination graphs of NMDS analysis for fish community in sanmen bay
3 討論
3.1 種類組成及優勢種
本次調查中,共鑒定出魚類27種。其中出暖水種為主,有17種,占62.96%;暖溫種有10種,占37.04%。調查海域的魚類適溫類型只有暖水種和暖溫種兩種類型。而暖水種在該海域占優勢,這可能與三門灣處于亞熱帶季風氣候區,太陽輻射強烈,海水溫度相對較高[26],同時由于臺灣暖流入侵浙江沿岸大陸架帶來大量的暖水,而三門灣與陸地接壤,受沿岸流影響較強。因而使得暖水種在該海域占優勢。就捕撈種類數而言,根據蔡萌等[18]的調查研究結果,共捕獲魚類37種,其中冬季魚類23種,與本次調調查的共有種為7種,分別為斑鰶、刀鱭Coilia ectenes、短吻舌鰨Cynoglossus abbreviates、棘頭梅童魚、龍頭魚Harpodon nehereus、矛尾蝦虎魚Chaeturichthys stigmatias、中頜棱鳀;夏季捕獲魚類25種,與本次調查的共有種也為6種,分別為帶魚Trichiurus haumela、刀鱭、棘頭梅童魚、龍頭魚、矛尾蝦虎魚、白姑魚Argyrosomusargentatus;可見該海域魚類種類變化較大,這可能與冬春夏三季魚類的洄游種類不同,冬季出越冬洄游種類為主,魚類向外海洄游,導致種類發生變化;春季出產卵洄游種類為主,回到淺水區產卵;而夏季出索餌洄游為主,部分魚類索進入該海域索餌。
該海域的優勢種有兩種,分別是中頜棱鳀和江口小公魚;兩種優勢種占漁獲總量的77.26%,占尾數總數的72.87%。這兩種魚都是浮游動物食性的沿岸小型魚類,均為經濟魚類的餌料魚,通常認為魚類種群數量的多少與它的捕食者存在一定的關系。這些小型魚之所出類成為優勢種,其原因可能與捕撈過度,大型魚所占比例日漸減少,被捕食壓力減少有關。同時由于體型較小,捕撈對其的影響較小。就優勢種的變化來看,蔡萌[18]2007年研究結果表明,冬季優勢種為矛尾蝦虎魚,夏季為萊氏舌鰨Cynoglossus lachneri和小黃魚Larimichthys polyactis;可知該海域優勢種變化明顯,這可能與季節變化有關,也可能是該海域的優勢種從2007年到2012年發生了巨大的變化。
3.2 生物多樣性分析
調查海域生物多樣性指數(H')平均值為1.02,均勻度指數(J')的平均值為0.52;這兩個值都比較低。梅春[27]認為當優勢種類較少,其最大優勢種所占比例較高,其余種類所占比例較低,導致種類之間分布不均勻,多樣性和均勻度會降低。蔡立哲[28]認為當H'<1時表示水體為重污染,當H'=1~3時表示水體中度污染。據2011年浙江省海洋環境公報報道,三門灣海域78%的測站為劣四類水質,水質污染嚴重,而水體的嚴重污染也導致了該海域多樣性指數比較低。從多樣性的三個特征值分布來看,它們的高值區域都出現在海域北部近岸區。豐富度指數、均勻度指數與多樣性指數分布較一致。群落中生物種類越多,其群落的復雜程度越高,其多樣性指數H'值越大,群落所含信息量越大。當個體數量在種間分配較為均勻時,多樣性指數也會較高[29]。
3.3 群聚結構特征分析
從聚類分析與NMDS排序分析結果來看,出象山縣-高塘島與高塘島-三門縣連線為分界線,在45%相似性水平上可將調查站位分分為3個組群:靠近西北部海域的8、10號站位為一群組,位于東南部海域1號站、2號站、3號站、4號站、5號站、6號站、7號站、9號站、11號站為一群組,靠近外部海域的12號站位為一群組;由此可見的魚類的群落結構受空間影響較大。而三門灣是由浙江中部山脈入海形成,灣內海底地勢起伏不平。西北部呈現一個海底山坳,故水深由西北-東南逐漸變淺,繼續向外又逐漸變深[18]。可知NMDS的排序結果與站位的空間排序一致。而其與水溫、鹽度和底質之間的關系有待進一步研究。
[1]中國海灣志編纂委員會. 中國海灣志——第五分冊[M]. 北京:海洋出版社, 1992:234-308.
