不同水生植物抑藻效果及機理研究
2014-05-25 00:33:06蘇躍龍王正芳
環(huán)境科學(xué)導(dǎo)刊 2014年6期
蘇躍龍,王正芳
(1.云龍縣環(huán)境監(jiān)測站,云南云龍672700;2.復(fù)旦大學(xué)環(huán)境科學(xué)與工程系,上海200433)
不同水生植物抑藻效果及機理研究
蘇躍龍1,王正芳2
(1.云龍縣環(huán)境監(jiān)測站,云南云龍672700;2.復(fù)旦大學(xué)環(huán)境科學(xué)與工程系,上海200433)
對水芹、美人蕉、黃菖蒲、菹草4種水生植物對銅綠微囊藻的化感特征進(jìn)行了研究。不同的植物組織器官及不同的植物處理方式下,抑藻效果不同,最具抑藻潛力的植物為黃菖蒲和美人蕉。經(jīng)黃菖蒲和美人蕉種植水培養(yǎng)5d后,銅綠微囊藻的抑制率達(dá)70.12%~77.25%。質(zhì)量分?jǐn)?shù)為0.53%的黃菖蒲葉提取物,經(jīng)11d的培養(yǎng)后,對銅綠微囊藻的抑制率為65.30%。抑藻機理可能在于化感物質(zhì)的脅迫使微囊藻細(xì)胞內(nèi)活性氧自由基過量堆積,進(jìn)而引起對藻細(xì)胞的傷害和影響藻類正常的光合作用。SOD酶活性與藻的比增長率均呈現(xiàn)較好的負(fù)相關(guān)關(guān)系。
水生植物;銅綠微囊藻;超氧化物歧化酶;抑藻;化感作用
植物化感抑藻為富營養(yǎng)化湖泊水體藍(lán)藻的治理提供了新的途徑。大部分化感物質(zhì)是植物生長中產(chǎn)生的次生代謝物質(zhì),在環(huán)境中不易造成危害與污染,較易于降解,具有很好的生態(tài)安全性[1]。越來越多的研究表明[2,3],植物化感抑藻是可以替代物理、化學(xué)治理的安全有效的方法。目前,陸生植物化感作用的研究進(jìn)行得較為深入,大麥秸稈[4]、灌木[5]、稻殼[6]、柑橘皮、香蕉皮[7]等都表現(xiàn)出較好的抑藻效果。
水生植物的化感作用雖與陸生植物類似,但其在化感特征、作用方式等方面存在一些差別:水生植物的代謝產(chǎn)物更為復(fù)雜,化感物質(zhì)在水體環(huán)境中更易稀釋、擴散,抑藻效果受水體中化感物質(zhì)濃度影響較大[8]。胡光濟(jì)等人[9]研究了水生植物菖蒲的干體提取液對銅綠微囊藻生長的影響,結(jié)果表明,菖蒲干體提取液中含有抑藻活性的化感物質(zhì),高濃度的提取液(>40ml/L)對銅綠微囊藻的抑制率高達(dá)98.66%;而低濃度的提取液(<30ml/L)對藻的生長并無顯著影響。Wu等人[10]從浮萍中成功提取出化感物質(zhì),當(dāng)化感物質(zhì)濃度為60mg/L時,抑藻率為50%,藻細(xì)胞內(nèi)的葉綠素a含量減少50%;當(dāng)化感物濃度提高到100mg/L時,抑藻率可達(dá)90%,藻細(xì)胞內(nèi)的葉綠素a含量可減少75%。
水生植物釋放的化感物質(zhì)降解速率較快,在非連續(xù)分泌情況下只能對浮游藻類產(chǎn)生短期的抑制作用[11,12]。因此,在天然淺水湖泊中,化感物質(zhì)連續(xù)釋入環(huán)境,在水體中維持一定濃度才可發(fā)揮有效的抑藻作用。不同植物產(chǎn)生的化感抑藻化合物的種類和數(shù)量都可能不同,即便是同一植物在不同的培養(yǎng)體系中產(chǎn)生的相同化感物質(zhì)其抑藻能力也可能存在很大差異,在同一植物體內(nèi)不同組織、器官化感化合物的量也會不同,植物產(chǎn)生和分泌化感物質(zhì)的差異,可能對其抑藻能力產(chǎn)生較大影響。
