CCK-8及其受體拮抗劑對嗎啡戒斷大鼠額葉皮質及海馬CREB與pCREB表達的影響*
2014-08-08 09:46:54高平蕊馬興友楊勝昌倪志宇李淑瑾馬春玲
中國病理生理雜志 2014年7期
關鍵詞:海馬
高平蕊, 馬興友, 文 迪, 楊勝昌, 于 峰, 倪志宇, 李淑瑾, 馬春玲, 叢 斌
(河北醫科大學基礎醫學院法醫學系, 河北省法醫學重點實驗室,河北 石家莊 050017)
八肽膽囊收縮素(cholecystokinin octapeptide,CCK-8)是迄今體內最強的抗阿片肽類物質[1]。有研究證明,CCK 受體與內源性阿片肽受體在中樞神經系統的分布密切相關,參與調節疼痛、焦慮等情緒和記憶過程[2],且在阿片成癮過程中也發揮著重要作用。阿片成癮是一種長時程的生物學效應,轉錄因子cAMP反應元件結合蛋白(cAMP response element binding protein, CREB)在此過程中具有重要的作用,并且磷酸化CREB(pCREB)對下游靶基因轉錄過程的調控作用可能是嗎啡依賴長時程適應性改變的基礎之一。本研究通過觀察CCK-8及CCK受體拮抗劑對嗎啡成癮大鼠額葉皮質和海馬CREB及pCREB的影響,探討CCK-8 在嗎啡成癮及戒斷過程中的分子機制,為CCK-8 在戒毒方面的應用提供相關實驗依據。
材 料 和 方 法
1 動物與試劑
成年雄性 Wistar 大鼠 30 只,體質量(200±10) g,由河北省實驗動物中心提供(實驗動物質量合格證為901014)。鹽酸嗎啡(沈陽第一制藥廠);鹽酸納絡酮和CCK-8(Sigma);L-364718和LY-288513(Tocris);CREB、pCREB和β-actin兔抗大鼠單克隆抗體(Cell Signaling);辣根過氧化物酶-山羊抗兔IgG(KPL);化學發光法顯色試劑盒(Santa Cruz);鏈霉卵白素免疫組化染色試劑盒(北京中杉金橋生物技術有限公司)。
2 動物模型的建立及分組
參照文獻[3]建立大鼠慢性嗎啡依賴及急性催促戒斷模型:以劑量遞增法連續皮下注射嗎啡(10、20、30、40、50 mg·kg-1)5 d,每天2次,間隔12 h(8:00、20:00)。第6天 8:00 皮下注射嗎啡 50 mg·kg-1,2 h 后腹腔注射鹽酸納絡酮(5 mg·kg-1)進行催促戒斷,用改良 Gellert-Holtzman 法[4]評價模型建立是否成功,剔除未成模動物。大鼠隨機分組如下:(1)鹽水對照組(saline-saline, Sal-Sal):背部皮下注射同等劑量的生理鹽水,12只;(2)嗎啡依賴組(saline-morphine, Sal-Mor):按照上述方案進行嗎啡皮下注射,12只;(3)戒斷組(morphine-naloxone, Mor-Nal): 按照上述方案給予嗎啡皮下注射后 2 h進行納絡酮催促戒斷,12只;(4)慢性藥物干預組(CCK-8/L-364718/LY-288513-Mor-Nal):給予嗎啡前30 min分別腹腔注射 CCK-8(50 μg·kg-1,n=10)、L-364718(1 mg·kg-1,n=8)、LY-288513(1 mg·kg-1,n=10),余同(3);(5)溶劑對照組(vehicle-Mor-Nal):給予嗎啡前 30 min,注射同等體積的溶劑(DMSO : 1,3-丙二醇=1∶4,1 mL·kg-1),余同(3),8只。
3 主要方法
3.1Western blotting 實驗結束后應用腦立體定位模具,參照大鼠腦立體定位圖譜,急性分離大鼠腦額葉皮質和海馬。腦組織按每100 mg組織加入1 mL的組織蛋白裂解液( 50 mmol/L Tris-HCl,pH 7.4,150 mmol/L NaCl,1 mmol/L EGTA,1 mmol/L EDTA,1% NP40,1 mmol/L DTT,1 mmol/L PMSF,1 mg/L aprotinin,1 mg/L pepstatin A,1 mg/L leupeptin) ,用電動勻漿器勻漿30 s,冰浴裂解30 min。