黃姑魚仔稚魚發育過程中氨基酸和脂肪酸的變化
2014-07-12 08:19:18金煜華謝中國史會來
浙江海洋大學學報(自然科學版) 2014年1期
金煜華,謝中國,樓 寶,史會來
(1.浙江省海洋水產研究所,浙江省海水增養殖重點實驗室,浙江舟山 316021;2.浙江海洋學院水產學院,浙江舟山 316022;3.浙江舟山普陀海洋高科技園,浙江舟山 316102;4.湖南文理學院生命科學學院,湖南常德 415000)
黃姑魚Nibea albiflora Richardson俗稱黃婆雞,屬鱸形目、石首魚科、黃姑魚屬,具有肉味鮮美、營養豐富、養殖效益高等特點,是我國的重要海水經濟魚類。近年來陸續開展了黃姑魚生物學、全人工繁殖及大規格魚種培育技術研究等工作[1,2]。傳統的黃姑魚苗培育一般采用輪蟲、鹵蟲和橈足類等生物活餌,但生物餌料成本昂貴,營養不全面,產量不穩定且易攜帶病原菌,育苗成活率僅為10%左右,嚴重制約著黃姑魚產業的發展。研制適用于黃姑魚仔稚魚的人工配合飼料部分或全部代替生物餌料是解決這一問題的迫切需求。由于對黃姑魚仔稚魚營養需求了解十分有限,因此難以配置出營養均衡全面的飼料。研究黃姑魚仔稚魚發育過程中氨基酸、脂肪酸的變化,以期為開發優質高效的仔稚魚微粒飼料提供理論基礎。
1 材料與方法
1.1 試驗材料
實驗于2012年6-7月在浙江省海洋水產研究所西閃試驗場進行,實驗所用黃姑魚親魚為該試驗場人工繁育的體質健壯、無病無傷的2年齡黃姑魚。經人工催產,收集受精卵于5 m×2 m×2 m的育苗池中充氣培養,育苗所用水為經沉淀和沙濾后的天然海水,鹽度26.5~27.0。受精卵孵化期間水溫27±1℃,經30 h左右孵出;仔稚幼魚在原池中培育,水溫27±2℃。
初孵仔魚2日齡開口攝食,開口餌料為經過小球藻強化培育的褶皺臂尾輪蟲,10日齡左右投喂鹵蟲無節幼體,12日齡開始投喂橈足類或人工配合飼料。人工育苗餌料系列及使用時間如圖1所示。