化感物質(zhì)抑藻途徑可能有以下幾種:① 破壞葉綠素。有些化感物質(zhì)通過破壞藻類的葉綠素,影響其光合作用,從而抑制藻類的生長。②破壞細(xì)胞膜。化感物質(zhì)能降低細(xì)胞膜的完整性,使細(xì)胞內(nèi)物質(zhì)大量滲出,滲出液的電導(dǎo)率增加。③影響酶活性。化感物質(zhì)能影響生物體的酶活性,由于酶的特性不同,化感物質(zhì)能提高某些酶的活性,卻能抑制另一些酶的活性。
本文以富營養(yǎng)化水體為對象,選取水芹、美人蕉、黃菖蒲、菹草4種水生植物,通過人工模擬自然條件的方法,一方面,通過研究各植物對氮磷離子吸收濃度動力學(xué)差異,研究植物根、莖、葉各器官中氮磷的分布特征,分別從基質(zhì)相、植物相、水相中氮磷營養(yǎng)物質(zhì)含量及濃度的變化來探討不同水生植物系統(tǒng)中氮磷在水相、基質(zhì)相、植物相中的富集與轉(zhuǎn)移效果,確定各相作用對氮磷去除的貢獻(xiàn)率,并判別氮磷去除率與植物根系基質(zhì)酶的相互關(guān)系。另一方面,開展不同水生植物、不同植物組織器官及化感物質(zhì)的不同提取方式對銅綠微囊藻的化感作用研究,考察其抑藻特性,確定其生長拐點時間及受損機制。結(jié)合氮磷削減及抑藻潛力對各水生植物進(jìn)行綜合評價,以期為利用水生植物抑制藻類生長、削減富營養(yǎng)化水體中氮磷、創(chuàng)造穩(wěn)定優(yōu)美的水生生態(tài)環(huán)境提供一定的理論依據(jù)。
1 材料與方法
1.1 植物材料
選擇4種水生植物:水芹(Oenanthe javanica)、黃菖蒲(Iris pseudacorus L)、美人蕉(Canna lily)、菹草(Potamogeton crispus)。在試驗前將植物用去離子水沖洗干凈,轉(zhuǎn)入0.2mmol/L的CaSO4溶液中,饑餓培養(yǎng)24h后待用。
1.2 化學(xué)試劑
試驗所用CaSO4、Ca(ClO)2等化學(xué)試劑均為市售分析純試劑。
1.3 實驗方法
1.3.1 藻的培養(yǎng)
將銅綠微囊藻(Microcystis aeruginosa,F(xiàn)ACHB-905)移入滅菌后的2L培養(yǎng)基中擴培,培養(yǎng)條件為:25℃,光照:12h,光強度2000 lux,記錄其生長曲線,取處于對數(shù)末期的藻液開展以下抑藻試驗。培養(yǎng)期間每日搖勻2次。
1.3.2 植物提取物抑藻試驗
將葉片用自來水水洗,去除附著的泥沙等雜質(zhì)后,用蒸餾水沖洗2~3遍。之后置于濾紙上,室溫干燥10min。新鮮葉片剪成<5mm×5mm大小,用四分法取出10g。混勻后,10g葉片混合物置于100m l蒸餾水中,水浴加熱2h。冷卻后,經(jīng)玻璃纖維濾紙(Whatman GF/C)過濾,將濾液定容至100ml,此時提取物濃度為10g/100ml=10%[4]。根莖提取物制備方法與葉片組織相同。將8ml植物提取物添加到BG-11培養(yǎng)基中,定容為150ml,使植物提取物體積分?jǐn)?shù)(V∶V)=5.3%,此時錐形瓶中有相當(dāng)于0.8 g植物組織的提取物,即植物提取物質(zhì)量分?jǐn)?shù) (m∶V) =0.53%。將錐形瓶置于高壓滅菌鍋中滅菌30min,待降至常溫后,將已培養(yǎng)好的藻液離心 (6000 rpm,10℃,4 min),接種,藻密度控制為3.0× 106cells/ml。另外,設(shè)不加植物提取物的藻密度為3.0×106cells/ml的錐形瓶作為空白。培養(yǎng)條件:25℃,光照:12h,光強度2000lux。培養(yǎng)期間每日搖勻2次。
