尿酸轉運蛋白是治療高尿酸血癥的新靶點
2014-03-16 01:47:32張冰清曾學軍
基礎醫學與臨床 2014年11期
關鍵詞:功能
張冰清,張 昀,曾學軍
(中國醫學科學院北京協和醫學院北京協和醫院普通內科,北京100730)
尿酸是人體內嘌呤代謝的終產物;近年來研究已證實高尿酸血癥與多種心血管事件相關。約90%的高尿酸血癥由尿酸排泄不足所致,且主要指腎臟排泄不足。腎臟排泄尿酸需多種位于腎小管上皮細胞的尿酸轉運蛋白共同協調完成,是腎臟調節體內尿酸穩態的關鍵,也是多種內源性和外源性物質的作用靶點。對這些轉運蛋白的深入了解,對臨床控制高尿酸血癥及其相關心血管事件具有重要意義。按照功能分類,尿酸轉運蛋白包括重吸收相關蛋白、排泄相關蛋白和骨架蛋白(表1)。
1 重吸收相關蛋白
1.1 尿酸轉運子1(urate transporter 1,URAT1)
URAT1 屬有機陰離子轉運蛋白(organic anion transporter,OAT)家族,編碼基因為SLC22A12。URAT1 含12 個跨膜區域,主要表達于腎臟近曲小管上皮細胞的管腔側膜,少量表達于血管平滑肌細胞和脂肪細胞[1]。
功能研究顯示,URAT1 為尿酸-陰離子交換轉運蛋白,介導細胞外尿酸向細胞內轉運。在腎小管上皮細胞,URAT1 介導尿酸從管腔內攝取,是尿酸重吸收的第一步[2]。SLC22A12 基因敲除小鼠的尿尿酸排泄顯著增加[3]。在人體,URAT1 失功能性突變可導致尿尿酸排泄率增高,血漿尿酸水平降低[1]。
在高脂血癥小鼠中,URAT1 的mRNA 和/或蛋白表達升高[4],提示URAT1 可能為高脂血癥與高尿酸血癥之間紐帶之一。在SLC22A12 上游存在一段雄激素應答片段,動物實驗證實雄激素可上調URAT1 mRNA[5];而雌激素可在翻譯后水平降低URAT1 的蛋白表達,影響尿酸的重吸收[6],提示URAT1 可能是不同性別間尿酸差異的原因之一。
促尿酸排泄劑苯溴馬隆可結合并抑制URAT1,減少尿酸的重吸收;當URAT1 突變時,苯溴馬隆無促尿酸排泄作用[7]。氯沙坦不僅可抑制URAT1 的活性,減少尿酸的重吸收,還可降低URAT1 mRNA 的量[8]。而吡嗪酰胺、呋塞米、內酰胺類抗生素等,則可增強URAT1 重吸收尿酸的功能[7]。對URAT1 結構和功能的深入理解,為新型降尿酸藥物的研發提供了新的靶點[9]:新型URAT1 抑制劑Lesinurad 目前正在進行Ⅲ期臨床試驗(clinicaltrials.gov NCT01493531,NCT0150158,NCT01510769,NCT01508702),而Arhalofenate 和tranilast 目前正在Ⅱ期臨床試驗中[10]。
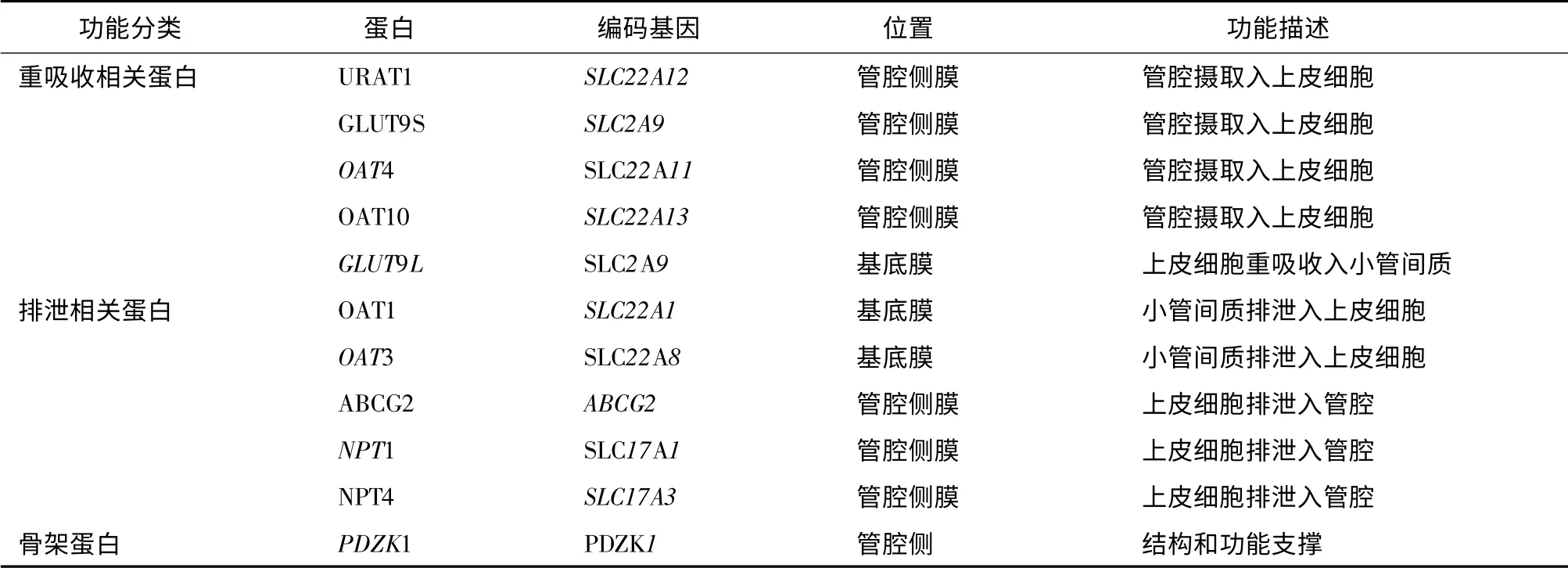
表1 各種尿酸轉運蛋白的分布部位和作用Table 1 Distribution and functions of urate transporters
