施氮對(duì)不同有機(jī)碳水平桉樹林土壤微生物群落結(jié)構(gòu)和功能的影響
2014-05-08 03:37:14蘇丹張凱陳法霖李睿達(dá)鄭華
生態(tài)環(huán)境學(xué)報(bào) 2014年3期
關(guān)鍵詞:水平
蘇丹,張凱,陳法霖,李睿達(dá),鄭華
中國科學(xué)院生態(tài)環(huán)境研究中心城市與區(qū)域生態(tài)國家重點(diǎn)實(shí)驗(yàn)室,北京100085
土壤微生物是土壤中營養(yǎng)周轉(zhuǎn)的主要參與者,對(duì)碳氮變化較為敏感。施氮及氮素的添加都可能影響土壤中微生物群落的變化,進(jìn)而影響微生物群落功能和土壤營養(yǎng)過程。圍繞氮素施用對(duì)土壤微生物群落的影響已經(jīng)開展了大量研究(Cusack等,2011;Waldrop等,2004;Zhong等,2010),并發(fā)現(xiàn)施氮可以改變土壤中主導(dǎo)微群落,由真菌主導(dǎo)的微生物群落轉(zhuǎn)化為細(xì)菌主導(dǎo)。Frey等施氮研究發(fā)現(xiàn),真菌微生物量隨著氮的增加而降低,而細(xì)菌變化并不顯著(Frey等,2004)。微生物群落的變化——真菌和細(xì)菌比例等變化,可能導(dǎo)致參與木質(zhì)素降解的真菌功能群豐度及土壤中酚氧化酶等酶活性降低,而改變土壤中有機(jī)物的代謝(Osono, 2007)。
土壤微生物群落同樣受到土壤中有機(jī)碳含量的影響,有機(jī)碳含量直接影響土壤微生物的生長(zhǎng),而影響土壤微生物群落豐度。其中有機(jī)質(zhì)高的土壤中微生物量往往高于有機(jī)質(zhì)低的土壤(Burger和Jackson,2003)。此外土壤有機(jī)物的組成中碳氮比影響土壤中微生物活性Hobbie, 1996),而影響微生物群落功能,如通過改變土壤酶活,影響土壤碳形態(tài),最終影響土壤物質(zhì)代謝。然而,在研究施氮水平對(duì)土壤微生物群落的影響時(shí),很少考慮土壤有機(jī)碳水平的影響。
桉樹人工林是我國南方廣泛種植的經(jīng)濟(jì)速生林,土壤養(yǎng)分缺乏(陳少雄,2009;廖觀榮,2003)。而氮肥的補(bǔ)充是維持桉樹養(yǎng)分的重要方式,則土壤中微生物群落及功能變化對(duì)營養(yǎng)周轉(zhuǎn)更為敏感。本研究以我國南方廣泛種植的桉樹人工林為研究對(duì)象,采用野外控制試驗(yàn)的方法,探究不同土壤有機(jī)碳水平下施氮對(duì)桉樹林土壤微生物群落結(jié)構(gòu)及功能的影響,其目標(biāo)是:明確不同有機(jī)碳水平桉樹林土壤微生物群落對(duì)施氮水平的響應(yīng)是否一致,并探討微生物群落變化在土壤養(yǎng)分維持中的意義。
1 研究區(qū)域概況與研究方法
1.1 研究區(qū)域概況
本研究樣地位于我國廣西扶綏縣國有東門林場(chǎng)(E107°15′~108°00′,N22°17′~22°30′)。廣西東門林場(chǎng)是亞洲最大的桉樹基因庫,主要經(jīng)營以桉樹為主的商品用材林。該地區(qū)屬于北熱帶季風(fēng)氣候區(qū),光熱充足,雨熱同季,夏濕冬干,年平均氣溫21.2~22.3 ℃,1月均溫13.2 ℃,7月均溫27.9 ℃。年降雨量1100~1300 mm,主要集中在6—8月,占全年降雨量的51.03%。研究區(qū)域土壤以砂頁巖發(fā)育而成的赤紅壤為主,pH值在4.5~6.0。