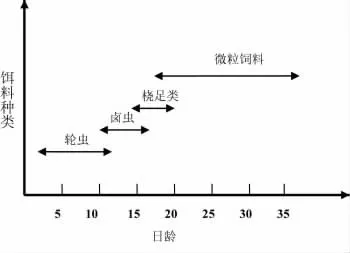
圖1 黃姑魚人工育苗餌料系列及使用時期Fig.1 Feeding regime of the N.albiflora larvae
1.2 樣品采集
實驗所取樣品為1、5、10、20、45日齡的仔稚魚,取樣時間均在早晨投喂飼料前,樣品取全魚,隨機取出的仔稚魚放入無餌料水族箱內1 h,20、45日齡稚魚饑餓24 h。然后將所取的仔稚魚置于冰盤上致死。所得的樣品放在-80℃冰箱保存。仔稚魚樣品在氨基酸、游離氨基酸、脂肪酸分析前,用美國電熱YO-230冷干機經充分冷凍干燥至恒重。
1.3 氨基酸分析
樣品用6 mol/L的鹽酸溶液在110℃下水解22 h,然后過濾水解液,用超純水定容后取2 mL減壓蒸干,干燥完后加入0.02 mol/L的鹽酸溶液2 mL溶解全部氨基酸,上機進行色層分析。用美國安捷倫Agilent 1100全自動氨基酸分析儀測定樣品中的的氨基酸組成。
1.4 游離氨基酸分析
準確稱取1.0 g樣品,加15 mL 5%三氯乙酸溶液勻漿,定容至25 mL,于4℃放置24 h。用定性濾紙過濾吸取0.1 mL在10 000 r/min下離心10 min,于4℃保存待檢測用。用美國安捷倫Agilent 1100全自動氨基酸分析儀測定樣品中游離氨基酸。
1.5 脂肪酸分析
檢測方法采用GB/21514—2008,使用日本島津GC-2010氣相色譜儀測定。
1.6 數據處理
實驗所得數據采用SPSS19.0軟件處理,做單因素方差(One-way ANOVA)分析,若差異顯著(P<0.05)則進行Duncan多重比較。結果數據均用平均值±標準差(mean±SD,n=3)表示。
2 結果與討論
2.1 黃姑魚仔稚魚氨基酸變化模式
