甘肅省野生草地早熟禾多胚苗發生研究
2014-09-11 06:30:26田晨霞馬暉玲武新赫
草原與草坪 2014年6期
關鍵詞:融合
田晨霞,馬暉玲,張 靜,武新赫
(甘肅農業大學 草業學院/草業生態系統教育部重點實驗室/甘肅省草業工程實驗室/中-美草地畜牧業可持續發展研究中心,甘肅 蘭州 730070)
草地早熟禾為禾本科草本植物,原產歐洲和亞洲北部,在我國華北、西北、東北地區以及西南高海拔地區生長良好[1,2]。我國北方的草坪草種中,由于草地早熟禾的坪用特性好,廣泛用于各種類型的草坪建植中[3,4]。如今我國草坪建植的面積在逐步增大,而建植草坪所需種子幾乎全靠進口。從長遠看,培育我國的草地早熟禾系列品種已勢在必行。
草地早熟禾的生殖方式為兼性無融合生殖,其中以體細胞無孢子生殖占絕對優勢,還有孤雌生殖和極少量的二倍體孢子生殖[5]。無融合生殖是指不經過雌雄配子融合而產生種子的一種特殊生殖方式,能使基因型的雜合性得以保持,從而可以固定雜種優勢,對作物育種具有極其重要的意義。無融合生殖具有雙胚珠、雙胚囊、雙胚雙苗或多苗等特點[6,7],黃群策等[8]報道,被子植物多胚現象的發現可以追溯到18世紀著名的植物學家Leeuwenhok對柑桔屬(Citrus)植物種子形態學的觀察工作。隨后,越來越多的研究者不斷地在其他植物中觀察到多胚現象和多胚苗現象。被子植物的多胚苗來源于種子中相應多胚的發育,而被子植物的多胚包括真多胚、假多胚和不定胚3種類型。在被子植物的1個胚珠中包含2個或2個以上胚的現象稱之為多胚現象。由一粒多胚種子可以同時萌發出兩株或兩株以上的幼苗,這種一籽多首稱之為多胚苗。近半個世紀以來,全球主要農作物增產潛力在生產上的明顯發揮主要受益于充分地利用了農作物的雜種優勢。在充分證實植物雜種優勢具有普遍性的同時越來越多的研究者熱心于通過無融合生殖途徑來固定作物雜種優勢的探索性研究。母錫金等[9]對草地早熟禾的多胚現象進行了研究,得出早熟禾的穎果產生雙胚苗和三胚苗是常見的。并且過去大量的研究結果表明,在一些被子植物中多胚現象與其無融合生殖現象具有一定的正相關性[10-12]。
無融合生殖是植物生殖生物學中的一個重要方面,對其的深入研究將有助于闡明植物生殖生物學中的重大理論問題,對培育新品種也具有重要的現實意義[13]。因此,對草地早熟禾無融合生殖的方式和過程的理解和研究,對其野生種或生態型的搜集并對其多胚現象進行研究,對不同品種草地早熟禾多胚率進行統計,具有重要的科學意義和應用價值。
本次研究主要以收集的甘肅定西、隴西、清水、秦州、隴南5個地區的野生草地早熟禾種質材料和商用草地早熟禾品種“巴潤”為材料,通過種子發芽試驗,比較不同來源種質材料草地早熟禾發芽率及多胚苗發生率的不同,為進一步研究其無融合生殖發育機理,選育我國本土的草地早熟禾品種提供基礎數據。
1 材料和方法
1.1 試驗材料