1.2 葡萄糖轉運蛋白9(glucose transporter 9,GLUT9)
GLUT9 屬于葡萄糖轉運蛋白家族,編碼基因為SLC2A9。GLUT9 有兩種剪切變異體:GLUT9L(長型)和GLUT9S(短型)。兩種變異體均可表達于胎盤和腎臟近曲小管,但GLUT9L 位于基底膜側,GLUT9S 則位于管腔側膜[11]。
功能研究顯示:GLUT9 是一個高效的電位依賴型單轉運蛋白。GLUT9S 與URAT1 的功能相似;而GLUT9L 是唯一位于腎小管上皮細胞基底膜側的重吸收相關蛋白,是尿酸重吸收的最后步驟,細胞研究顯示當GLUT9L 突變時,尿酸重吸收幾乎完全停止[2]。在動物水平,SLC2A9 轉基因小鼠的尿尿酸排泄顯著降低,血尿酸水平輕度增高[12]。在人類,GLUT9 蛋白失功能性突變可導致尿酸排泄率增高,血漿尿酸水平降低[13]。
相比URAT1,GLUT9 受外源性藥物的影響較小。但由于GLUT9 在尿酸重吸收過程中的關鍵作用,針對此靶點的藥物將具有強大的降尿酸作用。同時GLUT9L 還可表達于軟骨細胞,這類新型藥物還可能對痛風相關骨侵蝕具有保護作用[14]。
2 尿酸分泌相關蛋白
2.1 ATP 結合盒蛋白家族G 蛋白亞家族成員2(ATP-binding cassette family,G subfamily,type 2,ABCG2)
ABCG2 主要表達于胎盤、肝臟、小腸和腎臟近曲小管,可介導多種物質向細胞外的轉運。在腎臟和腸道,ABCG2 均位于上皮細胞的管腔側膜,與尿酸排泄有關。功能研究顯示ABCG2 陽性細胞內尿酸蓄積顯著降低,蛋白突變或被抑制時細胞內尿酸蓄積明顯增高[15]。動物實驗證實ABCG2 突變小鼠的血尿酸水平增高[16],在人類,ABCG2 突變被認為是痛風和高尿酸血癥的重要原因,特別是家族性痛風和青年高尿酸血癥的主要原因[17]。
研究顯示:ABCG2 突變不僅影響腎臟排泄尿酸的能力,還導致經腸道排泄的尿酸減少,繼而使得腎臟的尿酸負荷增大[18]。因此,以其作為藥物靶點,可避免腎小管功能受損以及泌尿系結石的形成。Woodward 等人[19]最近發現了一種分子伴侶,可糾正ABCG2 突變蛋白的構象并恢復其功能,有望成為新型降尿酸藥物。
3 其他尿酸轉運蛋白
除上述3 個最重要的尿酸轉運蛋白外,腎臟近曲小管上皮細胞還存在其他具有尿酸轉運功能的蛋白,例如有機陰離子轉運蛋白OAT4、OAT10 和磷酸鹽轉運子NPT1,NPT4 等(表1)。盡管這些蛋白轉運尿酸的活性較弱,但這些蛋白可能是某些藥物性高尿酸血癥的致病位點[14]。
4 骨架蛋白
PDZK1 是一種骨架蛋白,在腎臟近曲小管上皮細胞的管腔側胞漿內,與多種尿酸轉運蛋白相結合,是維持這些蛋白的正常構象和功能的關鍵[14]。PDZK1 上游存在過氧化物酶體增生物激活受體α(proliferators activated receptor-α,PPARα)結合區,貝特類藥物為PPARα 激動劑,可增強PDZK1 的表達,但其對尿酸的影響尚有待研究[20]。