各日齡黃姑魚仔稚魚所含必需氨基酸與非必需氨基酸的變化模式見表1。不同日齡的黃姑魚水解氨基酸中,必需氨基酸中的精氨酸、纈氨酸、亮氨酸、異亮氨酸、賴氨酸5種氨基酸的含量最高,占必需氨基酸總量的70%左右。必需氨基酸中的組氨酸、蛋氨酸含量最低,僅占必需氨基酸的10%左右。水解氨基酸的總量呈上升趨勢,必需氨基酸的總量,先下降,后上升。從不同日齡的氨基酸含量來看,賴氨酸為第一限制性氨基酸。非必需氨基酸中,天冬氨酸、谷氨酸和丙氨酸的含量最高,占非必需氨基酸總量的60%左右。隨發育階段不同,仔稚魚的各種氨基酸含量差異較大。
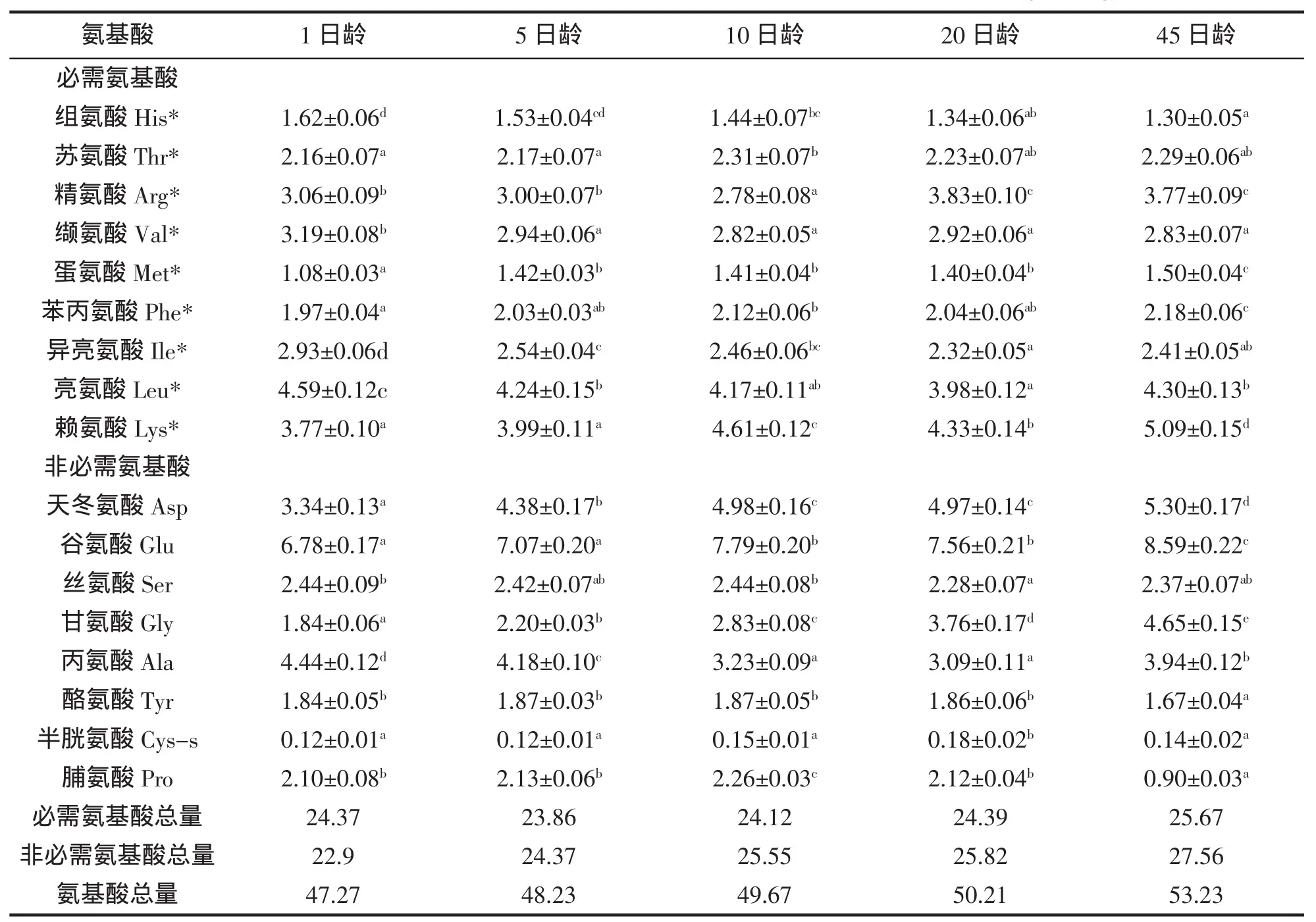
表1 黃姑魚仔稚魚氨基酸的含量(g/100 g)Tab.1 Contents and compositions of amino acids of N.albiflora larvae(g/100 g)
對仔稚魚飼料而言,任何一種必需氨基酸(EAA)的缺乏或者不足均會提高氨基酸的氧化代謝,導致魚類生長性能下降和飼料轉化率的降低[3-4]。因此,了解仔稚魚對每種必需氨基酸的特定需求量對能否配制出營養平衡且成本較低的飼料至關重要。海水仔稚魚具有較高的生長率[5],因而對氨基酸有較高的需要量,以滿足其體蛋白的沉積、更迭、能量代謝及其它生理功能。海水仔稚魚的必需氨基酸通常認為與已經確定的魚類的必需氨基酸一致,為蛋氨酸、賴氨酸、精氨酸、組氨酸、亮氨酸、異亮氨酸、苯丙氨酸、蘇氨酸、色氨酸、纈氨酸10種氨基酸。在實踐中采用魚肉氨基酸組成指標作為魚類必需氨基酸需要量能較好地反映魚類氨基酸需要量,理想的飼料氨基酸模式應該是仔稚魚的體組成氨基酸模式[6]。與幼成魚相比,由于不同時期仔稚魚器官和組織的發育速度不同,從而導致仔稚魚體的氨基酸組型隨發育階段不同而變化較大。