1.2 實(shí)驗(yàn)設(shè)計(jì)
為了研究不同有機(jī)碳水平桉樹林土壤微生物群落碳代謝功能對(duì)施氮的響應(yīng)特征,本研究從預(yù)實(shí)驗(yàn)的 20個(gè)樣地中,選取土壤有機(jī)碳含量差異顯著(P<0.01)的 2個(gè)二代桉樹林樣地:高有機(jī)碳樣地(HC):有機(jī)碳含量 14.80 g·kg-1;低土壤有機(jī)碳樣地(LC):有機(jī)碳含量12.89 g·kg-1),開展野外控制實(shí)驗(yàn)。兩塊樣地平均坡度約為10°。林木行距4 m,
株距2 m,林下植被有桃金娘Rhodomyrtus tomentos、余甘子Phyllanthus emblica、三叉苦Euodi lepta、飛機(jī)草Eupatorium odoratum、白茅Imperata cylindrica等,植被覆蓋度為60%。樣地的基本特征見表1。
結(jié)合當(dāng)?shù)厥┓柿?xí)慣和施肥強(qiáng)度,在桉樹林樣地中設(shè)置3個(gè)施氮梯度:對(duì)照(CK,0 kg·hm-2)、常規(guī)氮(NN,166.8 kg·hm-2)和高氮(HN,333.7 kg·hm-2)。每個(gè)施氮水平設(shè)置重復(fù) 3個(gè),每個(gè)樣方面積10 m×10 m。施氮時(shí)間為2013年5月20日,肥料種類為脲甲醛緩釋肥。取樣時(shí)間為 2013年 8月26日。
1.3 土壤理化性質(zhì)測(cè)定及酶活測(cè)定
土壤理化性質(zhì)測(cè)定參考土壤農(nóng)化分析(鮑士旦,2000)。土壤總碳、總氮使用元素分析儀測(cè)定(Vario EL III,Elementar,Germany),有機(jī)碳含量用重鉻酸鉀滴定法(LYT 1237-1999),土壤pH值采用土水比(質(zhì)量體積比)為1:2.5的方法測(cè)定。土壤可溶性有機(jī)碳由0.5 mol/L MK2SO4提取(H?gberg等2002;Jones和Willett,2006)并通過Liqui TOCⅡ分析儀(Elementar,Germany)測(cè)定,土壤酶活的測(cè)定參考DeForest熒光酶測(cè)定方法,及L-DOPA為底物酶培養(yǎng)測(cè)定方法(Deforest, 2009; Marx等,2001)。
1.4 磷脂脂肪酸(Phospholipid Fatty-acid Analysis,PLFA)提取、分離、鑒定
利用有機(jī)試劑對(duì)PLFA進(jìn)行提取(氯仿、甲醇),分離(氯仿、甲醇、丙酮),甲酯化(甲醇、甲苯、正己烷)后,上機(jī)前用1 mL含內(nèi)標(biāo)物19:0的正己烷溶液溶解吹干的脂肪酸甲脂,進(jìn)行 GC-MS(HP6890/MSD5793(Agilent Technologies,Bracknell,UK))測(cè)試(陳法霖等,2012;Frosteg?rd 等,1996)。
1.5 數(shù)據(jù)分析