4 ℃、12 000 r/min離心20 min 后取上清,用Tiangen公司考馬斯亮藍蛋白定量試劑盒(微板法)測定蛋白濃度,分裝后-70 ℃凍存備用。采用SDS-PAGE分離蛋白,用水浴式電轉移裝置將凝膠上的蛋白質轉移到硝酸纖維素膜上,5% 脫脂奶粉封閉1 h,Ⅰ抗4℃孵育過夜(1∶1 000稀釋),TTBS洗膜后,加入HRP標記的羊抗兔IgG Ⅱ抗(1∶10 000稀釋),37 ℃孵育1 h,化學發光法顯色1 min,暗盒中曝光1 min,顯影后將X光片用數碼相機照相后,用ImageJ圖像分析系統對圖像進行吸光度掃描,以各組目的蛋白條帶吸光度值與內參照蛋白吸光度值計算相對比值,以對照組的比值作為1,其余各組與之相比,再次計算比值作為統計值。
3.2免疫組織化學 實驗結束后,用10%水合氯醛麻醉,以0.9%生理鹽水行心腔灌注沖洗后,用4%多聚甲醛固定液灌注固定,迅速斷頭開顱取出全腦,常規制備組織切片、三步法免疫組織化學染色(Ⅰ抗CREB 1∶400、pCREB 1∶50 稀釋)。采用Media Cybernetics公司IPP圖像分析軟件測定陽性神經元平均吸光度值,對CREB和pCREB蛋白表達水平進行半定量分析。
4 統計學處理
數據以均數±標準差(mean±SD)表示,各組均數的比較行單因素方差分析(ANOVA),用最小顯著差異(LSD)法進行兩兩比較,以P<0.05為差異有統計學意義。
結 果
1 嗎啡戒斷模型的建立
實驗中觀察到齒顫、濕狗樣抖動、腹瀉、流淚、流涎、跳躍、體重明顯下降等戒斷癥狀,戒斷組Gellert-Holtzman評分與依賴組相比明顯升高,見圖1。

Figure 1. Effects of CCK-8, L-364718 and LY-288513 on the Gellert-Holtzman score of the overall withdrawal seve-rity.Mean±SD. ▲▲P<0.01 vs Mor-Sal group; *P<0.05,**P<0.01 vs Mor-Nal group; #P<0.05 vs L-364718-Mor-Nal group.
2 CCK-8及其受體拮抗劑對嗎啡戒斷大鼠額葉皮質及海馬CREB和pCREB蛋白表達的影響
Western blotting結果顯示,額葉皮質和海馬CREB蛋白在慢性嗎啡作用以及戒斷后均無明顯變化(P>0.05),見圖2A、C;但是pCREB蛋白表達在嗎啡依賴后明顯增加(P<0.01),在納洛酮急性催促戒斷后進一步升高(P<0.01),見圖2B、D。與戒斷組(Mor-Nal)相比,CCK-8、L-364718和LY-288513慢性干預對大鼠額葉皮質CREB蛋白表達無明顯影響,pCREB蛋白表達明顯降低,見圖2A、B;海馬CREB與pCREB蛋白在L-364718和LY-288513慢性干預后均明顯降低,但CCK-8慢性干預對CREB蛋白表達無明顯影響,僅pCREB蛋白表達明顯降低,見圖2C、D。
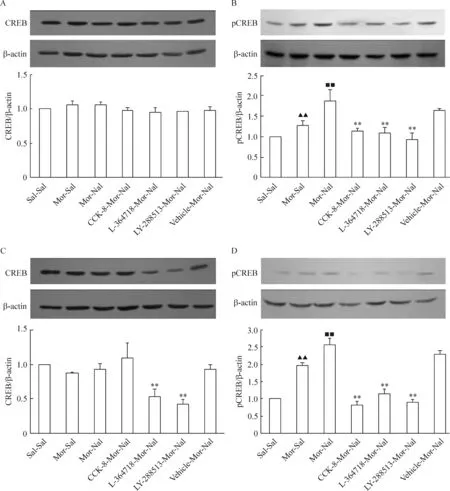
Figure 2. Effects of CCK-8 and CCK receptor antagonist chronic treatments on the expression of CREB (A,C) and pCREB (B,D) in frontal cortex (A,B) and hippocampus (C,D) of morphine withdrawal rats detected by Western blotting.Mean±SD.n=6 in Sal-Sal, Mor-Sal and Mor-Nal groups; n=5 in CCK-8-Mor-Nal and LY-288513-Mor-Nal groups; n=4 in L-364718-Mor-Nal and Vehicle-Mor-Nal groups. ▲▲P<0.01 vs Sal-Sal group; ■■P<0.01 vs Mor-Sal group; **P<0.01 vs Mor-Nal group.
