atp1基因在小麥BNS雄性不育系和自身轉換系中的差異表達
2014-04-29 10:18:06趙大華孫慧慧于同英李友勇
生命科學研究 2014年6期
趙大華 孫慧慧 于同英 李友勇
摘 要:atp1基因在植物中是線粒體基因組編碼,其產物ATPl是線粒體ATP合酶F1的α亞基,在小麥BNS的雄性不育系和它的轉換系中差異表達。為了檢測該基因的表達豐度,探討與BNS不育性的聯系,以BNS不育系和它的轉換系的幼穗、花藥和穗軸等組織為材料,利用實時熒光定量PCR方法,測定和比較該基因在不同組織中的表達水平,結果表明,該基因在各組織中的總表達量比內參甘油醛-3-磷酸脫氫酶基因表達量高3—4個數量級;在幼穗及各穗軸營養組織中表達量一致;花藥與營養組織比較,在不育系四分體和單核期花藥中表達均上調:在不育系中,該基因表達量在單核期花藥中顯著下調。 這些結果說明.線粒體atp1基因在小麥中是一個高水平表達基因.在BNS營養組織中組成型表達,在花藥中特異性上調表達,在不育系中表達受到抑制,表現顯著下調.表明atpl基因的下調表達與BNS不育性有關。
關鍵詞:小麥:BNS雄性不育;atp1;基因差異表達分析;熒光實時定量PCR
圖分類號:Q785
文獻標識碼:A
文章編號:1007-7847(2014)06-0488-06
ATP合酶(ATP synthase)是高等植物細胞內最人的酶系之一,定位在細胞質基質、葉綠體、線粒體等細胞器的內膜上。ATP合酶由兩部分組成,即F0和F1,當F0和F1在結合狀態時表現ATP合成酶的活性,在分離狀態下是ATP水解酶活性。線粒體ATP合酶的FI有9個亞基,Fo的亞基數不同物種數量不同。線粒體F1中的α亞基有3個分子,與β亞基3個分子交替排列,旋轉時合成ATP。研究已經明確,線粒體F1α亞基基因atp1位于線粒體基因組上,產物是α亞基蛋白ATPl,其表達是組成型的,但也受核基因調控。
植物雄性不育是植物的雌蕊發育止常,雄蕊發育不正常,不產生有功能花粉的一種現象。具有該特性的不育系是作物雜種優勢利用的核心材料。對不育機制進行研究,多數結果都顯示,不育系中與能量供應的相關代謝途徑和組成成分出現異常,表現ATP/ADP比率減少、ATP合酶活性降低、不育和可育植株之間ATPI差異表達、以及atpl基因差異表達等。
BNS (Bai-nong slerility,BNS)是一種新發現的對溫度敏感的小麥雄性不育系,有良好的不育性和轉換性,在雜交小麥利用中有重要價值。在前期的BNS蛋白質組學研究中,發現該不育系和轉換后的可育系的四分體到二核期花藥中,線粒體ATP合酶α亞基蛋白ATP1差異表達,在不育系中表達下調,并認為ATP1的數量下調,將導致ATP合酶組裝量減少,結果會使ATP合酶總體活性下降、ATP產量降低,能量供應不足。