據(jù)表2將PLFA劃分不同的微生物類群,并計(jì)算細(xì)菌和真菌含量,及真菌與細(xì)菌的比例。
通過主成分分析及方差分析實(shí)現(xiàn)不同處理上土壤微生物群落結(jié)構(gòu)的差異,由 CANOCO 5.0及SPSS 16.0統(tǒng)計(jì)軟件實(shí)現(xiàn),畫圖工具為Excel。

表1 土壤基本性質(zhì)Table 1 Property of the soil in the experiment

表2 微生物群落分類的PLFA標(biāo)記Table 2 PLFA markers for taxonomic microbial groups

圖1 土壤微生物PLFA總量Fig.1 Soil microbial total PLFA
2 結(jié)果
2.1 土壤微生物群落PLFA總量
施氮顯著影響土壤微生物群落磷脂脂肪酸總量(P<0.01),施二倍氮素處理(HN)的土壤微生物磷脂脂肪酸總量顯著低于對(duì)照(CK)和常規(guī)施氮處理(NN)(圖1)。高有機(jī)碳水平土壤微生物群落磷脂脂肪酸生物量顯著高于低有機(jī)碳水平(P<0.01)。
土壤有機(jī)碳水平和施氮水平對(duì)土壤微生物群落磷脂脂肪酸總量的影響存在交互作用(P<0.01),高有機(jī)碳水平與對(duì)照、常規(guī)施氮水平和施二倍氮素組合的土壤微生物群落磷脂脂肪酸總量均顯著高于低有機(jī)碳水平與對(duì)照、常規(guī)施氮水平和施二倍氮素組合,低有機(jī)碳水平和施二倍氮素組合的土壤微生物群落磷脂脂肪酸總量顯著最低。
2.2 微生物群落結(jié)構(gòu)
2.2.1 微生物群落組成
施氮顯著影響土壤細(xì)菌、真菌、放線菌磷脂脂肪酸量以及真菌/細(xì)菌比值,施二倍氮素處理的土壤細(xì)菌、放線菌磷脂脂肪酸量顯著低于對(duì)照和常規(guī)施氮處理;隨著施氮水平的增加,土壤真菌磷脂脂肪酸量和真菌/細(xì)菌比值顯著降低,對(duì)照、常規(guī)施氮水平和施二倍氮素處理之間土壤真菌磷脂脂肪酸量、真菌/細(xì)菌比值差異均達(dá)顯著水平(P<0.05)。高有機(jī)碳土壤細(xì)菌、真菌、放線菌磷脂脂肪酸生物量以及真菌/細(xì)菌比值均顯著高于低有機(jī)碳土壤處理(表3)。
土壤有機(jī)碳水平和施氮水平對(duì)土壤細(xì)菌、真菌、放線菌和真菌/細(xì)菌比值的影響存在交互作用,在低土壤有機(jī)碳水平中,施二倍氮素能顯著降低細(xì)菌磷脂脂肪酸量,但在高土壤有機(jī)碳水平中施氮對(duì)細(xì)菌磷脂脂肪酸量的影響不顯著。在低、高土壤有機(jī)碳水平中,真菌磷脂脂肪酸量對(duì)施氮的響應(yīng)不一致,在低土壤有機(jī)碳水平中,常規(guī)施氮水平顯著降低了真菌磷脂脂肪酸量;而在高土壤有機(jī)碳水平中,施二倍氮素水平才顯著降低真菌磷脂脂肪酸量。在低、高土壤有機(jī)碳土壤中,常規(guī)施氮水平均顯著降低了土壤真菌/細(xì)菌比值。
2.2.2 微生物群落主成分分析
土壤微生物群落磷脂脂肪酸PLFA主成分分析表明:PC1可以解釋56.63%的變異,土壤微生物群落的差異主要表現(xiàn)在主成分1(PC1),即:不同有機(jī)碳水平下的土壤微生物群落;在2種有機(jī)碳水平上,高氮與其他處理之間分異均明顯(圖2)。