1.3.3 植物組織抑藻試驗
將葉片用自來水水洗,去除附著的泥沙等雜質(zhì)后,用蒸餾水沖洗2~3遍。之后置于濾紙上,室溫干燥10min。稱取5 g新鮮葉片剪成5mm×5mm,在10%Ca(ClO)2溶液中泡15min,蒸餾水水洗4次。水洗后瀝干水分,在濾紙上室溫干燥10min。根莖組織制備方法與葉片組織相同。將0.8g植物組織添加到BG-11培養(yǎng)基中,定容為150ml,使植物組織質(zhì)量分?jǐn)?shù)(m∶V)=0.53%[13]。后續(xù)步驟同上。
1.3.4 種植水抑藻試驗
水生植物種植35d后,取其種植水先用中速濾紙抽濾一遍,后經(jīng)0.45μm微孔濾膜減壓過濾,濾掉可能帶入的原生動物及微生物等。過濾裝置為真空抽濾器及砂芯過濾裝置。取100ml種植水,加入已滅菌并裝有50 m培養(yǎng)基的三角瓶中,再接種藻類,使起始藻密度為3.0×106cells/ml,對照用同樣經(jīng)過真空過濾的自來水100ml,代替種植水。在培養(yǎng)過程中,用封口膜封住錐形瓶瓶口,每天定時搖動2次。
1.3.5 藻細(xì)胞超氧化物歧化酶的測定
按照GB/T5009.171-2003中鄰苯三酚自氧化的方法測定酶活力。
1.3.6 藻生物量的測定
以藻密度表示(cell/m l),取少量混合均勻的藻液,加入一滴盧哥氏液固定后,用滴管在藻液中部吸取少量藻液,緩慢滴入血細(xì)胞計數(shù)板 (事先將蓋玻片蓋好),用吸水紙吸去多余液體,在顯微鏡下用血球計數(shù)板對細(xì)胞數(shù)量計數(shù)2次,若2次相差超過20%則進(jìn)行第3次。
1.4 計算方法
1.4.1 藻抑制率及生長參數(shù)的確定
抑制率公式:IR=(1-Nt/Mt)×100%
式中:IR—抑制率,Nt—處理組第t天藻細(xì)胞密度值,Mt—對照組第t天藻細(xì)胞密度值。
以Logistic方程擬合藻類的增長過程。以每個處理組的最大生物量 (Nm)作為各自K的估計值。應(yīng)用其對數(shù)形式Ln[(K-Nt)/Nt]=a-rt,以最小二乘法進(jìn)行回歸分析,獲得該方程的斜率和截距作為a和r的估計值。藻類的生長拐點即為Logistic方程的二階導(dǎo)數(shù)等于0的時間,由公式tp=(a-ln2)/r來計算。
1.4.2 銅綠微囊藻比增長速率μ的計算
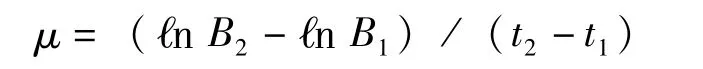
式中:μ-比增長速率 (d-1),B2-某一時間間隔終結(jié)時藻類的生物量(cell/ml),B1-某一時間藻類的生物量(cell/ml),t2-t1-時間間隔(d)。
1.4.3 最大生物量Nmax的確定
從接種當(dāng)天起,隔天取樣計數(shù)銅綠微囊藻的個數(shù),當(dāng)藻類增長<5%時,取其藻量作為最大生物量(×106cell/ml),用每毫升藻液中所含的藻細(xì)胞個數(shù)表示。
2 結(jié)果與討論
2.1 不同植物根莖葉的抑藻效果
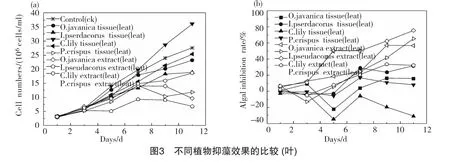