細胞內基因表達水平和它的產物蛋白質的表達水平并不都是一致的,但atp1基因在BNS的花藥和組縱中的表達水平尚未被檢測。為了進一步探時該基因在BNS中的表達特性,本文以BNS的花藥以及幼穗和穗軸等組織為材料,利用實時熒光定量PCR (Real time quafititative PCR,qRT-PCR)方法,檢測和比較該基因在不育系以及它的轉換后的可育系小孢子形成前后,也是BNS敗育的關鍵時期,在這些組織中的表達水平,揭示atpl基兇對BNS雄性不育發生的影響和作用。
1 材料與方法
1.1 BNS的種植和取材
BNS是本實驗室培育并套袋自交保存的材料,在河南輝縣小麥實驗基地種植,自 10 月 1日到11月18口,每8d播種l期,共播7期。 1)不育系取材:參考蘇晴等的方法,在不育播期(10月1日和10月9 日)的材料進入雌雄蕊分化期后取幼穗材料;到四分體期和單核期后,每天上午7時左右從田間取穗,實驗室顯微鏡檢查花粉發育時期正確,在15 ℃以下室溫環境,冰塊上剝取中上部小穗第一和第二小花的6枚花藥,并及時收取;同時取無小穗的穗軸為營養組織材料;2)轉換系和轉換后的可育系取材:10 月 l7 日播種的材料花粉發育開始轉換,可育花粉比例逐漸上升。轉換系取10 月 25 日播種的材料,可育系取11月18日播種的材料,取材的小孢子發育時期與不育系相同;3)F1的取材:F1組合為BNA×中國春,10月8日播種,是BNS的不育期,中國春可恢復BNS的育性,因此F1是可育的。
取下的幼穗、花藥、穗軸即時在液氮中冷凍1~2 min,取出放入-80℃冰箱巾保存。共取l0個樣品,10個樣品的編號為wh-l~wh-10,對應的發育時期分別為:不育系藥隔期幼穗、不育系叫分體花藥、不育系四分體穗軸、不育系單核期化藥、F1四分體期花藥、F1四分體期穗軸、轉換系單核期花藥、轉換系單核期穗軸、可育系單核期花約和可育系單核期穗軸。
1.2 qRT-PCR方法
1.2.1 引物
用蘇晴在BNS花藥中檢測設汁的引物,序列為:primer F 3'-TTCCGCGATAATGGAATG-CACGCA-5';Drirner R 3'-_AGCTACCTGCACCT-GTCTGGTCCC -5',由上海生工生物技術有限公司合成。
1.2.2 主要試劑與儀器
實驗所用試劑主要有RNA提取試劑盒(In-vitrogen,USA)、Promega公司(北京)的AMV逆轉錄酶系統、中國大連定顯PCR酶TaKaRa TaqTMHot Start Version(日本Takara公司)、Roche公司(瑞士)的Green I Master。實驗所用主要儀器有:羅氏480熒光實時定量PCR儀、2720 thermal cycler(ABI,USA) PCR擴增儀、和Nanodrop ND-1000 Spectrophotometer (Nan(Jdrop technologies, USA)檢測儀等。