真菌特征脂肪酸 16:1ω5c、18:1ω9c、18:2ω9c及細(xì)菌特征脂肪酸16:1ω7c、i17:0和放線菌特征脂肪酸10Me18:0等與PC1顯著相關(guān),而16:0、14:0、17:0、18:0在PC2上得分較高,這些脂肪酸對(duì)土壤微生物群落分異起到主要作用。
2.3 土壤酶活性及可溶性有機(jī)碳(DOC)特征
施氮顯著增加了土壤纖維素酶、酚氧化酶活性及土壤可溶性有機(jī)碳含量,施二倍氮素處理的土壤纖維素酶活性顯著高于對(duì)照和常規(guī)施氮水平處理;對(duì)照、常規(guī)施氮水平和施二倍氮素處理的酚氧化酶差異和土壤有機(jī)碳含量差異均顯著(P<0.05)(表4)。土壤有機(jī)碳水平也影響土壤纖維素酶活性和土壤可溶性有機(jī)碳含量,高土壤有機(jī)碳樣地中的土壤纖維素酶活性和土壤可溶性有機(jī)碳含量顯著高于低土壤有機(jī)碳樣地(表4)。
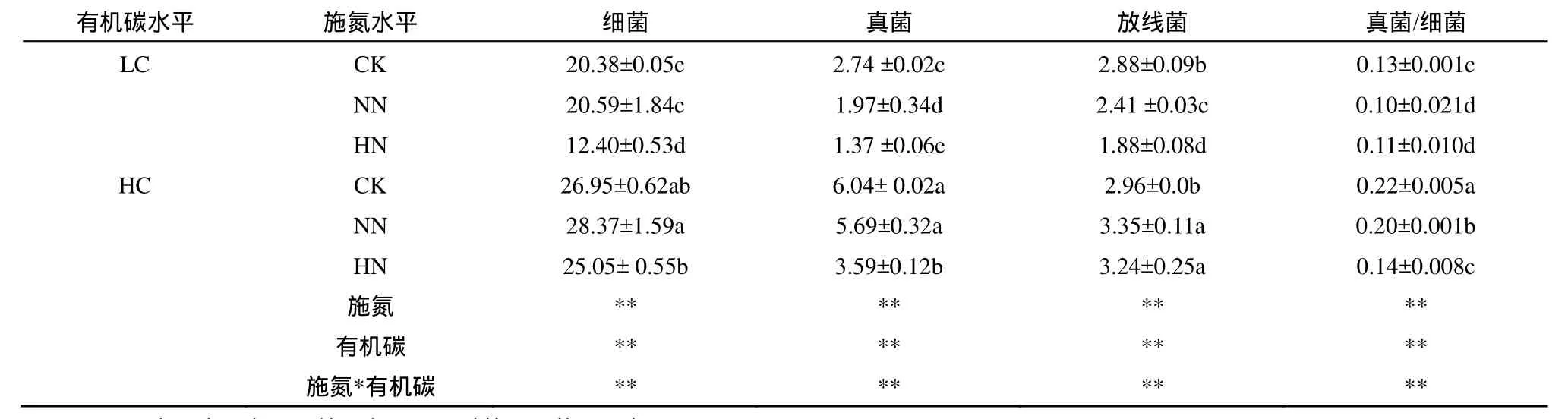
表3 土壤微生物群落組成1)Table 3 Soil microbial community composition
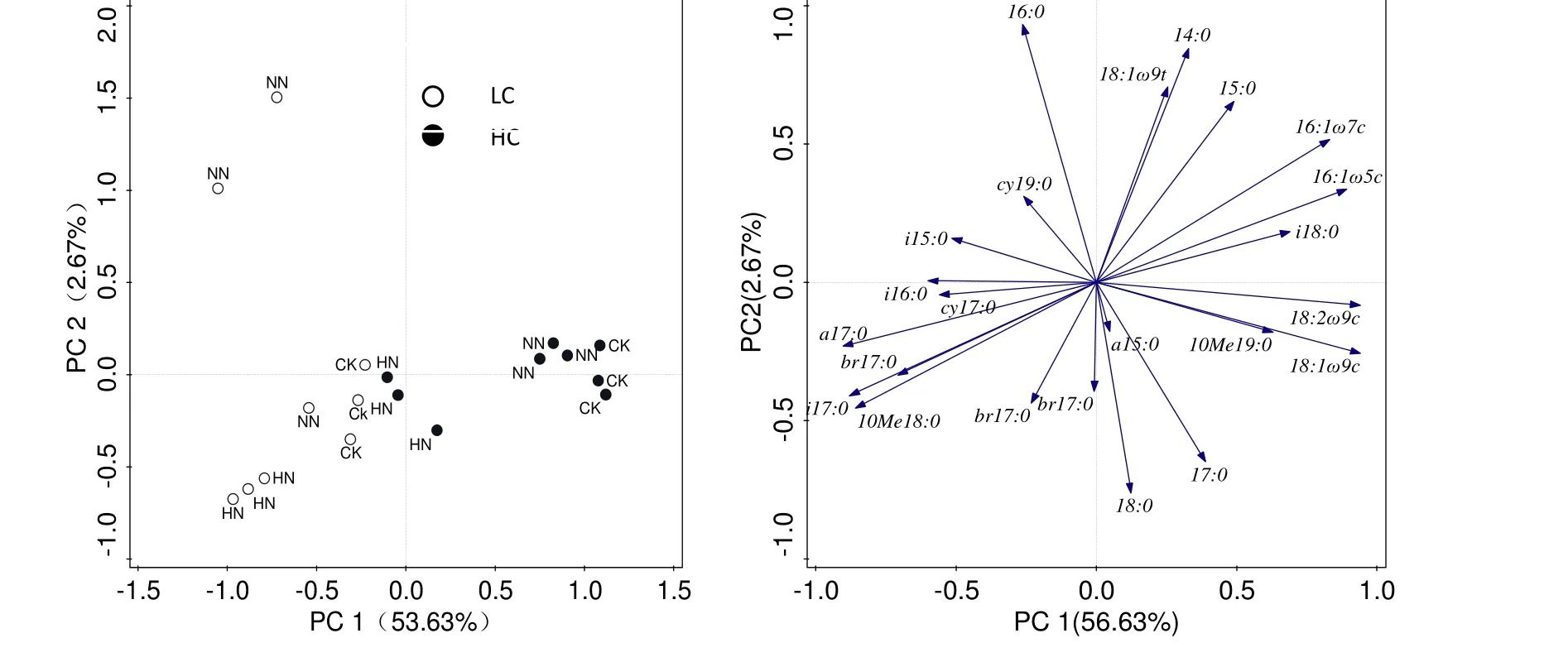
圖2 土壤微生物的群落結(jié)構(gòu)主成分分析Fig.2 The principal components analysis of soil microbial community composition
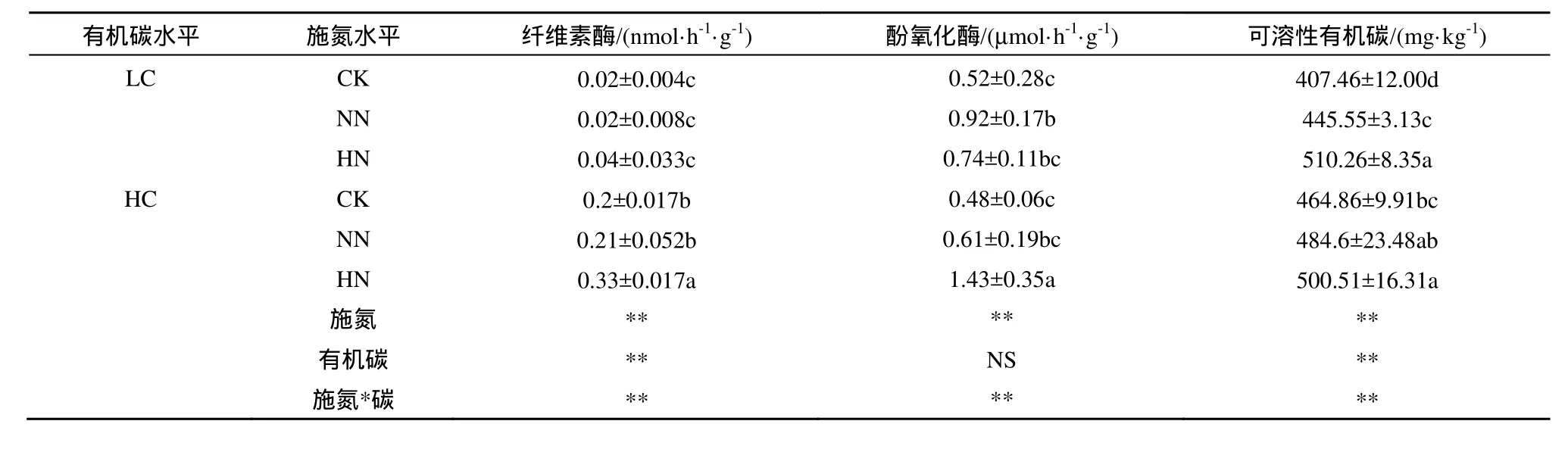
表4 土壤酶活及DOC 含量Table 4 Soil extracellular enzymes activity and DOC content
施氮水平和土壤有機(jī)碳含量對(duì)土壤纖維素酶、酚氧化酶活性及土壤可溶性有機(jī)碳含量的影響存在交互作用。在低土壤有機(jī)碳樣地中,土壤纖維素酶活性對(duì)施氮的響應(yīng)沒差異,但在高土壤有機(jī)碳樣地中,施二倍氮素處理的土壤纖維素酶活性顯著高于對(duì)照和常規(guī)施氮水平處理;在低土壤有機(jī)碳樣地中,常規(guī)施氮水平和施二倍氮素處理的酚氧化酶活性均顯著高于對(duì)照,而在高土壤有機(jī)碳樣地中,施二倍氮素處理的酚氧化酶顯著高于對(duì)照和常規(guī)施氮水平;對(duì)于土壤可溶性有機(jī)碳,低土壤有機(jī)碳樣地中,隨施氮水平的增加,土壤可溶性有機(jī)碳含量顯著增加,而在高土壤有機(jī)碳樣地中,施二倍氮素處理的土壤可溶性有機(jī)碳含量才顯著高于對(duì)照(表4)。
3 討論
土壤中氮可以通過調(diào)節(jié)微生物參與的碳氮代謝過程而影響微生物群落結(jié)構(gòu)(DeForest等,2004)。本實(shí)驗(yàn)發(fā)現(xiàn)施氮顯著影響了土壤微生物群落:(1)降低土壤微生物群落磷脂脂肪酸總量及土壤中細(xì)菌、真菌、放線菌磷脂脂肪酸量(P<0.05)。可能是因?yàn)橥庠吹奶砑佑绊懥送寥乐形⑸锔呈澄镦湥档臀⑸飳?duì)凋落物的吸收(Gan等2013),進(jìn)而降低了土壤微生物的PLFA總量。氮添加處理后革蘭氏陽性及陰性菌、真菌豐度都降低,這與Bi等的實(shí)驗(yàn)結(jié)果一致(Bi等,2012)。