ARAG?O等[3]的研究結果表明半滑舌鰨仔稚魚的氨基酸組型在發育過程不斷變化,必需氨基酸的需要量同時發生變化,很難制備能夠完全滿足仔稚魚各個時期氨基酸需要的平衡飼料。飼料中氨基酸的平衡能提高氨基酸的轉化率,促進仔稚魚的生長以及對蛋白質的利用效率。隨著仔稚魚營養研究的進一步深入,越來越多的研究結果表明仔稚魚具有較強的調節氨基酸代謝能力。由于傳統方法難以推測仔稚魚的氨基酸需要量,近年來不少科研工作者采用同位素標記的方法來研究仔稚魚的氨基酸需要量[7-9]。
2.2 黃姑魚仔稚魚游離氨基酸變化模式
黃姑魚仔稚魚發育階段的游離氨基酸含量見表2。1日齡的仔稚魚游離氨基酸含量最高,為12.134 g/100 g;45日齡仔稚魚的游離氨基酸含量最低,為1.497 g/100 g。隨著仔稚魚的生長,游離氨基酸的含量呈降低趨勢,20日齡的黃姑魚仔稚魚游離氨基酸含量略有升高,推斷可能跟攝食的餌料鹵蟲有關。隨著仔稚魚發育階段不同,各種游離氨基酸含量差異比較顯著。
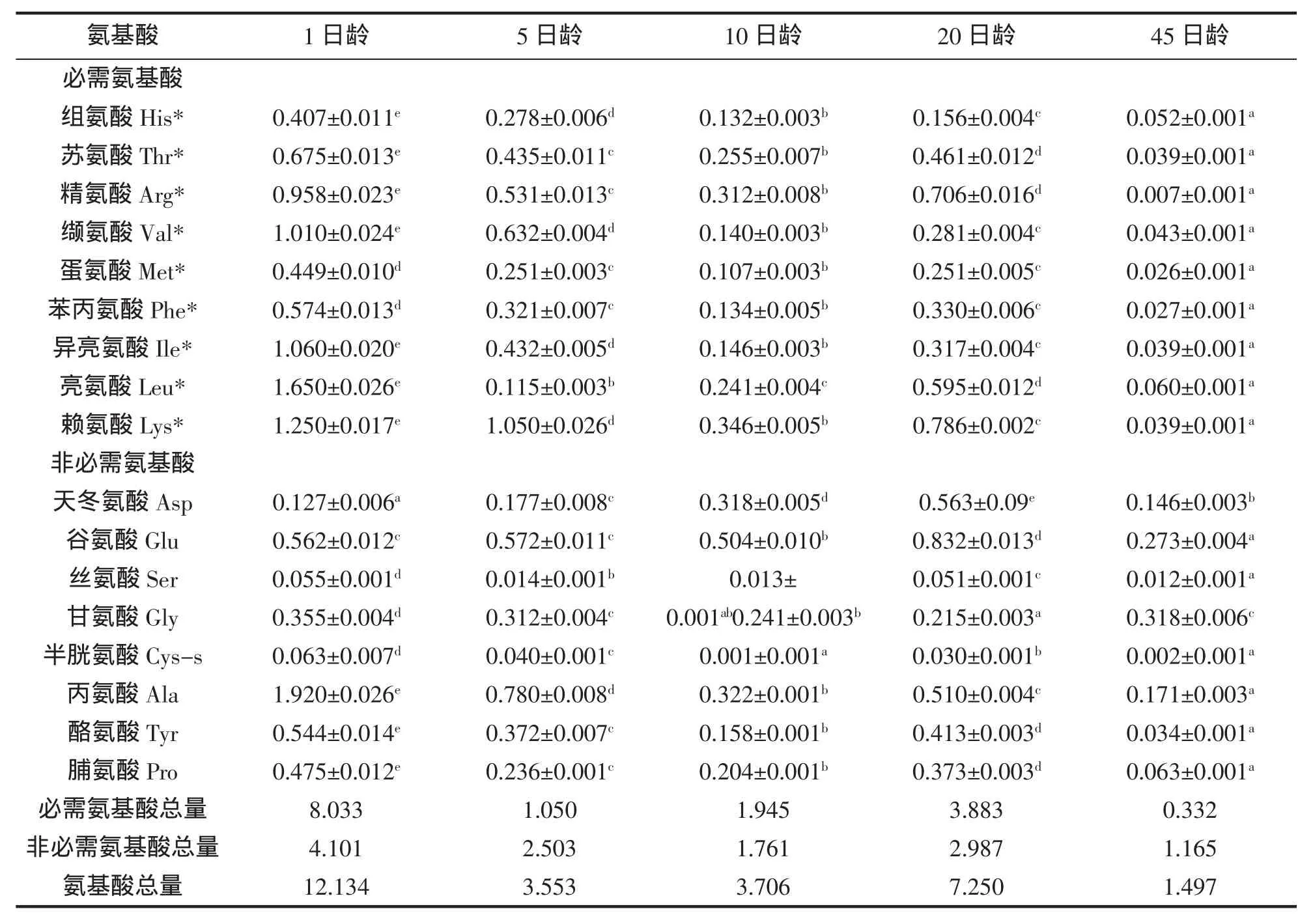
表2 黃姑魚仔稚魚游離氨基酸的含量(g/100 g)Tab.2 Contents and compositions of free amino acids of N.albiflora larvae(g/100 g)
游離氨基酸(FAA)既是重要的代謝供能物質,也是魚體蛋白質生物合成的主要底物。游離氨基酸在海水魚魚卵成熟過程及孵化早期發揮著重要的作用,研究發現游離氨基酸可以作為鮭魚Gadus morhua卵和仔稚魚發育的能量源[10]。R?NNESTAD等[4]認為海水魚類的胃在仔稚魚時期尚未成功分化,導致仔稚魚攝食的蛋白質無法高效率的變性、降解,而蛋白質的降解產物小分子肽類物質或游離氨基酸相對于蛋白質來說更容易被仔稚魚吸收和利用,科研工作者已逐漸重視游離氨基酸及小肽的營養作用。