1.2.3 總RNA提取
材料總RNA提取采用Trizol方法,見蘇睛,50 mg樣品液氮研磨至粉末,1 mL Trizol總RNA抽提液提取.預冷的75%乙醇洗滌RNA,真空冷凍干燥。干燥的RNA提取物溶于20~40 μL DEPC水,取1.5μL紫外分光光度計檢測其濃度和A260/A280的比值:根據濃度取0.5μg RNA用1%瓊脂糖做純度檢測,條件是:0.5xTBE,4 V/cm,45 min。
1.2.4 cDNA第一鏈合成
根據Promega公司的操作說明書,1μL OligodT (0.5 μg/μL)和2.0 μg Total RNA加入PCR管,補充DFPC-H20至9μL。混勻后離心,70℃溫浴10 min,在0 ℃冰浴中與模板退火。之后,用5xRT buffer 4μL、10 mmol/L dNTPs 2μL、RNasin0.5 μL、AMV-Rtase lμL、DEPC H20 3.5 μL,混勻后42 ℃水浴2min,再加入M-MLV逆轉錄酶2 μL, 42℃孵育l h完成RT,70 ℃處理10 min使RT酶失活,得到的RT反應產物cDNA,可立即用于PCR,或保存在-80 ℃下備用。
1.2.5 目標基因熒光定量
使用熒光標記PCR方法,熒光染料為SYBRCreen I,羅氏480熒光實時定量PCR儀,3次重復,采用20 μL反應體系:模板1μL,上下游引物各0.5μL, Q-PCR Enhancer 5μL,2xSYBRSYBR Green I 10 μL, DEPC-H20 3μL。PCR反應程序采用二步法:95℃預變性5 min,95℃變性5 s,60℃退火30 s,45個循環,每次在延伸階段讀取吸光值;在PCR反應完成后做熔解曲線分析,方法是在95℃變性l min,冷卻至65 ℃,然后從65℃到95℃,每秒增加0.1℃,同時讀取吸光值。
1.2.6 計算方法
qRT-PCR結果的比較有不同層次,本實驗數據處理參考Jiang等的計算方法,在△Ct和AACt水平比較,△△Ct≥l,相對豐度的倍數2-△△Ct≥2,即達差異顯著水平。
2 結果與分析
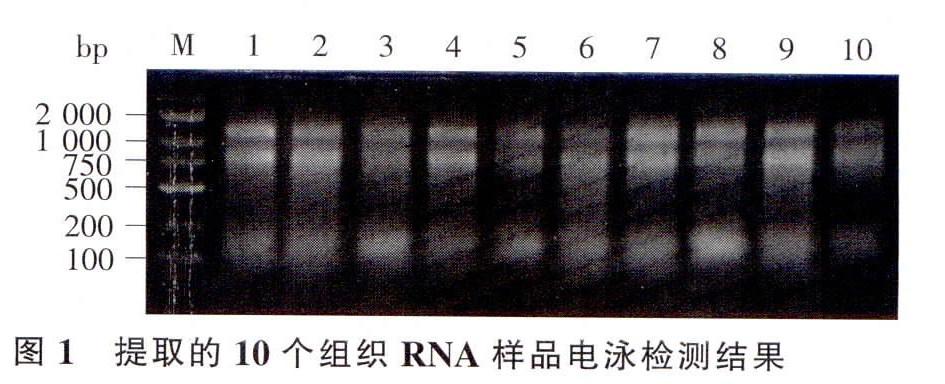
2.1 樣品RNA質量檢測
Trizol方法提取的花藥總RNA,50 mg樣品提取后定溶到30 μL體積,電泳檢測結果見圖l:OD260/OD20比值檢測結果,穗軸組織為1.79~1.84,花藥組織為1.83—1.94。兩項檢測結果均顯示RNA質量符合要求。
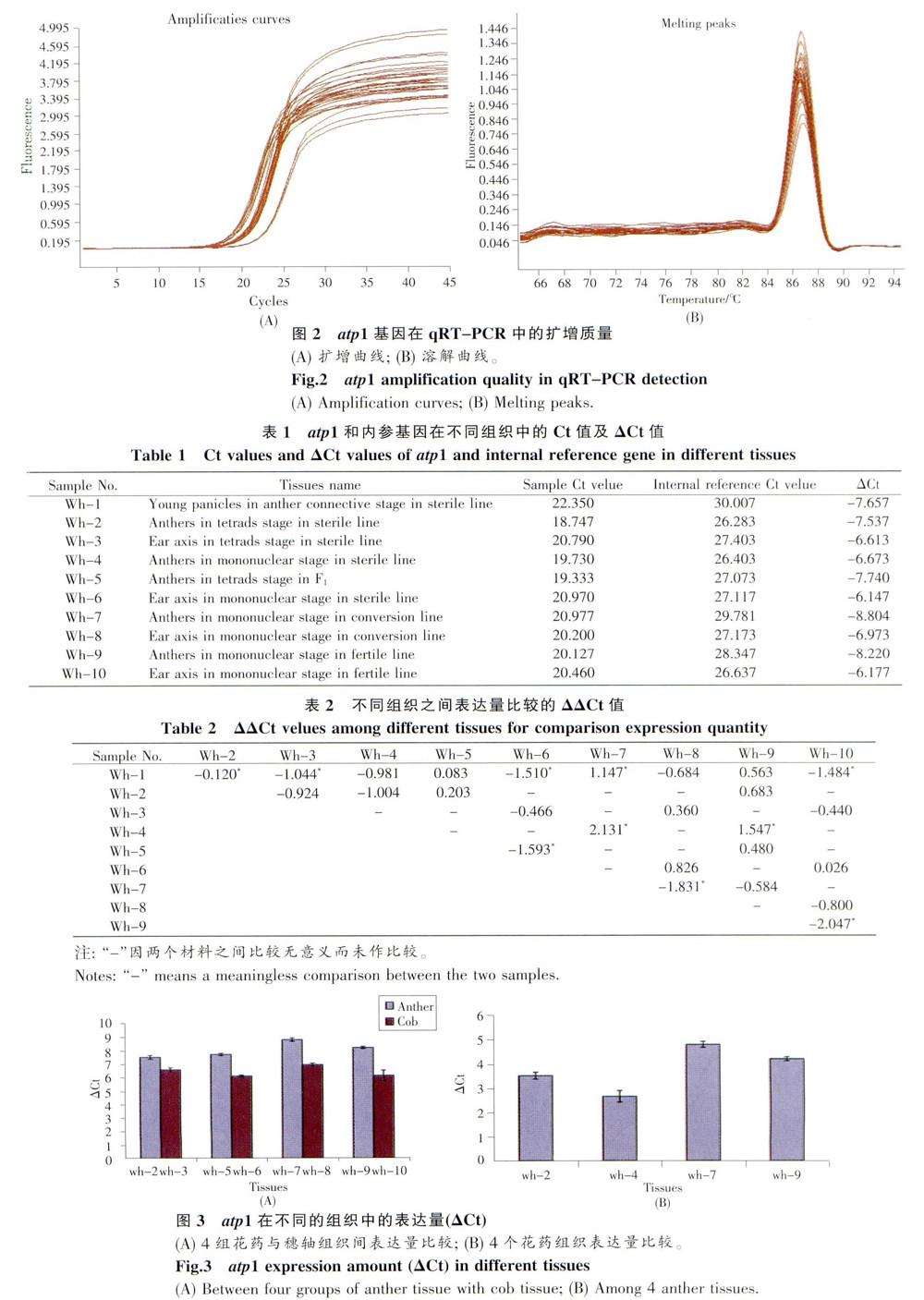
2.2 實時定量擴增結果