我們選取了額葉皮質第二運動(M2)區和海馬CA1區錐體細胞層神經元作為觀察對象,發現正常大鼠額葉皮質M2區神經元胞漿、胞核均表達CREB蛋白,并且慢性嗎啡作用后無明顯變化(P>0.05)急性納洛酮催促戒斷后CREB蛋白以胞核表達為主,但其表達量無明顯變化(P>0.05),見圖3;pCREB蛋白則僅在胞核中高表達,慢性嗎啡作用后其表達量增加(P<0.01),急性納洛酮催促戒斷后進一步升高(P<0.01),見圖4。海馬CA1區錐體細胞層神經元中,CREB蛋白在胞漿中高表達,胞核低表達,見圖5;pCREB蛋白則在胞核中高表達,見圖6。CCK-8、L-364718和LY-288513慢性干預對戒斷大鼠額葉皮質M2區和海馬CA1區錐體細胞層神經元CREB與pCREB蛋白表達的影響與Western blotting的結果一致。
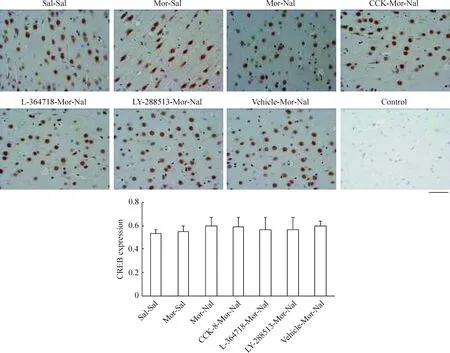
Figure 3. Effects of CCK-8 and CCK receptor antagonist chronic treatments on the expression of CREB in frontal cortex of morphine withdrawal rats detected by immunohistochemistry.Bar=50 μm. Mean±SD.n=6 in Sal-Sal, Mor-Sal and Mor-Nal groups; n=5 in CCK-8-Mor-Nal and LY-288513-Mor-Nal groups; n=4 in L-364718-Mor-Nal and Vehicle-Mor-Nal groups.