供試材料為2012年8~9月在甘肅省定西,秦州,清水,隴西,定西,隴南5個地區采集的野生草地早熟禾和商用品種“巴潤”。 種子經人工清選、自然干燥后,放入紙袋室內常溫(10~25 ℃)保存。
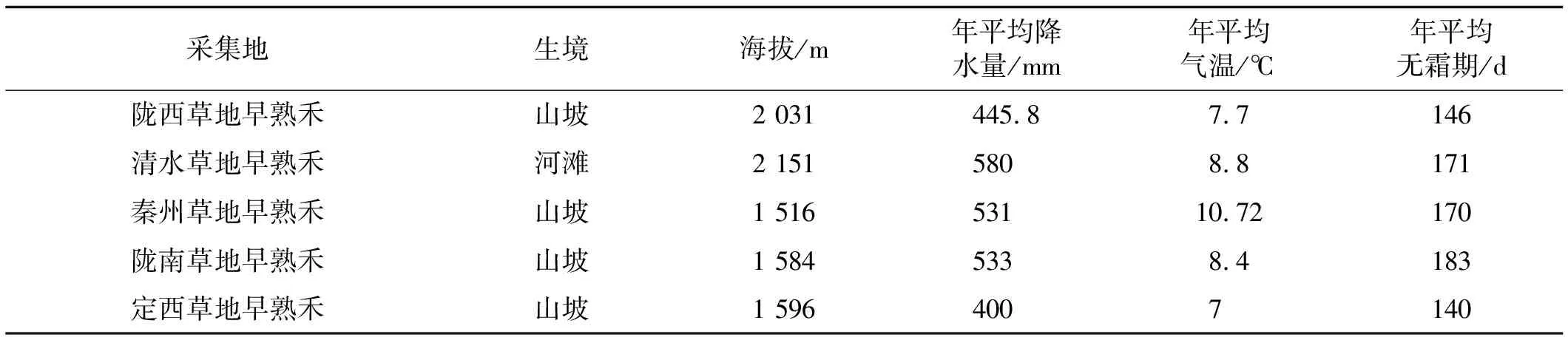
表1 野生草地早熟禾采集地生境信息
1.2 試驗方法
1.2.1 發芽試驗 種子發芽試驗采用國家規程鑒定標準(國家標準是種子幼根伸出長度達到與種子長度等長為正常發芽)。供試材料每品種各取種子2 000粒,每培養皿100粒種子,每品種重復20次。15/25 ℃變溫條件下紙上萌發法測定種子的萌發率及發芽指數,8 h光照,16 h黑暗。按規定日期計算發芽率(第20 d),并在種子出芽后統計出苗率及單苗、雙苗和多苗所占比率,并采用SPSS分析統計結果。
1.2.2 數據統計及計算方法
發芽率(%)=(n/N)×100%
雙胚苗率(%)=(n1/n)×100%
三胚苗率(%)=(n2/n)×100%
多苗率(%)=(n1+n2)/n×100%
式中:n1為雙胚苗數,n2為三胚苗數,n為種子正常發芽粒數,N為供試種子總數。
2 結果與分析
2.1 不同地域草地早熟禾品種發芽率的統計
各供試草地早熟禾材料的發芽率為48%~68%,其中,以秦州野生草地早熟禾發芽率最高,其數值為68%;清水、隴南和“巴潤”次之,分別為61%,60%和60%;定西和隴西發芽率低于以上4個品種,分別為50%和48%(表2)。
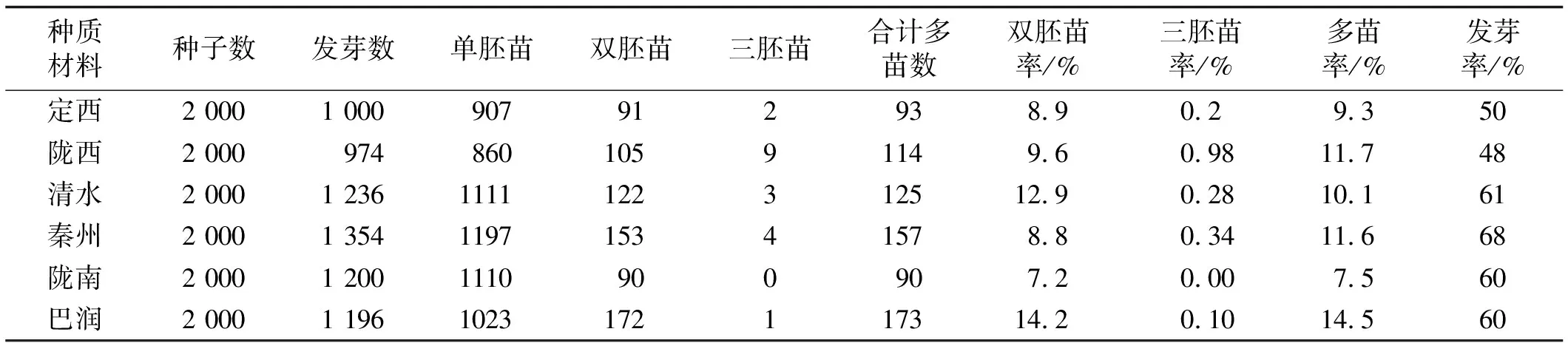
表2 各種質材料草地早熟禾多苗發生率的統計
2.2 不同地域野生草地早熟禾雙胚苗發生率
各供試品種草地早熟禾雙胚苗7.2%~14.2%,其中,商用品種巴潤雙胚苗率高于野生草地早熟禾,其數值為14.2%;清水、隴西、定西和秦州次之,分別為12.9%、9.6%、8.9%和8.8%;隴南的雙胚苗率最低,為7.2%(表2)。
2.3 草地早熟禾種質材料三胚苗發生頻率的統計
各供試種質材料草地早熟禾的三胚苗發生率為0%~0.98%,其中隴西野生草地早熟禾三胚苗率高于其他種質材料草地早熟禾,其數值為0.98%;秦州、清水、巴潤和隴南次之,分別為0.34%、0.28%、0.10%和0;隴南野生草地早熟禾中未發現三胚苗(表2)。
2.4 草地早熟禾種質材料多苗發生率的統計
各供試種質材料草地早熟禾的多苗發生頻率為7.5%~14.5%,其中以商用品種巴潤多苗率最高,其數值為14.5%;隴西、秦州、清水和定西次之,分別為11.7%、11.6%、10.1%和9.3%;隴南的多苗數最低,為7.5%(圖1)。
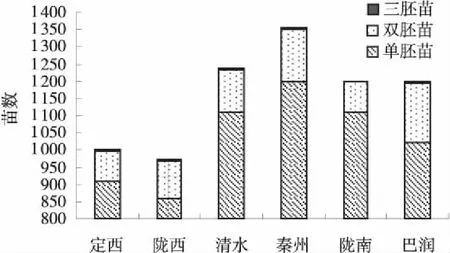