圖1為不同植物根的抑藻效果,與對照相比,水芹、黃菖蒲、美人蕉的根組織器官及其提取物組中藻的生長均受到不同程度的抑制。總體上看來,前5 d抑藻效果不明顯,第5d,對照組藻密度為1.03×107cell/ml,其他各組的藻密度為9.61×106~1.10×107cell/ml,抑藻率在-6.77%~6.77%波動;5d后各組藻類生長趨勢開始有了明顯不同。組織器官組中,黃菖蒲根抑藻效果最好,第11d生物量降為1.33×107cell/ml,與對照組生物量2.76× 107cell/ml相比,其抑制率為51.81%;水芹組及美人蕉組抑藻效果較差,試驗結(jié)束時抑藻率僅為4.00%、9.73%。與組織器官組相比,水芹及美人蕉根提取物組表現(xiàn)為更好的抑藻效果,第11d時,抑制率分別為33.10%、20.68%;黃菖蒲根提取物組表現(xiàn)較差,試驗結(jié)束時,其抑藻率僅為16.43%,低于黃菖蒲根組織組。
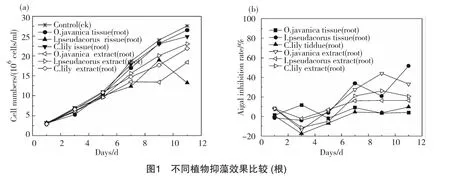
從圖2可以看出,莖組織器官組中抑藻效果最為明顯的是美人蕉莖組,其生物量從第5d后,緩慢增加,不再呈對數(shù)式增長,試驗結(jié)束時,其生物量為1.52×107cell/ml,與對照組2.76×107cell/ml相比抑制率為44.95%;菹草莖組抑藻效果次之,11d時,其抑制率為13.48%;黃菖蒲莖組表現(xiàn)最差。組織提取物組中,美人蕉及菹草組在0~11d均表現(xiàn)出抑藻效果,抑制率變化較為平穩(wěn),但值較低,在2.66%~24.47%內(nèi)波動;水芹及黃菖蒲組在7d后才開始有抑藻效果,抑藻率保持在18.80%~29.33%。
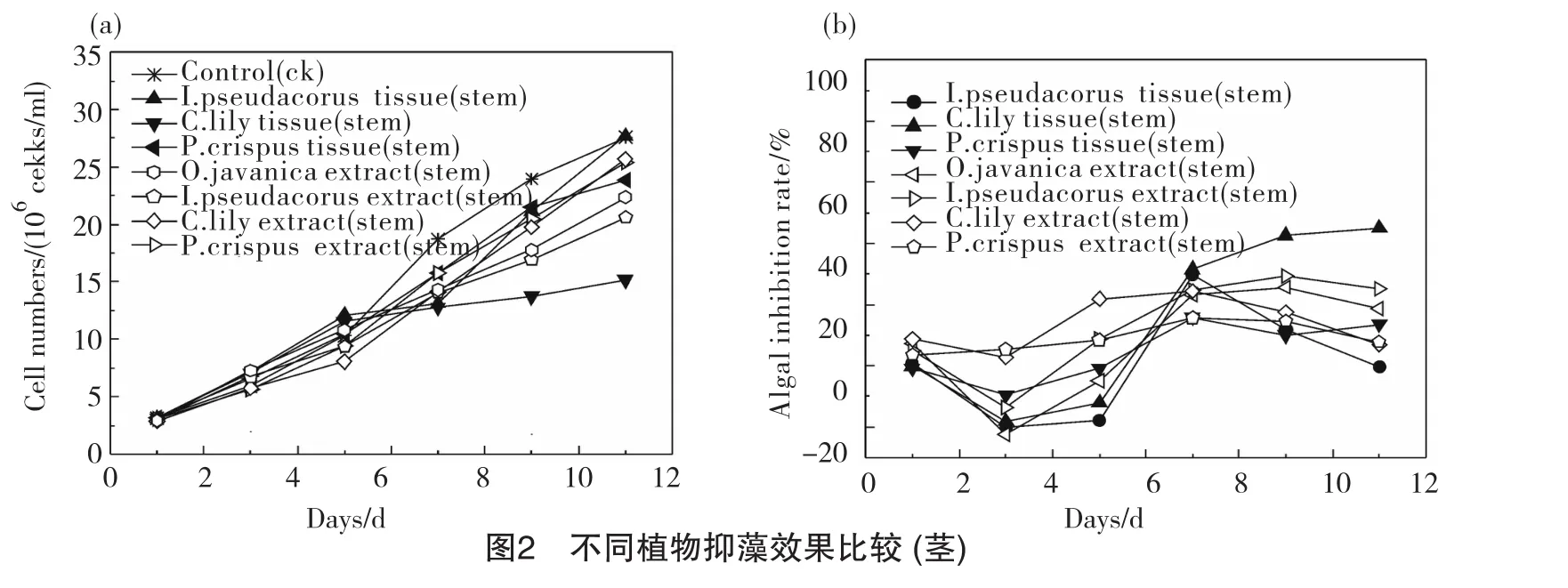
從圖3可以看出,植物葉組織提取物抑藻比葉組織更具優(yōu)勢,水芹、黃菖蒲、美人蕉、菹草4組的葉提取物對藻的抑制效果均好于葉組織組,特別是美人蕉葉提取物組,從試驗開始到結(jié)束,生物量均保持較低水平,11d時其生物量為6.75×106cell/ml,僅為對照組的1/4;水芹葉提取物組對藻的抑制率隨著時間的增加而增大,呈穩(wěn)定上升趨勢,3d、7d、11d時的抑制率分別為12.52%、 24.47%、56.96%;黃菖蒲葉提取物從第5d開始表現(xiàn)出抑藻效果,試驗結(jié)束時抑制率達(dá)到65.30%;相比之下,菹草葉提取物抑藻效果一般,抑制率為32.28%。植物葉組織組中,各植物葉組織對藻的抑制率從大到小依次為:黃菖蒲(31.71%)>水芹(15.86%)>菹草(8.09%)>美人蕉(-30.89%)。
在水生植物化感抑藻技術(shù)領(lǐng)域中,化感作用是一種重要的化學(xué)生態(tài)防御機制,是植物代謝活動對藻類所產(chǎn)生的不利作用[14]。植物與周圍的生物群落以次生代謝物質(zhì)為媒介建立了穩(wěn)固的化學(xué)作用關(guān)系,它們通過莖葉揮發(fā)、莖葉淋溶、根系分泌以及植物殘株的腐解等途徑向水體中釋放化感物質(zhì),來影響水體中藻類的生長[15]。研究和掌握這種化學(xué)作用規(guī)律,對認(rèn)識植物群落結(jié)構(gòu)、演替并合理開發(fā)和利用,實現(xiàn)藻類的有效控制,減少生態(tài)系統(tǒng)修復(fù)對化學(xué)藥劑的依賴具有重要意義。通過上述分析,可以發(fā)現(xiàn),植物組織及組織提取物的抑藻效果大不相同,同一種植物的相同器官在不同的處理方式下及同一處理方式下的不同植物,抑藻效果都有著強烈的反差,比如,黃菖蒲根組織對藻的抑制率為51.81%,提取物卻僅為16.43%;美人蕉葉提取物對藻的抑制率高達(dá)75.52%,葉組織不僅對藻沒有抑制效果反而促進(jìn)了藻的生長。一般認(rèn)為,水生植物的化感抑藻作用是多種化學(xué)物質(zhì)混合作用的結(jié)果,以上試驗結(jié)果的差異性可以歸結(jié)為不同植物、不同植物器官中以及不同處理方式下導(dǎo)致抑藻效應(yīng)的化學(xué)物質(zhì)種類及含量的不同[1,16,17]。因此,只有掌握了不同植物類型及處理方式下的抑藻潛力,才能在水體富營養(yǎng)化植物修復(fù)或水華生物控制工程中實現(xiàn)有效控藻。