討 論
20世紀80年代,有學者提出腦內可能存在阿片肽的對立面,即抗阿片肽,以維持慢性阿片作用下機體的穩態。膽囊收縮素(cholecystokinin,CCK)是一個具有廣泛生物學活性的腦腸肽,它既是胃腸道主要調節激素之一,也作用于中樞神經系統而發揮廣泛的生物學作用。CCK-8是迄今體內最強的抗阿片肽類物質,其通過與靶細胞表面的CCK受體結合后發揮多種生物學作用。實驗研究表明,CCK受體拮抗劑不但能針對戒斷癥狀進行對癥治療,而且有一定程度的防止復吸作用。Blandine等[3]報道CCK2受體的缺失可以導致內源性阿片肽的釋放發生變化。也有研究發現CCK2受體拮抗劑可以有效地減輕慢性嗎啡作用后納絡酮催促戒斷癥狀的發生[4],明顯抑制嗎啡誘導的條件性位置偏愛的形成[5]。然而關于CCK-8減輕嗎啡戒斷癥狀的機制尚不完全明確。
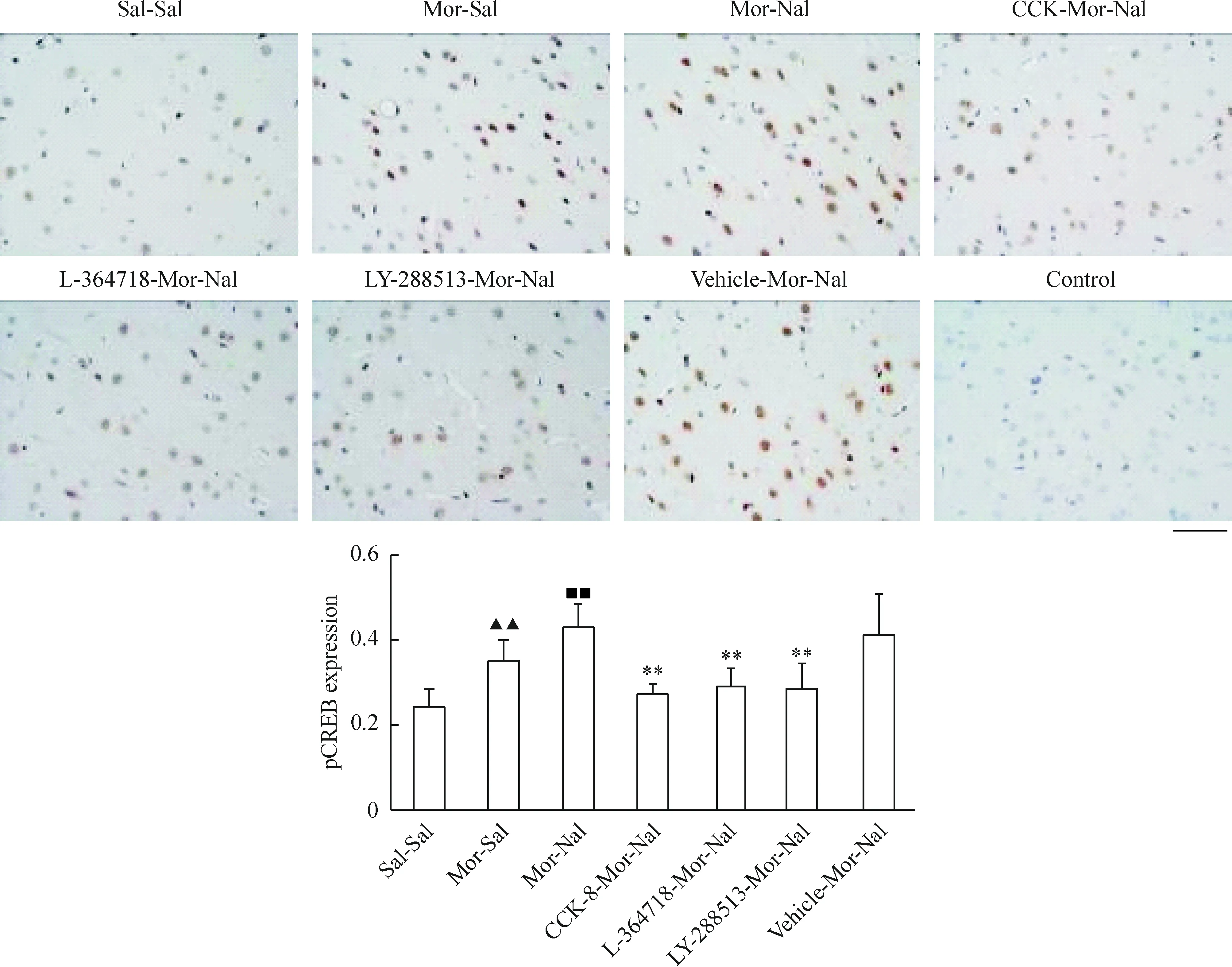
Figure 4. Effects of CCK-8 and CCK receptor antagonist chronic treatments on the expression of pCREB in frontal cortex of morphine withdrawal rats detected by immunohistochemistry.Bar=50 μm.Mean±SD.n=6 in Sal-Sal, Mor-Sal and Mor-Nal groups; n=5 in CCK-8-Mor-Nal and LY-288513-Mor-Nal groups; n=4 in L-364718-Mor-Nal and Vehicle-Mor-Nal groups. ▲▲P<0.01 vs Sal-Sal group; ■■P<0.01 vs Mor-Sal group; **P<0.01 vs Mor-Nal group.