將14C標記的游離氨基酸精氨酸、賴氨酸添加到飼料,采用管飼技術飼喂大西洋鮭Hippoglossus hippoglossus仔稚魚,30 min后發現腸道的平均吸收率高達71%,而排出率僅為6%[11]。海水仔稚魚攝食生物活餌后,游離氨基酸迅速釋放被仔稚魚吸收利用,因此推測游離氨基酸在魚類消化系統發育不完善的早期階段發揮著重要的生理作用。研究證實,飼料中蛋白的可溶性以及肽的構型會極大程度地影響仔魚的生長[12]。游離氨基酸中的甘氨酸、丙氨酸和精氨酸可作為仔稚魚的誘食劑[13]。在仔稚魚飼料中添加過量的游離氨基酸,其效果往往會適得其反。受限于魚體體內有限的轉運載體,大量的游離氨基酸不能被吸收利用,而是通過鰓、泌尿系統等途徑排出體外,不會被用于蛋白質合成以促進生長。
2.3 黃姑魚仔稚魚脂肪酸組成
黃姑魚仔稚魚體脂肪酸的變化模式見表3。黃姑魚仔稚魚發育過程的主要脂肪酸為:16:0、18:0、16:1、18:1和DHA,其總量占脂肪酸總量的70%左右。仔稚魚必需的脂肪酸DHA、EPA、ARA在仔稚魚的發育過程中含量變化均較為顯著,但均處于較高的含量水平。飼料脂肪酸組成直接影響到養殖對象脂肪酸組成和含量,由于不同日齡仔稚魚攝食不同的餌料,而不同餌料的脂肪酸組成不同,從而造成不同日齡仔稚魚的脂肪酸組成差異較大。
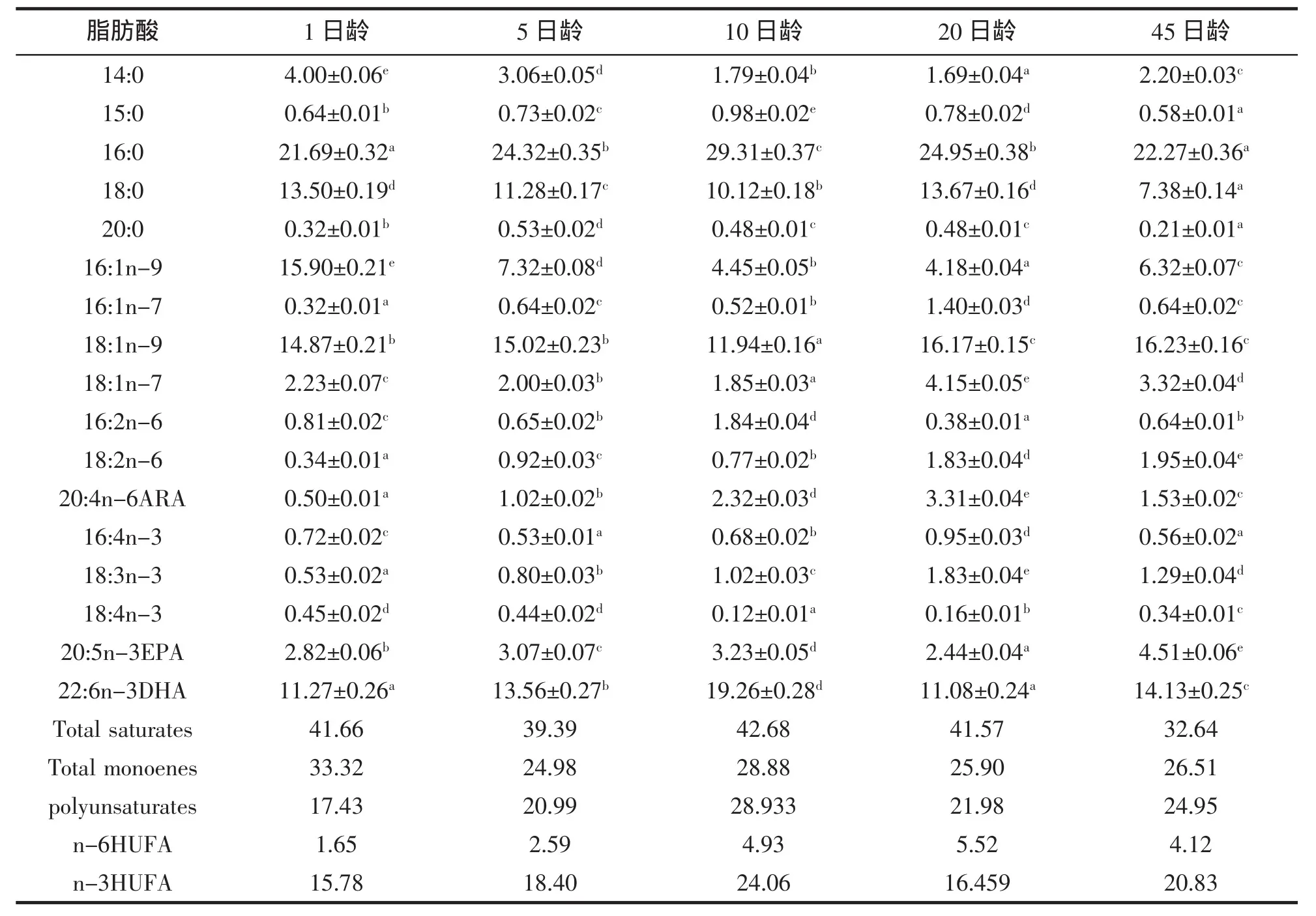
表3 黃姑魚仔稚魚脂肪酸組成(占總脂肪酸的質量百分比)Tab.3 Contents and compositions of fatty acids of the N.albiflora larvae(weight percentage of total fatty acids)