圖1 不同野生種質材料草地早熟禾多胚苗數及發生率Fig.1 Chart of polyembryony seeding rate in wild Poa pratensis
2.5 草地早熟禾多胚苗類型的劃分
草地早熟禾種子出芽20 d后,對多胚苗進行拍照。不同種質材料的草地早熟禾中均出現較多的多胚苗現象,以雙胚苗居多。
從草地早熟禾的幼苗的外部形態進行分類,可將草地早熟禾的苗型分為正常單苗、雙苗和三苗;其中雙苗又分為簇生雙苗、對生雙苗、間生雙苗3種情況,在雙苗中簇生雙苗發生頻率最高;其中三苗種子又包括對生三苗、簇生雙苗,三苗發生頻率極低,還不到百分之一。按照草地早熟禾種子萌發多苗的生長狀況,又可將多苗詳細劃分為以下幾種類型:
單苗型:種子萌發出1個幼小苗(圖2-A)。
雙苗型:種子萌發出2個幼小苗。
簇生雙苗型:種子同一個萌發孔萌發出2個幼小苗(圖2-B-a)。
簇生雙苗同步型:種子同一個萌發孔萌發出2個大小相當的幼小苗(圖2-B-a1)。
簇生雙苗異步型:種子同一個萌發孔萌發出2個大小不等的幼小苗(圖2-B-a2)。
對生雙苗型:種子兩個相對的萌發孔各萌發出1個幼小苗。
對生雙苗同步型:種子兩個相對的萌發孔各萌發出1個幼小苗,兩個幼苗大小相當(圖2-B-b1)。
對生雙苗異步型:種子兩個相對的萌發孔各萌發出1個幼小苗,兩個幼苗大小存在較大差異(圖2-B-b2)。
間生雙苗異步型:種子一端的的萌發孔和種子的中間部位各生出1個幼苗,這兩個幼苗一般一大一小(圖2-B-c)。
三苗型:種子萌發出3個幼小苗。
簇生三苗型:種子同一個萌發孔萌發出 3個幼小苗。
簇生三苗同步型:種子同一個萌發孔萌發出 3個大小相當的幼小苗(圖2-C-a1)。
簇生三苗異步型:種子同一個萌發孔萌發出 3個幼小苗,其中1大2小或1小2大(圖2-C-a2)。
對生三苗異步型:種子的兩個相對的萌發孔一端生出1個幼小苗,另一端生出2個幼小苗。
對生三苗異步型:種子的萌發孔一端生出 1個較大的幼苗,另一端生出2個較小幼苗生長同步(圖2-C-b1)。
對生三苗異步型:種子一端的萌發孔萌發出 2大小不等的幼苗,另1端生長出1個幼苗(圖2-C-b2)。
3 討論與結論
3.1 不同草地早熟禾材料的發芽率及多胚苗率
試驗結果表明,不同草地早熟禾材料的發芽率、雙胚苗率、三胚苗率及多苗率之間存在差異。不同草地早熟禾發芽率在48%~68%,單胚苗率在86%~92%,雙胚苗率在7.5%~14.5%,三胚苗率在0~0.98%。母錫金等[9]對草地早熟禾的多胚現象的研究表明,單胚苗, 雙胚苗或三胚苗的比率為93.17%、5.57%和1.24%。趙世緒[14]早期在研究早熟禾屬作物多胚現象時報道,草地早熟禾屬產生多胚的頻率為8%~12%。但很少有文獻記載野生草地早熟禾多胚苗率,且鮮有文獻對不同品種草地早熟禾的多胚苗率進行統計。
3.2 草地早熟禾多胚苗類型

圖2 草地早熟禾多胚苗Fig.2 Poly-embryo seedling of P.pratensis