(2)施氮處理下,真菌/細(xì)菌比值顯著降低(P<0.05)。可能是施氮改變了土壤有機(jī)物的組成,而減少了真菌可利用有機(jī)物的比例,從而抑制了真菌的生長(zhǎng)。Norris等認(rèn)為土壤中外源氮的添加超過一定有效氮閾值,微生物真菌豐度降低,有真菌主導(dǎo)的微生物群落轉(zhuǎn)化為細(xì)菌主導(dǎo)(Janssens等,2010),進(jìn)而改變土壤微生物分解功能(Norris等,2012;Wallenstein等,2006),同樣Tietema發(fā)現(xiàn)氮飽和土壤中真菌含量降低(Tietema等,1998),與本文實(shí)驗(yàn)結(jié)果表現(xiàn)一致。
同時(shí)施氮土壤影響微生物群落進(jìn)而調(diào)節(jié)了土壤中酶活,而影響土壤碳代謝過程(Sinsabaugh等,2005;Carreiro等2000)。本實(shí)驗(yàn)發(fā)現(xiàn)施氮顯著增加了土壤纖維素酶、酚氧化酶活性及土壤中 DOC含量。酶活性在不同氮水平間的變化與相應(yīng)微生物PLFA總量變化并不一致,可能是由于施氮主要是通過影響微生物功能群的變化而調(diào)節(jié)土壤酶活(Gallo等,2004)。而與此前報(bào)道(Frey等,2004)有差異的是,酚氧化酶活性并沒有受到氮的抑制,而是對(duì)氮做出正響應(yīng)(P<0.05),說明施氮并沒有影響酚氧化酶的主要微生物功能群,其機(jī)制需要進(jìn)一步通過分子生物學(xué)等方法探究。土壤可溶性有機(jī)碳(dissolved organic carbon, DOC)來源于微生物對(duì)凋落物等植物殘?bào)w及土壤有機(jī)物的分解,與微生物參與的代謝過程顯著相關(guān)(方華軍等,2007)。原因可能是兩個(gè)方面:從來源上來說,較高的纖維素酶活性能夠分解有機(jī)物成為DOC;而從損失上來說,DOC是土壤微生物的能量來源(Yano等,2000),HN處理下微生物總量降低,對(duì)DOC的利用降低,這兩方面最終結(jié)果表現(xiàn)為DOC總量的增加。
同時(shí),本實(shí)驗(yàn)發(fā)現(xiàn)土壤有機(jī)碳影響土壤微生物群落結(jié)構(gòu)及功能。在不同有機(jī)碳水平土壤中,微生物總量變化顯著(P<0.01),高有機(jī)碳水平桉樹林土壤微生物PLFA高于低有機(jī)碳水平桉樹林,強(qiáng)調(diào)了土壤有機(jī)碳為微生物提供能量,而調(diào)節(jié)微生物的生長(zhǎng)(Fontaine等,2004;劉來等,2013),而Williams發(fā)現(xiàn)土壤有機(jī)碳顯著影響微生物群落組成(Williams等,2013),與本實(shí)驗(yàn)結(jié)果一致。不同有機(jī)碳水平影響了土壤微生物群落,從而影響土壤中酶活性及 DOC的變化,整體表現(xiàn)為高土壤有機(jī)碳水平上2種酶活性及DOC高于低土壤有機(jī)碳水平。