脂類是魚類胚胎發育過程中重要的代謝能源,同時為海洋魚類提供必需脂肪酸。對海水魚魚卵的脂肪酸組成分析發現,DHA和EPA在總脂肪酸中所占比重非常高,并且這種高水平一直持續于胚胎發育的全過程中,說明這兩種脂肪酸在胚胎發育的過程中發揮著重要的作用。特別是n-3系列高度不飽和脂肪酸HUFA能影響水產動物親體的繁殖力、受精率以及仔魚生長和存活率[14-16]。n-3HUFA如DHA、EPA是海水仔稚魚的必需脂肪酸,必須從飼料中獲得[17-18]。n-6HUFA如ARA的作用也越來越得到重視,ARA對海水仔稚魚的生長、滲透壓調節、存活率、魚體脂肪酸組成、繁殖、變態過程、體色、免疫力等方面同樣發揮著重要的影響[19]。仔稚魚脂肪酸的研究方向由分析單獨的脂肪酸營養成分向研究脂肪酸整體營養成分發展,考慮各種營養成分合適的配比,RAINUZZO等[20]發現DHA和EPA的比例平衡對于維持生物膜的流動性極為重要,一旦兩者比例失衡,尤其當EPA高而DHA低時,會使磷脂結構的組成不平衡,從而導致仔稚魚不能正常生長。海水魚卵中脂類的含量和組成相當于仔、稚魚的脂類需要,故對于開始外源性攝食的仔魚而言,理想餌料的脂肪酸組型應是與卵或卵黃囊仔魚的脂肪酸組型相近[21-22]。飼料脂肪酸組成直接影響到養殖對象脂肪酸組成已被大量的文獻報道所證實。由于本研究中不同日齡的仔稚魚攝食的餌料脂肪酸組成不同,這也是造成魚體脂肪酸組成差異的直接原因。
3 結論
黃姑魚仔稚魚發育過程中,由于各器官和組織的發育速度不同,不同餌料之間的氨基酸組型存在差異,導致黃姑魚仔稚魚的氨基酸組型是不斷變化的。必需氨基酸中的精氨酸、纈氨酸、亮氨酸、異亮氨酸、賴氨酸5種氨基酸的含量最高。1日齡的黃姑魚仔稚魚的游離氨基酸含量最高,隨后游離氨基酸的含量呈降低趨勢。黃姑魚仔稚魚的主要脂肪酸為:16:0、18:0、16:1、18:1和DHA。仔稚魚必需的脂肪酸DHA、EPA、ARA均處于較高的含量。
[1]雷霽霖,陳 超,徐延康,等.黃姑魚工廠化育苗技術研究[J].海洋科學,1992,11(6):5-10.