2.2 累積效應(yīng)分析
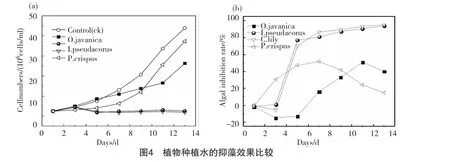
以上水生植物各器官的抑藻效果,在植株有機整體中最終以經(jīng)各種途徑釋放到水體中化感物質(zhì)的累積效應(yīng)表現(xiàn)出來。分別取4種植物35d的種植水進(jìn)行抑藻試驗,結(jié)果如圖4所示。從圖中可看出,黃菖蒲及美人蕉種植水的抑藻效果非常好,生物量從第3d開始明顯下降且保持較低水平,抑制率在第5d分別已經(jīng)達(dá)到77.25%和70.12%。水芹抑藻效果次之,試驗結(jié)束時,其對藻的抑制率為40.02%。菹草抑藻效果最差,該組藻生物量與對照組類似,呈指數(shù)增長模式;其抑制率隨著時間的增長先增大后減小,最高可達(dá)51.94%,出現(xiàn)在第7d,試驗結(jié)束時抑制率最低為15.05%。水生植物能連續(xù)釋放某些化感物質(zhì)抑制藻類的生長,但這些物質(zhì)的理化性質(zhì)不穩(wěn)定,在天然水體中容易被微生物所利用。當(dāng)化感物質(zhì)達(dá)到一定量時才表現(xiàn)為有效的抑藻效應(yīng),因此,水生植物的化感抑藻作用表現(xiàn)出明顯的累積性[18,19]。從以上結(jié)果可以看出,黃菖蒲、美人蕉種植水抑藻效果較為明顯,而水芹、菹草種植水抑藻效果較差,這說明同一種植條件下,相同時間范圍內(nèi),黃菖蒲、美人蕉兩種植物體系中有效化感物質(zhì)的累積效應(yīng)要明顯大于水芹及菹草。
2.3 不同植物抑藻特征分析
對藻生物量、增長率及生長拐點的把握對于藻的抑制有著一定的指示意義,本研究采用Logistic方程,以每個處理組的最大生物量(Nm)作為各自K的估計值,以ln[(K-Nt)/Nt]為y,時間t為x,以最小二乘法進(jìn)行回歸分析,獲得該方程的斜率和截距作為a和r的估計值。藻類的生長拐點即為Logistic方程的二階導(dǎo)數(shù)等于0的時間,由公式tp=(a-ln2)/r計算得出。各植物組織及組織提取物化感作用下銅綠微囊藻的logistic方程參數(shù)、判定系數(shù)及拐點時間見表1和表2。
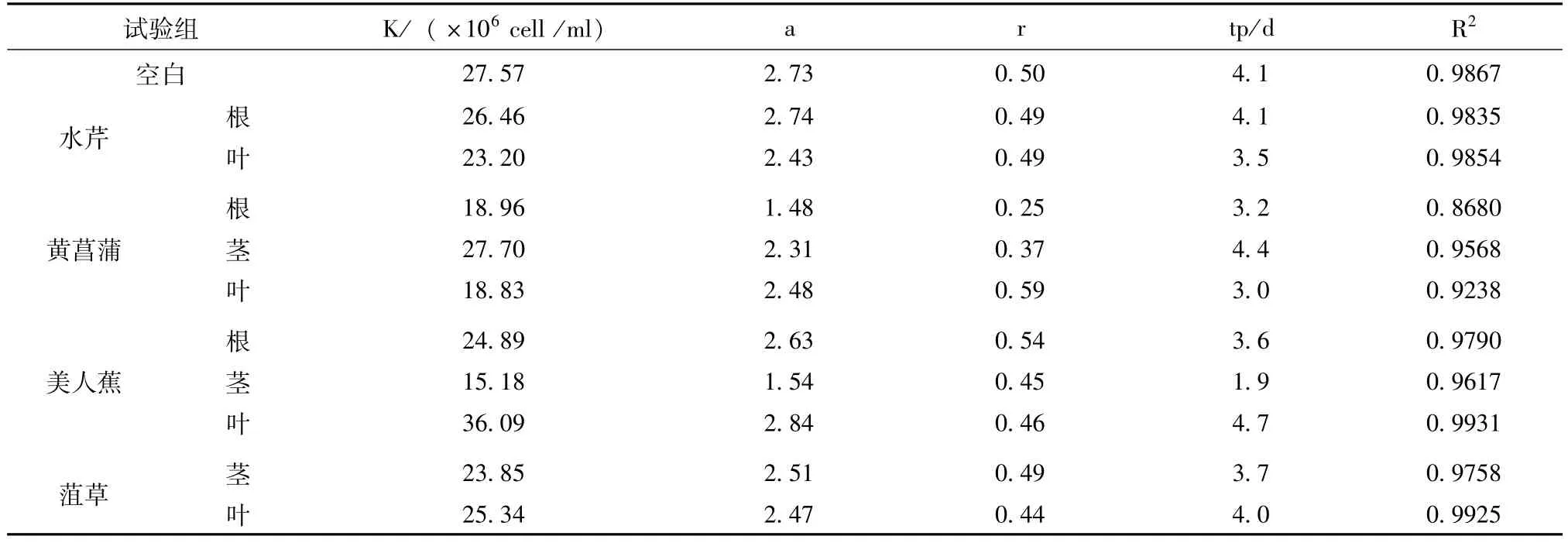
表1 培養(yǎng)條件下銅綠微囊藻的logistic方程參數(shù)、判定系數(shù)及拐點時間(植物組織)
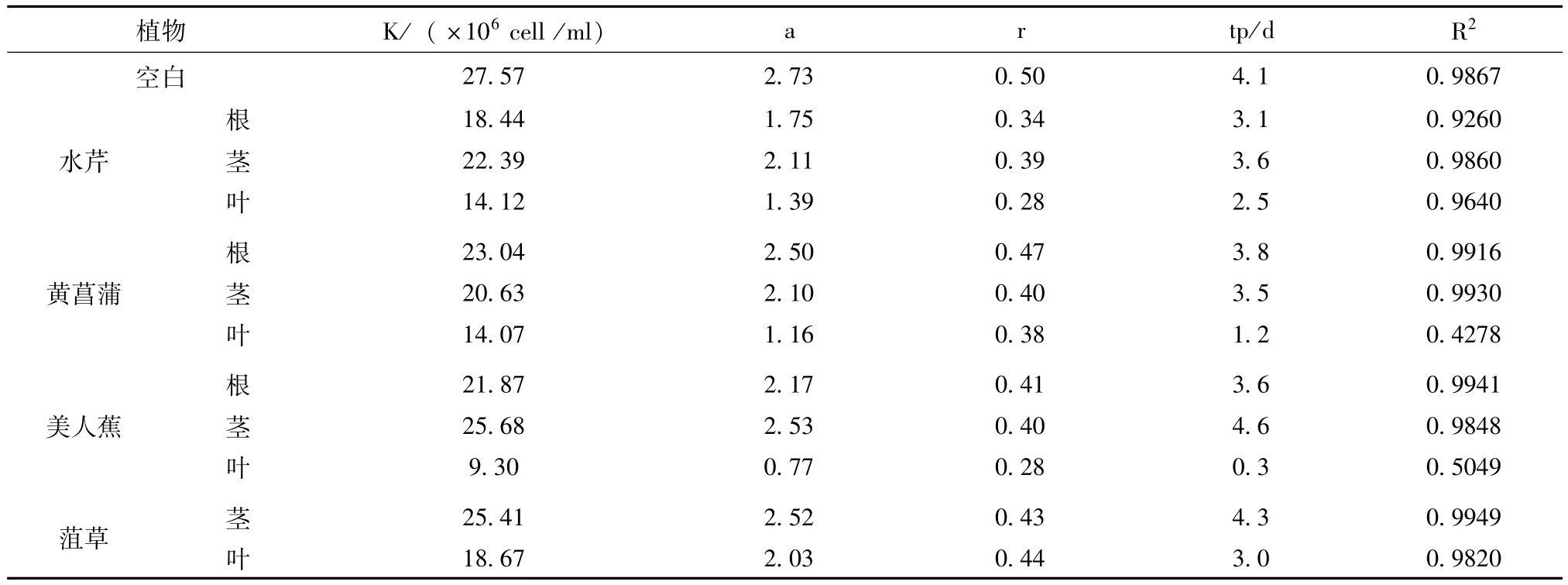
表2 培養(yǎng)條件下銅綠微囊藻的logistic方程參數(shù)、拐點出現(xiàn)時間(組織提取物)
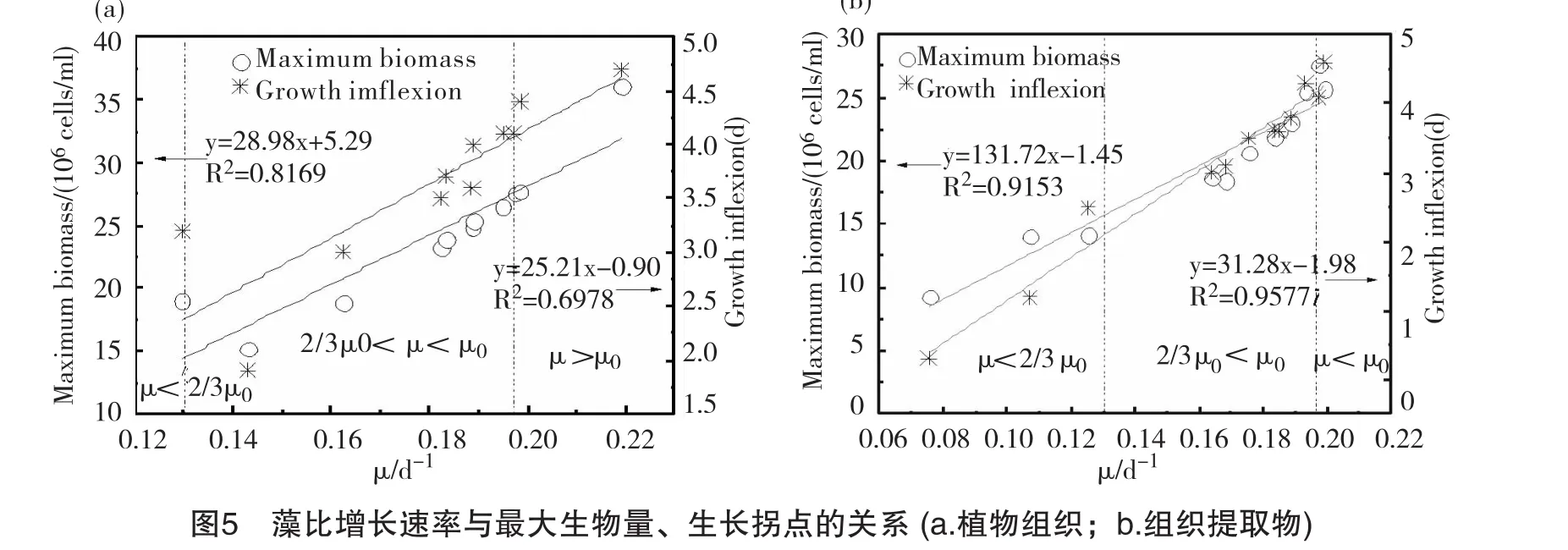
為了清晰地比較藻最大生物量、生長拐點及比增長率的關(guān)系,對以上K、tp參數(shù)及藻比增長率μ作圖,如圖5。從圖中可以看出,藻最大生物量及生長拐點時間與藻的比增長率有著較好的相關(guān)性,組織提取物組相關(guān)性較好,K與μ,tp與μ的線性擬合系數(shù)分別為0.9153、0.9577;相比之下,組織器官組相關(guān)性較差,從圖上可以看出,在比增長率為0.13/d時,藻最大生物量及生長拐點偏離擬合直線較遠(yuǎn)。總體而言,較好的抑藻作用表現(xiàn)為比增長速率低、生物量小、生長拐點時間早。