5 小結
腎小管上皮細胞存在多種尿酸轉運蛋白,對這些蛋白的表達與功能的研究,極大的幫助新型降尿酸藥物的開發。同時,這些尿酸轉運蛋白在定位、結構和功能上相互協同,形成“尿酸轉運蛋白復合體”,提示需從整體的角度評估和研究尿酸的轉運。尿酸轉運蛋白是高尿酸血癥與其他代謝紊亂的聯系紐帶,對該領域的研究具有深遠的基礎和臨床意義。
[1]Tasic V,Hynes A M,Kitamura K,et al.Clinical and functional characterization of URAT1 variants[J].PLoS One,2011,6:e28641.
[2]Nakanishi T,Ohya K,Shimada S,et al.Functional cooperation of URAT1 (SLC22A12)and URATv1 (SLC2A9)in renal reabsorption of urate[J].Nephrol Dial Transplant,2013,28:603-611.
[3]Eraly S A,Vallon V,Rieg T,et al.Multiple organic anion transporters contribute to net renal excretion of uric acid[J].Physiol Genomics,2008,33:180-192.
[4]Doshi M,Takiue Y,Saito H,et al.The increased protein level of URAT1 was observed in obesity/metabolic syndrome model mice[J].Nucleosides Nucleotides Nucleic Acids,2011,30:1290-1294.
[5]Hosoyamada M,Takiue Y,Shibasaki T,et al.The effect of testosterone upon the urate reabsorptive transport system in mouse kidney[J].Nucleosides Nucleotides Nucleic Acids,2010,29:574-579.
[6]Takiue Y,Hosoyamada M,Kimura M,et al.The effect of female hormones upon urate transport systems in the mouse kidney[J].Nucleosides Nucleotides Nucleic Acids,2011,30:113-119.