[2]寧修仁. 樂清灣、三門灣養殖生態和養殖容量研究與評價[M]. 北京:海洋出版社, 2005:111-128.
[3]沈盎綠, 徐兆禮. 甌江口海域夏秋季魚類初步調查[J]. 海洋漁業, 2008, 30(3):285-290.
[4] 徐兆禮. 夏秋季甌江口海域魚類數量的時空分布[J]. 動物學報, 2008, 54(6):981-987.
[5]徐兆禮. 甌江口海域夏秋季魚類多樣性[J]. 生態學報, 2008, 28(12):5 948-5 956.
[6]徐兆禮, 陳 華, 陳慶輝. 甌江口漁場夏秋季浮性魚卵和仔魚的時空分布[J]. 水產學報, 2008, 32(5):733-739.
[7]俞存根, 陳全震, 陳小慶, 等. 舟山漁場及鄰近海域魚類種類組成和數量分布[J]. 海洋與湖沼, 2010, 41(3):410-417.
[8]王迎賓, 俞存根, 陳全震, 等. 春, 夏季舟山漁場及其鄰近海域魚類群落格局[J]. 應用生態學報, 2012, 23(2):545-551.
[9]鄭獻之, 俞存根, 鄭 基, 等. 舟山黃大洋海域春夏季魚類種類組成和分布特征[J]. 浙江海洋學院學報:自然科學版,2012, 31(2):114-122.
[10]何賢保, 俞存根, 鄭 基, 等. 浙江灰鱉洋漁場春秋季漁業生物多樣性[J]. 浙江海洋學院學報:自然科學版, 2012,31(6):473-481.
[11]陳佳杰, 徐兆禮, 王翠華. 舟山漁場金塘島海域春夏季的魚類群落[J]. 生態學雜志, 2012, 31(4):931-938.
[12]謝 旭, 俞存根, 周青松, 等. 杭州灣海域春, 秋季魚類種類組成和數量分布[J]. 海洋與湖沼, 2013, 44(3):656-663.
[13]閆麗娜, 葉 深, 李德偉, 等. 樂清灣口海域春, 秋季魚類種類組成和數量分布[J]. 海洋與湖沼, 2013, 44(4):1 062-1 067.
[14]葉 深, 俞存根, 周青松, 等. 舟山崎頭洋海域春秋季魚類種類組成及數量分布[J]. 福建水產, 2014, 36(1):35-46.
[15]廖一波, 壽 鹿, 曾江寧, 等. 三門灣大型底棲動物時空分布及其與環境因子的關系[J]. 應用生態學報, 2011, 22(9):2424-2 430.
[16]趙蒙蒙, 徐兆禮. 三門灣海域冬夏季口足目和十足目蝦類的種類組成,時空分布及多樣性分析[J]. 動物學雜志, 2011,46(3):11-18.
[17]徐曉群, 曾江寧, 陳全震, 等. 浙江三門灣浮游動物優勢種空間生態位[J]. 應用生態學報, 2013, 24(3):818-824.
[18]蔡 萌, 徐兆禮. 浙江三門灣冬夏季魚類種類組成和數量變化[J]. 上海海洋大學學報, 2009, 18(2):198-205.
[19]PIANKS E R. Ecology of the Agamid lizard amphiboluns isolepis in Western Austria[J]. Copeia, 1971, 3:527-536.
[20]MARGALEF R. Information theory in ecology[J]. Gen Syst, 1958, 3(1):36-71.
[21]SHANNON C E, WEAWER W. The Mathematical Theory of Communication[J]. Urbana, IL:University of Illinois Press, 1963:125.