草地早熟禾的多胚苗因其多胚的來源不同,出苗方式也存在較大差異。劉向東等[15]將水稻的多胚苗途徑歸納為4類10種。王巨媛等[16]曾對韭菜多胚苗發生頻率及類型進行劃分,與草地早熟禾相似,從外部形態特征上可將發芽的草地早熟禾種子分為3種類型,即正常單芽、雙芽和三芽種子;其中雙芽種子又包括對生雙芽、簇生雙芽、種子中部和萌發孔各萌發一個芽3種情況,在雙芽中以簇生雙芽種子的發生頻率最高;其中三芽種子又包括對生三芽、簇生三芽,三芽發生頻率極低,還不到百分之一。
3.3 多胚現象與無融合生殖的關系
大量的研究結果表明,在被子植物中多胚現象與其無融合生殖現象具有一定的正相關性,因而許多學者以此為依據試圖通過在被子植物的種質資源中尋找具多胚現象的植物材料,進而篩選出一些可能具有無融合生殖特性的基因資源[8]。但是只有特定起源的多胚苗材料在其雜種優勢固定中才具有較大的實用價值,因而在評價多胚苗材料時,必須要弄清其胚胎學特性[17]。Grazi等[18]指出草地早熟禾生殖方式為兼性無融合生殖,其無融合率因品種的不同而異,其值在23%~90%,有性品種中僅有2%的后代植株由合子胚發育而來。由此證明無融合生殖和多胚現象之間必然存在一定的聯系。試驗對野生草地早熟禾的多胚苗率及多胚苗類型進行了系統分析,試圖為闡明草地早熟禾多胚的發生機制提供依據,從而估價其應用價值。
3.4 草地早熟禾多胚現象發生的影響因素
多胚苗的發生涉及多胚的形成和多胚苗的表現兩大階段,多胚的形成不僅受到遺傳背景的影響,多胚苗的表現又受到外界條件的影響。歸納其發生的原因,主要為:
3.4.1 基因效應 草地早熟禾中多胚現象發生的原因可能是某種遺傳性狀在特定環境下基因表達所致。草草地早熟禾的無融合生殖方式主要以無孢子生殖為主[19],還有少量的孤雌生殖和二倍體孢子生殖[20,21]。所以在胚形成過程中一籽多苗發生頻率較高,這些生殖性狀可能是由遺傳基因所控制。
3.4.2 環境效應 多胚苗性狀的發生和發生頻率的高低在一定程度上受某些環境因素的影響。Lakshmanan等[22,23]發現在柑桔屬植物中,植株的年齡、年產果量、植株的營養狀態和枝條的著生部位等因素對其不定胚的發生類型及數目有明顯影響。本試驗研究的甘肅野生草地早熟禾和商用草地早熟禾“巴潤”,因其生長環境的差異,其多胚苗率也存在顯著差異。
3.4.3 其他效應 試驗觀察到草地早熟禾的各個材料多胚率在9.3% ~14.5%,但是多胚在種子上生長的位置各不一樣,其中,在正常位且發育正常的胚,也有多胚中的額外胚,尤其是那些遠離正常胚位的是否是由胚囊外體細胞發育即無融合生殖所形成的不定胚,還有待進一步研究。
參考文獻:
[1] 殷朝珍,王兆龍,葛才林.草地早熟禾無融合生殖及其育種利用研究進[J].草原與草坪,2006(1):18-23.