[7]Shin H J,Takeda M,Enomoto A,et al.Interactions of urate transporter URAT1 in human kidney with uricosuric drugs[J].Nephrology (Carlton),2011,16:156-162.
[8]Nindita Y,Hamada T,Bahrudin U,et al.Effect of losartan and benzbromarone on the level of human urate transporter 1 mRNA[J].Drug Res (Stuttg),2014.doi:10.1055/s-0033-1363730.
[9]Wempe M F,Jutabha P,Quade B,et al.Developing potent human uric acid transporter 1 (hURAT1)inhibitors[J].J Med Chem,2011,54:2701-2713.
[10]Richette P,Garay R.Novel drug discovery strategies for gout[J].Expert Opin Drug Discov,2013,8:183-189.
[11]Witkowska K,Smith K M,Yao S Y,et al.Human SLC2A9a and SLC2A9b isoforms mediate electrogenic transport of urate with different characteristics in the presence of hexoses[J].Am J Physiol Renal Physiol,2012,303:F527-F539.
[12]Kimura T,Amonpatumrat S,Tsukada A,et al.Increased expression of SLC2A9 decreases urate excretion from the kidney[J].Nucleosides Nucleotides Nucleic Acids,2011,30:1295-1301.
[13]Kawamura Y,Matsuo H,Chiba T,et al.Pathogenic GLUT9 mutations causing renal hypouricemia type 2(RHUC2)[J].Nucleosides Nucleotides Nucleic Acids,2011,30:1105-1111.
[14]Bobulescu IA,Moe OW.Renal transport of uric acid:evolving concepts and uncertainties[J].Adv Chronic Kidney Dis,2012,19:358-371.
[15]Nakayama A,Matsuo H,Takada T,et al.ABCG2 is a high-capacity urate transporter and its genetic impairment increases serum uric acid levels in humans[J].Nucleosides Nucleotides Nucleic Acids,2011,30:1091-1097.
[16]Ichida K,Matsuo H,Takada T,et al.Decreased extrarenal urate excretion is a common cause of hyperuricemia[J].Nat Commun,2012,3:764.doi:10.1038/ncomms 1756.
[17]Matsuo H,Ichida K,Takada T,et al.Common dysfunctional variants in ABCG2 are a major cause of early-onset gout[J].Sci Rep,2013,3:2014.doi:10.1038/srep 02014.
[18]Matsuo H,Nakayama A,Sakiyama M,et al.ABCG2 dysfunction causes hyperuricemia due to both renal urate underexcretion and renal urate overload[J].Sci Rep,2014,4:3755.doi:10.1038/srep 03755.
[19]Woodward O M,Tukaye D N,Cui J,et al.Gout-causing Q141K mutation in ABCG2 leads to instability of the nucleotide-binding domain and can be corrected with small molecules[J].Proc Natl Acad Sci U S A,2013,110:5223-5228.
[20]Tachibana K,Anzai N,Ueda C,et al.Regulation of the human PDZK1 expression by peroxisome proliferator-activated receptor alpha[J].FEBS Lett,2008,582:3884-3888.
猜你喜歡
鐘表(2023年5期)2023-10-27 04:20:44
中華詩詞(2022年6期)2022-12-31 06:41:24
當代陜西(2021年21期)2022-01-19 02:00:26
中學生數理化(高中版.高考數學)(2020年1期)2020-02-20 13:23:44
經濟技術協作信息(2018年11期)2019-01-14 03:07:20
中國科技論壇(2017年7期)2017-07-25 08:49:53
制造技術與機床(2017年3期)2017-06-23 08:11:33
媽媽寶寶(2017年2期)2017-02-21 01:21:24
國際漢語學報(2016年1期)2017-01-20 08:21:20
中國中醫藥現代遠程教育(2014年22期)2014-03-01 04:32:55