此外,本研究發(fā)現(xiàn)土壤微生物對(duì)施氮的響應(yīng)受到有機(jī)碳的調(diào)節(jié)。低有機(jī)碳水平土壤中,微生物群落受到有機(jī)碳的限制,而對(duì)氮的添加更為敏感,在低氮(常規(guī)氮)水平上表現(xiàn)出群落結(jié)構(gòu)及功能降低的趨勢(shì)。而在高土壤有機(jī)碳水平樣地中只有在高氮(施二倍氮素)處理中才顯著降低或不變化。在不同有機(jī)碳水平上,施氮可能改變了土壤的碳氮比,而表現(xiàn)為不同有機(jī)碳水平上微生物群落結(jié)構(gòu)及功能的差異(Allison等,2008;Liu和 Greaver, 2010)。施氮傾向于引起土壤中碳氮比降低,當(dāng)?shù)枯^豐富時(shí),微生物可利用的有效氮增加,但是土壤中可利用的碳成為微生物的限制因素(Aber等,1989),所以產(chǎn)生施氮條件下,土壤微生物整體豐度降低。而在高有機(jī)碳土壤中,有效碳的含量高,而土壤微生物對(duì)氮的響應(yīng),所表現(xiàn)出的碳的限制并不明顯。
本實(shí)驗(yàn)結(jié)果表明施氮顯著影響了土壤中微生物群落結(jié)構(gòu)及功能,同時(shí),土壤微生物的響應(yīng)又受到土壤有機(jī)碳的調(diào)節(jié),而且不同有機(jī)碳水平上微生物群落及功能變化對(duì)氮的響應(yīng)并不一致。不同施氮水平對(duì)人工林生態(tài)系統(tǒng)中土壤微生物的影響,應(yīng)當(dāng)考慮土壤中有機(jī)碳水平。
4 結(jié)論
通過對(duì)不同有機(jī)碳水平桉樹林土壤中微生物實(shí)驗(yàn)的研究,我們得出以下結(jié)論。
(1)施氮顯著影響土壤微生物群落結(jié)構(gòu)及功能,土壤微生物群落PLFA總量隨著施氮的增加反而降低。而且隨著施氮的增加,土壤微生物中真菌與細(xì)菌比例降低;而土壤微生物群落功能——土壤微生物酶活性,在施氮處理的影響下而增加,說明施氮促進(jìn)部分微生物分解功能增強(qiáng)。
(2)土壤微生物群落結(jié)構(gòu)以及功能的變化同時(shí)受到土壤中有機(jī)碳水平的調(diào)節(jié),在高有機(jī)碳水平土壤中,微生物 PLFA量高于低有機(jī)碳水平,同時(shí)土壤微生物酶活性也表現(xiàn)出不同有機(jī)碳水平上的顯著差異,在高有機(jī)碳水平上酶活要高于低有機(jī)碳水平。
(3)施氮和土壤有機(jī)碳變化對(duì)土壤微生物的影響有交互作用,不同有機(jī)碳水平上,土壤微生物群落組成差異顯著,尤其是真菌含量,在低土壤有機(jī)碳水平中,常規(guī)施氮水平下真菌磷脂脂肪酸量顯著降低;而在高土壤有機(jī)碳水平中,施二倍氮素水平才顯著降低真菌磷脂脂肪酸量。而碳氮對(duì)不同酶活性的影響表現(xiàn)不同的交互作用,酚氧化酶,纖維素酶在高有機(jī)碳高氮條件下活性高于其他處理,施氮在高有機(jī)碳的水平條件下促進(jìn)土壤微生物的分解功能。所以不同施氮水平對(duì)人工林生態(tài)系統(tǒng)中土壤微生物的影響研究,土壤中有機(jī)碳水平不容忽視。
ABER J D, NADELHOFFER K J, STEUDLER P, et al. 1989.Nitrogen saturation in northern forest ecosystems[J]. BioScience, 39(6):378-286.