熒光染料標記的PCR方法,預試的擴增曲線和溶解曲線見圖2,顯示擴增的3個階段典型,擴增產物單一。內參基因為甘油醛-3-磷酸脫氫酶(GAPDH, glyceraldehyde -3 -phosphate dehydroge-nase)基因,獲得的樣品ACt、內參△Ct見表l,各樣品間△△Ct見表2,部分樣品間AACt的比較見圖3。
2.3 不同組織間的表達量比較
從表1、2和圖2可看出,atp1基因是一個高表達水平基因,相對表達量比內參基因甘油醛一3-磷酸脫氫酶高4個以上△Ct數量級,在BNS的10個組織中的表達模式有如下特點:1)該基因在不育系、可育系和F1的4個穗軸組織中,即,不育系四分體穗軸、F1四分體期穗軸、轉換系單核期穗軸和可育系單核期穗軸,它們的△Ct值相近,分別是-6.613、-6.147、-6.973和-6.177,相互之間的△△Ct值均小于l(見表2),說明該基因在營養組織中組成型表達;2)該基因在4組花藥組織和穗軸組織中的表達,均顯示花藥中表達水平高于穗軸組織,見圖3A,其中3組(wh-5/wh-6、wh-7/wh -8、wh -9/wh -10) △△Ct值大于l,分別為一1.593、-1.831、-2.047,差異達顯著水平,wh -2/wh一3為-0.923,接近1,表說明atp1基因在花藥中特異性上調表達;3)該基因在4個花藥組織之間的表達水平比較,顯示不育四分體期花藥中與藥隔期穗組織中表達一致,仍在較高水平,但到單核期,不育系花藥中表達下調,而在轉換系和可育系的花藥中,基因表達保持較高水平(圖3B),單核期的下調數量,與不育系四分體期、轉換系單核期和可育系單核期的表達量之間的△△Ct值絕對值均大于l,達顯著水平(見表2),表明該基因在不育系單核期的下調表達與BNS的不育性相關:4)樣品Wh-l是不育期BNS減數分裂前的幼穗組織,該樣品的基因表達量與5個花藥樣品表達量結果比較,4個不顯著,但與4個穗軸的ACt比較.3個顯著.說明藥隔期幼穗的基因表達特性與花藥接近。幼穗組織雖屬營養組織,但它是穗營養組織、花藥營養組織、孢母細胞生殖組織等的混合體,已經觀察到BNS在該時期對溫度敏感,說明atp1基因在雌雄蕊分化時期時已開始差異表達。
從這些結果可以看出,atp1基因在BNS不育系、轉換后的可育系,以及與中國春雜交的F1的穗各組織中總體上是組成型表達;在不育播期條件下,分化的幼穗組織,包括四分體期的花藥,表達量處在較高水平,但到單核期表達量顯著下調;在轉換播期和可育播期的條件下,這些組織的該基因表達量保持在較高水平。
3 討論
植物雄性不育體系是雜種優勢利用的關鍵技術.探討其不育機制,對認識不育的特征特性和創造多遺傳背景不育類型等有重要指導作用。對水稻、煙草、油菜等作物雄性不育的研究,曾發現花藥能量供應短缺,ATP合酶α亞基蛋白及它的基因差異表達,因此認為atpl是一個重要雄性不育相關基因,本文檢測到該基因在BNS不育系小孢子單核期表達量顯著下調,說明該基因的表達與小麥的雄性不育性存在聯系。