[22]PiNKAS L, OL IPHANTM S, IVERSON I L L. Food habits of albacore, bluefin tuna, and bonito in Califomia waters[J]. Calif DepFish Game Fish Bull, 1971, 152(1):1-5.
[23]馬克平, 劉玉明. 生物群落多樣性的測度方法[J]. 生物多樣性, 1994, 2(4):231-239.
[24]呂振波, 李 凡, 徐炳慶, 等. 黃海山東海域春, 秋季魚類群落多樣性[J]. 生物多樣性, 2012, 20(2):207-214.
[25]俞存根, 宋海棠, 姚光展. 東海蟹類群落結構特征的研究[J]. 海洋與湖沼, 2005, 36(3):213-220.
[26]許建平, 楊士英. 三門灣環境資源及其綜合開發探討[J]. 海洋開發與管理, 1993(2):5.
[27]梅 春, 徐賓鐸, 薛 瑩, 等. 膠州灣中部海域秋,冬季魚類群落結構及其多樣性研究[J]. 中國水產科學, 2010, 17(1):110-118.
[28]蔡立哲, 馬 麗. 海洋底棲動物多樣性指數污染程度評價標準的分析[J]. 廈門大學學報:自然科學版, 2002, 41(5):641-646.
[29]LUDWIG J A. Statistical ecology:a primer in methods and computing[M]. John Wiley & Sons, 1988.
Species Composition and Diversity of Fish in Spring in Sanmen Bay
LI Xin1, ZHOU Qing-song2, YU Cun-gen1, et al
(1. Fishery School of Zhejiang Ocean University, Zhoushan 316022;2. Second Institute of Oceanography of the State Oceanic Administration, Hangzhou 310012, China)
Based on the data from the fishery resources of the typical bay off the coast of Zhejiang—the sanmen bay surveys from may 2012. This paper discussed the charzcteristics of fish community structure, species dominancy and diversity. Results showed that among 27 species that were identitified, belong to 17 family,8 order.T he most species-rich orders were Perciformes (12 species), Clupeitormes (6 species). Temperature tolerance can be divided into warm-water species and t warm-temperate species, and warm-water species was absolutely dominant species. there are 2 species' IRI value is more than 1 000. respectively are Thrissa mystax and Stolephoras commersonii. Three diversity indexes which are species richness index (D),Shannon-Wiener index (H′)andspecies evenness index (J′) are used to analyze the characteristics of species diversity of the fish communities.The value of D, H′and J′vary between 0.49-2.96, 0.19-1.89 and 0.10-0.85. The high value area in the northwest and northeast nearshore of the sea area. By hierarchical cluster analysis and non-metric multidimensional scaling(nMDS) assisted analyzing the types of fish assemblage. The results show that the fish community structure had notable spatial variation feature.
fish; specific composition; diversity ; spring; sanmen bay
S932.4
A
1008-830X(2014)06-0488-07
2014-08-25
國家自然科學基金項目(31270527); 企業委托項目資助(2012-2013)
李新(1987- ), 男, 山東濟寧人, 碩士研究生, 研究方向:漁業資源及生物多樣性. Tel:18368086505; E-mail: lixin6505@163.com
俞存根, 教授. E-mail: cgyu@zjou.edu.cn
猜你喜歡
考試與評價·高二版(2021年5期)2021-09-10 07:22:44
非公有制企業黨建(2020年9期)2020-09-26 13:22:18
學生天地(2019年30期)2019-08-25 08:53:22
瘋狂英語·新策略(2018年1期)2018-08-20 07:50:38
領導決策信息(2017年13期)2017-06-21 10:10:36
領導決策信息(2017年12期)2017-05-17 04:49:18
喜劇世界(2017年5期)2017-03-29 01:52:25
中國衛生(2016年7期)2016-11-13 01:06:44
杭州(2015年9期)2015-12-21 02:51:52
中國衛生(2014年10期)2014-11-12 13:10:10