ALLISON S D, CZIMCZIK C I,TRESEDER K K. 2008.Microbial activity and soil respiration under nitrogen addition in Alaskan boreal forest[J].Global Change Biology, 14(5): 1156-1168.
BI J, ZHANG N, LIANG Y, et al.2012. Interactive effects of water and nitrogen addition on soil microbial communities in a semiarid steppe[J].Journal of Plant Ecology,5(3): 320-329.
BURGER M,JACKSON L E.2003. Microbial immobilization of ammonium and nitrate in relation to ammonification and nitrification rates in organic and conventional cropping systems[J].Soil Biology and Biochemistry, 35(1): 29-36.
BUYER J S,SASSER M. 2012.High throughput phospholipid fatty acid analysis of soils[J].Applied Soil Ecology, 61: 127-130.
CARREIRO M, SINSABAUGH R, REPERT D, et al. 2000. Microbial enzyme shifts explain litter decay responses to simulated nitrogen deposition[J].Ecology, 81(9): 2359-2365.
CUSACK D F, SILVER W L, TORN M S, et al. 2011.Changes in microbial community characteristics and soil organic matter with nitrogen additions in two tropical forests[J].Ecology, 92(3): 621-632.
DEFOREST J L, ZAK D R, PREGITZER K S, et al. 2004.Atmospheric nitrate deposition and the microbial degradation of cellobiose and vanillin in a northern hardwood forest[J].Soil Biology and Biochemistry, 36(6): 965-971.
DEFOREST J L. 2009.The influence of time, storage temperature, and substrate age on potential soil enzyme activity in acidic forest soils using MUB-linked substrates and L-DOPA[J].Soil Biology and Biochemistry, 41(6): 1180-1186.
Fontaine S, Bardoux G, Abbadie L, et al. 2004.Carbon input to soil may decrease soil carbon content[J].Ecology letters, 2004, 7(4): 314-320.
FREY S D, KNORR M, PARRENT J L, et al. Chronic nitrogen enrichment affects the structure and function of the soil microbial community in temperate hardwood and pine forests[J].Forest Ecology and Management, 196(1): 159-171.
FROSTEG?RD ?,B??TH E.1996. The use of phospholipid fatty acid analysis to estimate bacterial and fungal biomass in soil[J].Biol Fert Soils, 22(1-2): 59-65.
GALLO M, AMONETTE R, LAUBER C, et al. 2004.Microbial community structure and oxidative enzyme activity in nitrogen-amended north temperate forest soils[J].Microbial Ecology, 48(2): 218-229.
GAN H, ZAK D R,HUNTER M D. 2013.Chronic nitrogen deposition alters the structure and function of detrital food webs in a northern hardwood ecosystem[J].Ecological Applications,23(6):1311-1321.
HOBBIE S E. Temperature and plant species control over litter decomposition in Alaskan tundra[J].Ecological Monographs, 1996,66(4): 503-522.
H?GBERG M N,H?GBERG P. 2002.Extramatrical ectomycorrhizal mycelium contributes one‐third of microbial biomass and produces,together with associated roots, half the dissolved organic carbon in a forest soil[J].New Phytologist, 154(3): 791-795.
JANSSENS I, DIELEMAN W, LUYSSAERT S, et al. 2010.Reduction of forest soil respiration in response to nitrogen deposition[J].Nature Geoscience, 3(5): 315-322.
JONES D, WILLETT V. 2006. Experimental evaluation of methods to quantify dissolved organic nitrogen (DON) and dissolved organic carbon (DOC) in soil[J].Soil Biology and Biochemistry, 38(5):991-999.