阿片類物質依賴的神經分子生物學機制尚未闡明,其過程涉及到受體、受體后的信號轉導,基因的轉錄、翻譯和調控,效應子的表達及生物效應的產生等多個方面。已知急性嗎啡可以降低細胞內cAMP水平。然而,隨著嗎啡持續處理,cAMP水平逐漸恢復到正常。再用阿片受體拮抗劑處理,cAMP水平就增加到遠遠超過正常值。cAMP的上調可促進PKA活性的增加,進而促進CREB的轉錄因子特異性絲氨酸殘端磷酸化。CREB屬于堿性氨基酸亮氨酸拉鏈轉錄因子家族,它具有轉錄活性的二聚體和無轉錄活性的單體。Ser133是CREB激活的關鍵位點,磷酸化后的CREB 形成同源二聚體或與CREB/ATF 家族的其它成員形成異二聚體,使CREB活化[6]。目前普遍認為,磷酸化激活的CREB形成二聚體與cAMP反應元件(cAMP response element, CRE)結合,引起一組靶基因表達上調,導致多種蛋白產物持續增多,參與了應激、認知等功能的調節[7-8]。另外,CREB誘導的蛋白表達還可引起神經元可塑性和適應性改變,這種適應性的改變是慢性嗎啡依賴和耐受的共同環節,也是形成慢性嗎啡依賴和戒斷的重要機制之一[9-10]。CREB第133位絲氨酸磷酸化是其調控基因表達的基本條件,CREB 磷酸化的水平與細胞膜的去極化能力有關, 介導細胞膜的去極化過程, 誘導c-fos基因的表達。磷酸化CREB對下游靶基因轉錄過程的調控作用可能是嗎啡依賴長時程適應性改變的基礎之一,CREB在藥物依賴的形成及表達過程中起著承上啟下的作用。