3.1 atpl在BNS可育系穗組織中組成型表達,花藥中上調表達,不育系花藥中顯著下調
atp1基因在植物雄性不育中的表達模式,過去的研究有不同結果。Bergman在煙草中檢測atpl基因位于雄性不育基因orf274下游,并共轉錄,對ATP的合成影響顯著,但基因和蛋白質表達水平沒有檢測出差異;黃思齊在紅麻中檢測到atp1基因序列在不育系和保持系中完全相同,但在不育系的葉片和花藥中表達量下降;Wenliang等檢測了油菜細胞質雄性不育中10個線粒體基因表達譜.除atp6差異表達外,其他9個,包括atpl,表達差異不顯著,宋國琦等在小麥YS雄性不育A3017中利用cDNA-AFLP方法分離出atpl基因。可以看出,這些研究結果中,一些顯示atp1基因的表達是組成型的,一些結果是差異表達的。本實驗結果顯示atp1基因在小麥BNS各營養組織中表達量一致,符合組成型表達的基本模式,但在可育花藥中表達量比在穗軸等組織中表達顯著上調,說明該基因表達在花藥中具有上調表達特異性。在不育系單核期榆測到表達量顯著下調,說明BNS的不育性與atp1基因的缺陷表達存在聯系,這種缺陷表達,基因表達下調模式與蛋白質組學檢測的蛋白質下調表達模式結果一致。
3.2
atpl是BNS的重要不育相關基因
BNS是一個對溫度敏感的雄性不育系,當花藥發育過程中氣溫升高,花粉育性可白行恢復,這說明BNS花粉發育的一些重要功能基囚,包括atp1基因,其基因本質沒有改變,改變的是在不同的發育條件下,一些基因功能發生改變,發育條件恢復,基因功能恢復。因此atpl基因不是不育基因,而應是不育基因調控下的一個重要相關基因。推測該基因,或該基因的產物,是上游不育基因及產物的重要受體,以一種尚未知的方式影響著花粉的發育。雖然這個影響過程未知,但其結果是易被理解的,ATP在生物體中的重要地位,足以影響到花粉代謝的全部過程。因此,atp1的差異表達,以及前期研究的ATP1蛋白的缺陷表達,可直接導致ATP合成酶的組裝,ATP合成酶數量減少,將直接導致能量生成減少和供應短缺,導致花粉發育敗育。
3.3 atp1基因在單核期表達下調有發育生物學依據
檢測結果中發現atpl基因在不育四分體期花藥中表達量仍較高,到單核期花藥表達下調,該結果有花粉發育進程原因。atp1基因是線粒體基因組編碼,線粒體基因組基因表達受線粒體本身和核基因共同調控。小孢子是一個配子體世代,花粉內核基因和質基因表達是一個相對獨立的過程,這個過程應從小孢子游離后,即單核期開始。因此,四分體后,小孢子開始獨立發育,花粉發育基因表達程序性啟動。不育核基因對質基因的調控一般是抑制性的,因此atp1基因表達受到核不育基因抑制而下調。該結果對進一步探討不育基因對靶基因作用有重要意義。