[2]樓 寶,史會來,毛國民,等.黃姑魚全人工繁育及大規格苗種培育技術研究[J].現代漁業信息,2011,26(3):20-23.
[3]ARAG?O C,CONCEIC?O L E C,FYHN H,et al.Estimated amino acid requirements during early ontogeny in fish with different life styles:gilthead seabream(Sparus aurata)and Senegalese sole(Solea senegalensis)[J].Aquaculture,2004,242:589-605.
[4]RφNNESTAD I,THORSEN A,FINN R N.Fish larval nutrition:recent advances in amino acid metabolism[J].Aquaculture,1999,177:201-216.
[5]DABROWSKI K.Ontogenetical aspects of nutritional requirements in fish[J].Comp Biochem Physiol,1986,85:639-655.
[6]MAMBRINI M,KAUSHIK S J.Indispensable amino acid requirements of fish:correspondence between quantitative data and amino acid profiles of tissue proteins[J].Journal of Applied Ichthyology,1995,11:240-247.
[7]R?NNESTAD I,ROJAS-GARCA C R,TONHEIM S,et al.In vivo studies of digestion and nutrient assimilation in marine fish larvae[J].Aquaculture,2001,201:161-175.
[8]HOVDE S C,VIDAL M C,OPSTAD I,et al.Design and synthesis of 14C-labeelled proteins as tools for protein digestion studies in fish larvae[J].Aquaculture Nutrition,2005,11:395-401.
[9]LE VAY L,GAMBOA-DELGADO J.Naturally-occurring stable isotopes as direct measures of larval feeding efficiency,nutrient incorporation and turnover[J].Aquaculture,2011,315:95-103.
[10]FYHN H J,SERIGSTAD B.Free amino acids as energy substrate in developing eggs and larvae of the cod Gadus morhua[J].Marine Biology,1987,296:335-341.
[11]APPLEBAUM S L,RφNNESTAD I.Absorption,assimilation and catabolism of individual free amino acids by larval Atlantic halibut(Hippoglossus hippoglossus)[J].Aquaculture,2004,230:313-322.
[12]CARVALHO A P,SA R,OLIVA TELES A,et al.Solubility and peptide profile affect the utilization of dietary protein by common carp(Cyprinus carpio)during early larval stages[J].Aquaculture,2004,234:319-333.
[13]薛 敏,解綬啟,崔奕波,等.魚類促攝食物質研究進展[J].水生生物學報,2003,27(6):639-643.
[14]IZQUIERDO M S,SOCORRO J,ARANTZAMENDI L,et al.Recent advances in lipid nutrition in fish larvae[J].Fish Physiology and Biochemistry,2000,22:97-107.
[15]KOVEN W M,TANDLER A,KISSIL G W,et al.The effect of dietary n-3 highly unsaturated fatty acids on growth,survival and swim bladder development in Sparus aurata larvae[J].Aquaculture,1990,91:131-141.
[16]SALHI M,IZQUIERDO M S,HERNáNDEZ-CRUZ C M,et al.Effect of lipid and n-3 HUFA levels in microdiets on growth survival and fatty acid composition of larval gilthead seabream(Sparus aurata)[J].Aquaculture,1994,124:275-282.
[17]劉鏡恪.海魚早期階段必需脂肪酸和磷脂的研究現狀與展望[J].海洋水產研究,2002,23(2):58-64.
[18]IZQUIERDO M S.Essential fatty acid requirements of cultured marine fish larvae[J].Aquaculture Nutrition,1996,2:183-191.
[19]KOVEN W,ANHOLT R V,LUTZKY S,et al.The effect of dietary arachidonic acid on growth,survival,and cortisol levels in different-age gilthead seabream larvae(Sparus auratus)exposed to handling or daily salinity change[J].Aquaculture,2003,228:307-320.
[20]RAINUZZO J R,REITAN K I,OLSEN Y.The significance of lipids at early stages of marine fish:A review[J].Aquaculture,1997,155:103-115.
[21]SARGENT J R,BELL J G,MCEVOY L A,et al.Recent development s in the essential fatty acid nutrition of fish[J].Aquaculture,1999,177:191-199.
[22]RAINUZZO J R,REITAN K I,OLSEN Y.The significance of lipids at early stages of marine fish:A review[J].Aquaculture,1997,155:103-115.