LIU L, GREAVER T L. 2010. A global perspective on belowground carbon dynamics under nitrogen enrichment[J]. Ecol Lett, 13(7): 819-28.
MARX M-C, WOOD M, JARVIS S. 2001. A microplate fluorimetric assay for the study of enzyme diversity in soils[J].Soil Biology and Biochemistry, 33(12): 1633-1640.
NORRIS M D, AVIS P G, REICH P B, et al. 2013. Positive feedbacks between decomposition and soil nitrogen availability along fertility gradients[J]. Plant and Soil, 367(1/2): 347-361.
OLSSON P A, B??TH E, JAKOBSEN I, et al. 1995.The use of phospholipid and neutral lipid fatty acids to estimate biomass of arbuscular mycorrhizal fungi in soil[J].Mycological Research, 99(5):623-629.
OSONO T. 2007.Ecology of ligninolytic fungi associated with leaf litter decomposition[J].Ecological Research, 22(6): 955-974.
SINSABAUGH R L, GALLO M E, LAUBER C, et al. 2005.Extracellular Enzyme Activities and Soil Organic Matter Dynamics for Northern Hardwood Forests receiving Simulated Nitrogen Deposition[J].Biogeochemistry, 75(2): 201-215.
TIETEMA A. 1998.Microbial carbon and nitrogen dynamics in coniferous forest floor material collected along a European nitrogen deposition gradient[J].Forest Ecology and Management, 101(1): 29-36.
WALDROP M P, ZAK D R,SINSABAUGH R L.2004. Microbial community response to nitrogen deposition in northern forest ecosystems[J].Soil Biology and Biochemistry, 36(9): 1443-1451.
WALLENSTEIN M D, McNULTY S, FERNANDEZ I J, et al.2006.Nitrogen fertilization decreases forest soil fungal and bacterial biomass in three long-term experiments[J].Forest Ecology and Management, 222(1): 459-468.
WILLIAMS A, BORJESSON G, HEDLUND K. 2013.The effects of 55 years of different inorganic fertiliser regimes on soil properties and microbial community composition[J]. Soil Biology & Biochemistry, 67:41-46.
YANO Y, McDOWELL W, ABER J. 2000.Biodegradable dissolved organic carbon in forest soil solution and effects of chronic nitrogen deposition[J].Soil Biology and Biochemistry, 32(11): 1743-1751.
ZHONG W, GU T, WANG W, et al. 2010. The effects of mineral fertilizer and organic manure on soil microbial community and diversity[J].Plant and Soil, 326(1-2): 511-522.
鮑士旦, 2000.土壤農(nóng)化分析[M]. 北京:中國農(nóng)業(yè)出版社.
陳法霖, 鄭華, 歐陽志云, 等. 2011. 土壤微生物群落結(jié)構(gòu)對(duì)凋落物組成變化的響應(yīng)[J]. 土壤學(xué)報(bào), 48(3): 603-611.
陳少雄. 2009. 桉樹人工林土壤養(yǎng)分現(xiàn)狀與施肥研究[J].桉樹科技, 26(1):52-63.
方華軍, 程淑蘭,于貴瑞. 2007. 森林土壤碳, 氮淋失過程及其形成機(jī)制研究進(jìn)展[J]. 地理科學(xué)進(jìn)展, 26(3): 29-37.
廖觀榮, 李淑儀, 藍(lán)佩玲, 等. 2003. 桉樹人工林生態(tài)系統(tǒng)養(yǎng)分循環(huán)與平衡研究 Ⅰ. 桉樹人工林生態(tài)系統(tǒng)的養(yǎng)分貯存[J].生態(tài)環(huán)境, 12(2):150-154.
劉來, 黃保健, 孫錦,等. 2013. 大棚辣椒連作土壤微生物數(shù)量、酶活性與土壤肥力的關(guān)系[J]. 中國土壤與肥料, (2):5-10.
吳愉萍. 2009.基于磷脂脂肪酸 (PLFA) 分析技術(shù)的土壤微生物群落結(jié)構(gòu)多樣性的研究[D]. 杭州: 浙江大學(xué).
張秋芳, 劉波, 林營志, 等. 2009.土壤微生物群落磷脂脂肪酸PLFA生物標(biāo)記多樣性[J].生態(tài)學(xué)報(bào), 29(008): 4127-4137.
猜你喜歡
美與時(shí)代·美術(shù)學(xué)刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(shè)(2019年6期)2019-10-08 08:55:48
人大建設(shè)(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(shè)(2017年6期)2017-09-26 11:50:44
學(xué)苑創(chuàng)造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問題研究(2012年1期)2012-03-25 09:54:45
中國火炬(2010年12期)2010-07-25 13:26:22
中國火炬(2010年8期)2010-07-25 11:34:30