嗎啡成癮的形成是由多個腦區共同參與的結果。額葉皮質參與了中腦-皮層多巴胺能系統,涉及高級認知功能與動機功能。海馬作為邊緣結構則廣泛地參與了學習與記憶過程,尤其是長期記憶過程的形成,并且在嗎啡成癮記憶形成過程中起重要作用。慢性阿片處理能減弱海馬CA1區突觸的長時程增強,改變海馬的突觸可塑性[11]。研究表明額葉皮質、海馬等腦區不僅在成癮記憶方面發揮著重要作用,同時也參與軀體戒斷癥狀的形成[12-13]。
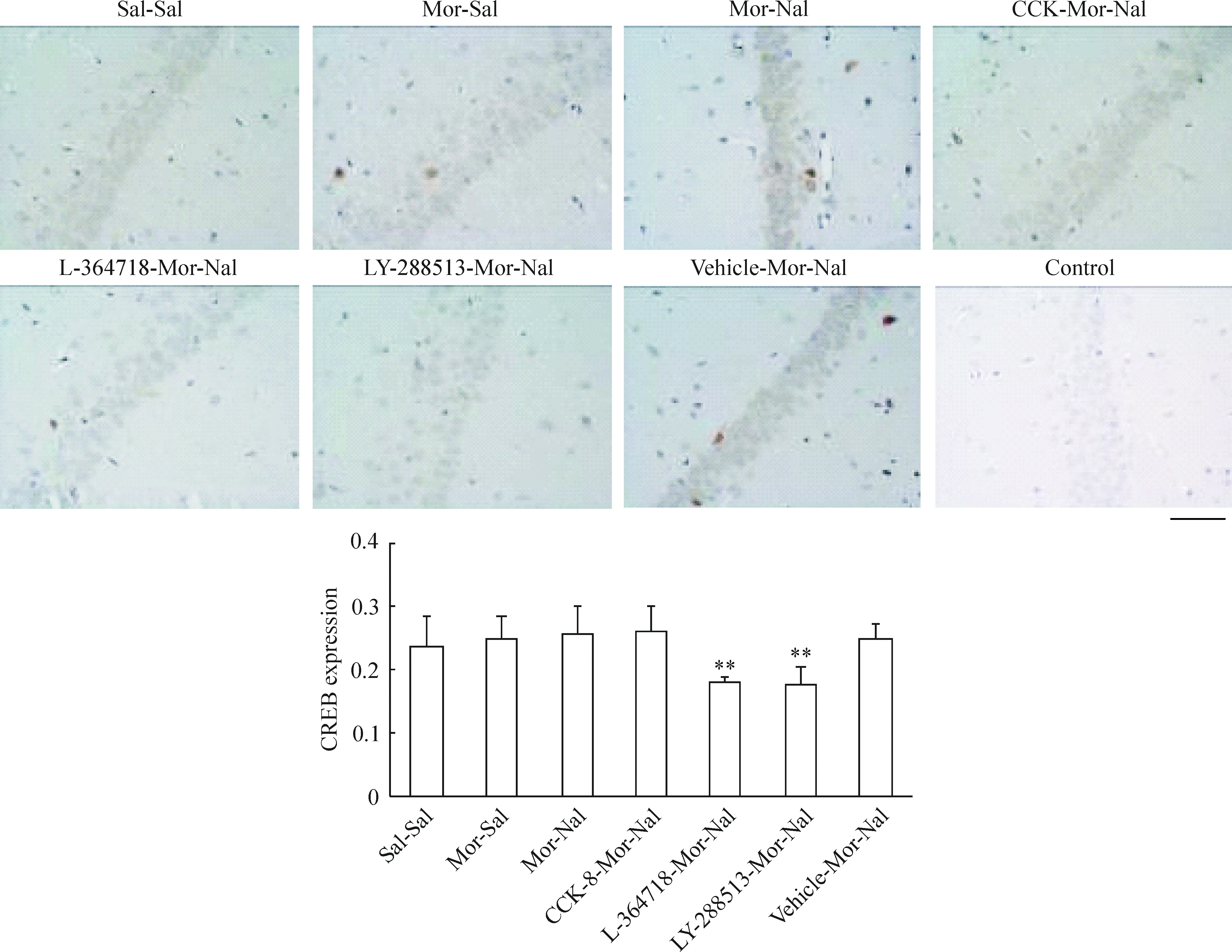
Figure 5. Effects of CCK-8 and CCK receptor antagonist chronic treatments on the expression of CREB in hippocampus of morphine withdrawal rats detected by immunohistochemistry.Bar=50 μm.Mean±SD.n=6 in Sal-Sal, Mor-Sal and Mor-Nal groups; n=5 in CCK-8-Mor-Nal and LY-288513-Mor-Nal groups; n=4 in L-364718-Mor-Nal and Vehicle-Mor-Nal groups.**P<0.01 vs Mor-Nal group.