以對照組生長速率μ0為參照標(biāo)準(zhǔn),將試驗范圍內(nèi)藻的生長參數(shù)按μ>μ0,μ0>μ>2/3μ0以及μ<2/3μ0劃分為3個區(qū)間,表示化感抑藻程度,判斷各組試驗的抑藻潛力。處于μ<2/3μ0區(qū)間的表現(xiàn)為抑藻效果最佳,也說明其對應(yīng)的植物組織及組織提取物最具抑藻潛力,順序分別為美人蕉葉提取物組>黃菖蒲葉提取物組>水芹葉提取物組>美人蕉莖組;其中提取物組生長拐點為0.3~2.5d,組織器官組為3.2d;處于μ0>μ>2/3μ0區(qū)間的抑藻效果次之,67%的試驗組處于這一區(qū)間;其中提取物組生長拐點為3.5~4.1d,組織器官組為1.9~4.1d;μ>μ0的為無抑藻效果,組織器官組與植物提取物組的生長拐點均>4.4d。從以上分析可以看出,組織提取物比植物組織組抑藻效果更好;另外,與根、莖器官相比,挺水植物葉的提取物更具抑藻潛力。
2.4 銅綠微囊藻化感受損機制
近幾年,“生物活性氧傷害”理論[20]在植物化感抑藻領(lǐng)域應(yīng)用較為廣泛,一般認(rèn)為,在正常的生理情況下,藻細(xì)胞中活性氧不斷產(chǎn)生的同時,也被不斷地清除,因而產(chǎn)生的自由基不會造成對藻細(xì)胞的損傷;但一旦當(dāng)藻細(xì)胞處于脅迫條件下,自由基的產(chǎn)生與清除失去平衡,活性氧含量會顯著增加。過量的氧自由基破壞脂體、蛋白、DNA燈,導(dǎo)致細(xì)胞內(nèi)抗氧化劑的減少,酶活性受到不同程度的影響[21]。在微囊藻細(xì)胞中,抗氧化酶主要包括超氧化物歧化酶(Superoxide Dismutase,SOD)、過氧化氫酶(Catalase,CAT)、過氧化物酶(Peroxidase,POD)等,其中SOD酶起主要作用;另外,SOD酶與CAT、POD酶的酶活性表達(dá)方式存在差異,正常生長的藻細(xì)胞,SOD酶活性較高,容易測定,而CAT、POD酶活性則相對較弱,因此本研究采用SOD酶為主要測試指標(biāo),研究銅綠微囊藻的化感受損機制。
圖6為植物組織及提取物對銅綠微囊藻SOD酶活性的影響趨勢。從圖中可以看出,對照組的SOD酶活性前期一直保持較低水平,說明對照組的藻類生長狀況良好,沒有受到外界條件的脅迫;到試驗結(jié)束時 (11d),對照組由于生物量的巨增導(dǎo)致遮光效應(yīng)明顯,使得藻類所受環(huán)境脅迫增強,SOD酶略有升高(1.14U10-6/cells),但依舊低于其他大部分試驗組。水芹、黃菖蒲、美人蕉及菹草4種植物的組織器官及組織提取物對藻的化感作用不同,因此在藻SOD酶活性大小上表現(xiàn)為較大差異,且隨著作用時間的變化呈現(xiàn)不同的變化趨勢。至試驗結(jié)束時,植物組織組中SOD酶活性最大的是黃菖蒲根組,其SOD值為2.27 U 10-6/cells;最小的是美人蕉葉組,其SOD值為0.77 U 10-6/cells。提取物組中SOD酶活性最大的是美人蕉葉組,其SOD值為4.30 U10-6/cells;最小的是美人蕉根組,其SOD值為0.95 U 10-6/cells。
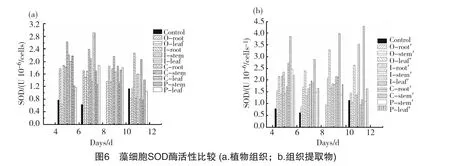
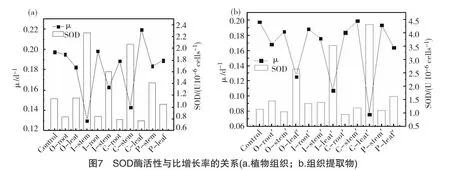