在Western blotting實驗結果中,我們發現慢性嗎啡作用后,大鼠額葉皮質和海馬CREB蛋白表達無明顯變化,但pCREB蛋白表達明顯增加。納洛酮急性催促戒斷后,CREB蛋白表達仍未發生明顯變化,pCREB蛋白表達進一步升高。與戒斷組相比,CCK-8、L-364718和LY-288513慢性干預對大鼠額葉皮質CREB蛋白表達無明顯影響,pCREB蛋白表達明顯降低;L-364718和LY-288513急性干預對大鼠額葉皮質CREB/pCREB蛋白表達均無明顯影響。海馬CREB和pCREB蛋白表達在L-364718和LY-288513慢性干預后均明顯降低,但CCK-8慢性干預及L-364718、LY-288513急性干預對CREB蛋白表達無明顯影響,僅pCREB蛋白表達明顯降低。CCK-8急性干預對戒斷大鼠額葉皮質、海馬CREB與pCREB蛋白表達均無明顯影響。免疫組織化學結果發現正常大鼠額葉皮質M2區神經元胞漿、胞核均表達CREB蛋白,并且慢性嗎啡作用后無明顯變化;急性納洛酮催促戒斷后CREB蛋白以胞核表達為主,但其表達量無明顯變化。pCREB蛋白則在胞核中高表達,慢性嗎啡作用后其表達量增加,急性納洛酮催促戒斷后進一步升高。海馬CA1區錐體細胞層神經元中,CREB蛋白在胞漿中高表達,而胞核低表達;pCREB蛋白則在胞核中高表達。CCK-8、L-364718和LY-288513慢性干預對戒斷大鼠額葉皮質M2區和海馬CA1區錐體細胞層神經元CREB與pCREB蛋白表達的影響與Western blotting實驗中的結果一致。
分析所得到的實驗結果,我們考慮給予CCK受體拮抗劑后,拮抗了內源性CCK的作用,使CCK的生理效應喪失,減輕了嗎啡戒斷癥狀,并使受體后途徑的相關指標發生了相應變化。CCK受體包括CCK1和CCK2受體,分別與Gq和Gq/Gs偶聯。CCK1受體通過IP3-Ca2+-CaMK-CREB以及DAG-PKC-CREB信號通路,CCK2受體通過cAMP-PKA-CREB以及DAG-PKC-CREB信號通路調節下游基因表達,參與體內多種生理功能的調節。這提示CCK1和CCK2受體拮抗劑均可通過各自的信號轉導通路降低嗎啡戒斷大鼠額葉皮質和海馬內CREB磷酸化的水平,進而減輕嗎啡戒斷癥狀。而CCK-8慢性干預卻減輕了納洛酮引起的急性催促戒斷癥狀,與CCK受體拮抗劑的作用一致。本文認為,這與CCK-8的效應濃度有密切關系[14-16],并推測在注射嗎啡前外源性給予高濃度CCK-8慢性干預時,發揮的是藥理效應,其可能通過抑制外源性阿片物質和阿片受體的結合,或者影響外源性阿片物質所產生的后效應而發揮作用;另一方面,高濃度的CCK還有可能通過直接激活阿片受體發揮作用[17],其具體機制還需進一步探討。

Figure 6. Effects of CCK-8 and CCK receptor antagonist chronic treatments on the expression of pCREB in hippocampus of morphine withdrawal rats detected by immunohistochemistry. Bar=50 μm. Mean±SD.n=6 in Sal-Sal, Mor-Sal and Mor-Nal groups; n=5 in CCK-8-Mor-Nal and LY-288513-Mor-Nal groups; n=4 in L-364718-Mor-Nal and Vehicle-Mor-Nal groups..▲▲P<0.01 vs Sal-Sal group; ■■P<0.01 vs Mor-Sal group; **P<0.01 vs Mor-Nal group.
總之,CCK在嗎啡成癮及戒斷過程中發揮了重要作用,并通過調節受體后相關轉錄調控信號蛋白CREB的表達及其磷酸化過程而發揮此作用,為戒毒提供了一種新的思路。
[參 考 文 獻]
[1] Faris PL, Komisaruk R, Watkins LR, et al. Evidence for the neuropeptide cholecystokinin as an antagonist of opiate analgesia[J]. Science, 1983, 219(4582): 310-312.
[2] Hebb AL,Poulin JF, Roach SP, et al. Cholecystokinin and endogenous opioid peptides: interactive influence on pain, cognition, and emotion[J]. Prog Neuropsychopharmacol Biol Psychiatry, 2005, 29(8):1225-1238.