藻細(xì)胞超氧化物歧化酶 (SOD)在生物體清除超氧化物陰離子自由基 ()過程中起著重要作用,是細(xì)胞抗衡脅迫的主要保護(hù)酶[22]。在抑藻試驗體系中,SOD酶的增大是微囊藻在生長環(huán)境中受到了培養(yǎng)基中化感物質(zhì)脅迫時的應(yīng)激反應(yīng);當(dāng)增大到某一值時,SOD酶活性有下降趨勢,可能是SOD酶不能與O2·-在更高的水平上建立新的平衡,表明此時藻細(xì)胞已經(jīng)受到了較大程度的損傷[23]。將試驗結(jié)束時各組中SOD酶活性與藻比增長率進(jìn)行比較分析,從圖7可以發(fā)現(xiàn),在植物組織及組織提取物組,SOD酶活性與藻的比增長率均呈現(xiàn)較好的負(fù)相關(guān)關(guān)系,即藻細(xì)胞內(nèi)SOD酶活性較大時,比增長率較低;反之較高。
3 結(jié)論
不同植物種植水對銅綠微囊藻的生長抑制影響差異很大,主要是因為種植系統(tǒng)中化感物質(zhì)的積累效應(yīng)不同,抑藻效果順序為美人蕉、黃菖蒲>水芹>菹草。不同的植物組織器官及不同的植物處理方式下,抑藻效果不同,最具抑藻潛力的幾組為美人蕉葉提取物組>黃菖蒲葉提取物組>水芹葉提取物組>美人蕉莖組,具體表現(xiàn)為比藻增長速率低、生物量小、生長拐點時間早。抑藻機理可能在于化感物質(zhì)的脅迫使微囊藻細(xì)胞內(nèi)活性氧自由基過量堆積,進(jìn)而引起對藻細(xì)胞的傷害和影響藻類正常的光合作用;SOD酶活性與藻的比增長率均呈現(xiàn)較好的負(fù)相關(guān)關(guān)系,即藻細(xì)胞內(nèi)SOD酶活性較大時,比增長率較低,反之較高。
[1]DaiW,Zhang S L,Lin Y J,et al.Allelopathic Effects of Berberine,a Plant Alkaloid,on the Algae,Microcystis aeruginosa(FACHB-905),at Different Initial Densities[J].Israeli Journal of Aquaculture-Bamidgeh,2013,(65):1-5.
[2]Ferrier M D,Butler B R,Terlizzi D E,et al.The effects of barley straw(Hordeum vulgare)on the growth of freshwater algae[J]. Bioresource Technology,2005,96(16):1788-1795.
[3]Xiao X,Chen Y-x,Liang X-q,et al.Effects of Tibetan hulless barley on bloom-forming cyanobacterium(Microcystis aeruginosa)measured by different physiological and morphologic parameters[J].Chemosphere,2010,81(9):1118-1123.
[4]Ball A S,Williams M,Vincent D,et al.Algal growth control by a barley straw extract[J].Bioresource Technology,2001,77(2):177-181.
[5]Park M H,Hwang SJ,Ahn C Y,etal.Screening of seventeen oak extracts for the growth inhibition of the cyanobacterium Microcystis aeruginosa Kutz.em.Elenkin[J].Bulletin of Environmental Contamination and Toxicology,2006,77(1):9-14.
[6]Park M-H,Kim B-H,Chung I-M,et al.Selective Bactericidal Potential of Rice(Oryza sativa L.var.japonica)Hull Extracton Microcystis Strains in Comparison with Green Algae and Zooplankton[J].Bulletin of Environmental Contamination and Toxicology,2009,83(1):97-101.
[7]Chen JZ,Liu Z L,Ren G J,etal.Control ofMicrocystisaeruginosa TH01109 with batangas mandarin skin and dwarf banana peel[J].Water Sa,2004,30(2):279-282.
[8]Chen J,Zhang H,Han Z,et al.The influence of aquatic macrophytes on Microcystis aeruginosa growth[J].Ecological Engineering,2012,(42):130-133.
[9]胡光濟(jì),張維昊,尚吟竹,等.菖蒲干體提取液對兩種水華藻類生長的影響[J].應(yīng)用生態(tài)學(xué)報,2009,(9):2277-2282.
[10]Wu X,Wu H,Chen J,et al.Effects of allelochemical extracted from water lettuce(Pistia stratiotes Linn.)on the growth,microcystin production and release ofMicrocystis aeruginosa[J].Environmental Science and Pollution Research,2013,20(11):8192-8201.
[11]Chang X,Eigemann F,and Hilt S.Domacrophytes supportharmful cyanobacteria Interactions with a green alga reverse the inhibiting effectsofmacrophyteallelochemicalson Microcystisaeruginosa[J].Harmful Algae,2012,(19):76-84.
[12]Mulderij G,MooijW M,Smolders A JP,etal.Allelopathic inhibition of phytoplankton by exudates from Stratiotes aloides[J].A-quatic Botany,2005,82(4):284-296.
[13]Chen J,Zhang H,Han Z,et al.The influence of aquatic macrophytes on Microcystis aeruginosa growth[J].Ecological Engineering,2012,(42):130-133.
[14]Nakai S,HosomiM.Allelopathic inhibitory effectsof polyphenols released by Myriophyllum spicatum on algal growth[J].Allelopathy Journal,2002,10(2):123-131.
[15]Nakai S,Zou G,Okuda T,etal.Anti-cyanobacterialallelopathic effectsof plants used for artificial floating islands[J].Allelopathy Journal,2010,26(1):113-121.
[16]Huang Y,Bai Y,Wang Y,et al.Allelopathic effects of the extracts from an invasive species Solidago canadensis L.on Microcystis aeruginosa[J].Letters in Applied Microbiology,2013,57(5):451-458.
[17]Zhang T T,He M,Wu A P,et al.Allelopathic effects of submerged macrophyte Chara vulgaris on toxic Microcystis aeruginosa[J].Allelopathy Journal,2009,23(2):391-401.
[18]Wu Z B,Gao Y N,Wang J,etal.Allelopathic effects of phenolic compounds present in submerged macrophytes on Microcystis aeruginosa[J].Allelopathy Journal,2009,23(2):403-410.
[19]Wang H Q,Cheng SP,Zhang SH,etal.Chemical Composition in Aqueous Extracts of Potamogeton malaianus and Potamogeton maackianus and their Allelopathic Effects on Microcystis aeruginosa[J].Polish Journal of Environmental Studies,2010,19(1):213-218.
[20]Mccord JM F I.Superoxide Dismutase-An Enzymic Function for Erythrocuprein(Hemocuprein)[J].JBiol chem,1969,(244):6049.
[21]Collen J,Davison IR.Reactive oxygen production and damage in intertidal Fucus spp.(Phaeophyceae)[J].Journal of Phycology,1999,35(1):54-61.
[22]Zhang S,Zhang B,DaiW,et al.Oxidative damage and antioxidant responses in Microcystisaeruginosa exposed to the allelochemical berberine isolated from golden thread[J].Journal of Plant Physiology,2011,168(7):639-643.
[23]Liu Y,Li F,Huang Q X.Allelopathic effects of gallic acid from Aegiceras corniculatum on Cyclotella caspia[J].Journal of Environmental Sciences-China,2013,25(4):776-784.
Research of Effects and M echanism on Algae Control by Different Aquatic M acrophytes
SU Yue-long1,WANG Zheng-fang2
(Yunlong Environmental Monitoring Station,Yunlong Yunnan 672700 China)
The allelopathic effects of four aquatic macrophytes(Iris pseudacorus,Canna indica,OenantheJavanica,and Potamogeton)on the growth of Microcystis aeruginosa were investigated.The allelopathic effectswere different by different organs and various pretreatment.Iris pseudacorus and Canna indica were found to have themost potentialities of controlling algae.The water after culturing Iris pseudacorus and Canna indica were used to culture algae.The inhibition rate of algae could reach 70.12%to 77.25%five days later.The liquid extracted from the leaves of Irispseudacorus with 0.53 percentage ofmass fraction was applied to culture algae for11 days.The inhibition rate was 65.30%.The allelopathic mechanism could be explained by the stress fromthe allelopathic materials that damaged and hindered the photosynthesis of algae through improving the accumulation of active oxygen radicals in the cells of algae.In addition,the activity of superoxide dismutase(SOD)showed a good negative correlation with the growth of Microcystis aeruginosa.
aquatic macrophyte;microcystis aeruginosa;superoxide dismutase;algae control;allelopathy
X17
A
1673-9655(2014)06-0015-08
2014-08-20
猜你喜歡
中老年保健(2021年12期)2021-11-30 02:58:01
攝影之友(影像視覺)(2019年2期)2019-03-05 08:27:14
中華詩詞(2018年11期)2018-03-26 06:41:34
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
Coco薇(2016年8期)2016-10-09 02:11:50
少兒科學(xué)周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學(xué)周刊·兒童版(2015年1期)2015-07-07 04:12:52