[3] Blandine P,Franc B,Axelle S,et al.Deletion of CCK2 receptor in mice results in an upregulation of the endogenous opioid system[J]. J Neurosci, 2002, 22(5):2005-2011.
[4] Noble F, Wank A, Crawley JN, et al. International Union of Pharmacology. XXI. Structure, distribution and functions of cholecystokinin receptors[J]. Pharmacol Rev, 1999, 51(4):745-781.
[5] Mitchell JM, Bergren LJ, Chen KS, et al. Cholecystokinin is necessary for the expression of morphine conditioned place preference[J]. Pharmacol Biochem Behav, 2006, 85(4):787-795.
[6] Chao J, Nestler EJ. Molecular neurobiology of drug addiction[J]. Annu Rev Med, 2004, 55:113-132.
[7] 于 琦, 金光亮. 三種復方對慢性應激模型大鼠海馬CREB、BDNF基因表達的影響[J]. 中國病理生理雜志, 2009, 25(3):591-594.
[8] 蔡曉紅, 張存雪, 周永海, 等. 慢性間歇低氧對幼鼠認知及相關腦區CREB的影響[J]. 中國病理生理雜志, 2010, 26(5):895-900.
[9] Goldstein A,Tachibana S,Lowney LI,et al. Dynorphin-(1-13),an extraordinary potent opioid peptide[J]. Proc Natl Acad Sci U S A, 1979, 76(12):6666-6670.
[10] 馬春玲, 楊勝昌, 文 迪, 等. CCK-8對嗎啡戒斷大鼠額葉皮質、海馬CREB和pCREB的影響[J]. 中國病理生理雜志, 2010, 26(10):2052.
[11] Pu L, Bao GB, Xu NJ, et al. Hippocampal long term potentiation is reduced by chronic opiate treatment and can be restored by re-exposure to opiates [J]. J Neurosci, 2002, 22(5):1914-1921.
[12] Ammon S, Mayer P, Riechert U, et al. Microarray analysis of genes expressed in the frontal cortex of rats chronically treated with morphine and after naloxone precipitated withdrawal[J]. Brain Res Mol Brain Res, 2003, 112(1-2):113-125.
[13] Fan GH, Wang LE, Qiu HC, et al. Inhibition of calcium/calmodulin-dependent protein kinase-II in rat hippocampus attenuates morphine tolerance and dependence[J]. Mol Pharmacol, 1999, 56(1):39-45.
[14] Doi T, Jurna I. Analgesic effect of intrathecal morphine demonstrated in ascending nociceptive activity in the rat spinal cord and ineffectiveness of caerulein and cholecystokinin octapeptide[J]. Brain Res, 1982, 234(2):399-407.
[15] Barbaz BS, Hall NR, Liebman JM.Antagonism of morphine analgesia by CCK-8-S does not extend to all assays nor all opiate analgesics[J]. Peptides, 1988, 9(6):1295-1300.
[16] Rezayat M, Azizi N, Zarrindast MR. On the mechanism(s) of cholecystokinin (CCK): receptor stimulation attenuates morphine dependence in mice[J]. Pharmacol Toxicol, 1997, 81(3):124-129.
[17] Romanelli L, Amico MC, Mattioli F, et al. Interactions between cholecystokinin and opioids in the isolated guinea-pig ileum[J]. Br J Pharmacol, 1999, 127(4): 909-918.
猜你喜歡
作文周刊·小學二年級版(2022年20期)2022-05-05 01:33:06
娃娃樂園·綜合智能(2020年8期)2020-08-28 00:32:14
創新作文(小學版)(2019年10期)2019-09-25 08:12:28
作文周刊·小學二年級版(2018年9期)2018-04-18 10:01:40
小學生導刊(2018年1期)2018-03-15 08:02:37
小學生學習指導(低年級)(2017年5期)2017-05-04 04:14:38
科技知識動漫(2016年6期)2016-06-24 21:04:53
大灰狼(2015年6期)2015-07-16 21:01:00
作文與考試·小學高年級版(2015年17期)2015-05-30 10:48:04
汽車觀察(2009年1期)2009-02-18 09:11:50