[2] 文金花,馬玉壽,施建軍,等.利用草地早熟禾改建江河源區“黑土型"退化草地的研究[J].草原與草坪,2006(2):41-44.
[3] 趙桂琴,曹致中.草地早熟禾無融合生殖的細胞學鑒定[J].草業學報,1997,6(4):64-70.
[4] 邱正強,曹玉紅,梁麗,等.青海草地早熟禾坪用性狀的比較研究[J].草原與草坪,2009(5):50-55.
[5] 田晨霞,馬暉玲,張詠梅.草地早熟禾胚胎發育類型及無融合生殖特征[J].中國農業科學,2013,46(13):2633-2642
[6] Hanna W W,Bashaw E C,Apomixis:Its identification and use in plant breeding[J].Crop Sci,1987,27:1136-1139.
[7] Tang G Y,Schertz K F,Bashaw E C.Apomixis in Sorghumlines and their F1Progenies[J].Botanical Gazette,1980,141(3):294-299.
[8] 黃群策,孫敬三.被子植物多胚苗的研究進展[J].植物學通報,1998,15(2):1-7.
[9] 母錫金,王伏雄,梁鐵兵.草地早熟禾的多胚現象[J].植物學通報,1994(1):68-75.
[10] 周開達.四川無融合生殖水稻(SAR-1)的初步研究[J].中國科學(B輯),1992(8):156-162.
[11] 吳伯驥,謝明唐,陳毅平,等.水稻雄性不育系C1001中發現無融合生殖株系的細胞學和胚胎學研究[J].中國科學,1990(10):1049-1052.
[12] 郭學興,劉剛,曾秀英,等.水稻C1001多胚苗發生規律的研究[J].四川師范學院學報,1991,12(3):227-229.
[13] 賀鳳麗,馬三梅.植物無融合生殖研究新進展[J].生命科學,2009,21(1):139-144.
[14] 趙世緒.作物胚胎學[M].北京:農業出版社,1982:189-214.
[15] 劉向東,陳啟鋒,李維明,等.水稻多胚苗的初步研究Ⅲ.來源與頻率[J].福建農學院學報,1993,22(2):148-154.
[16] 王巨媛,翟勝,馮輝.韭菜多胚苗發生頻率及其類型的劃分[J].種子,2005,24(10):32-35.
[17] 袁良琛.多胚水稻ApⅢ的形態學觀察和胚胎學研究[D].濟南:山東大學,2003.
[18] Grazi F,Umaerus M,Akerberg E.Oberservations on the mode of reproduction and the embryology of Poa pratensis[J].Hereditas,1961,47:489-541.
[19] Savidan Y.Transfer of apomixis through wide crosses[M]∥Savidan Y,Carman J G,Dresselhaus T.Mexico:CIMMYT,IRD,European Commissi on DG VI,2001:153-167.
[20] 王風翱.作物胚胎學基礎[M].長沙:湖南科技出版社,1987.
[21] Marshall D R,Brown A H D.The evolution of apomixis[J].Heredity,1981,47(1):1-15.
[22] Lakshmanan K K,Ambegaokar K B.Embryology in angosperms,1984,445-474.
[23] 許方.園藝植物胚胎學[M].北京:農業出版社,1993.
猜你喜歡
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
數學年刊A輯(中文版)(2022年4期)2022-02-16 08:17:34
今日農業(2021年19期)2022-01-12 06:16:36
中老年保健(2021年11期)2021-08-22 03:15:44
無線電通信技術(2021年4期)2021-07-13 08:58:28
無線電通信技術(2021年3期)2021-06-08 03:33:48
中學生數理化(高中版.高考數學)(2021年1期)2021-03-19 08:28:38
無線電工程(2020年11期)2020-10-29 01:25:46
現代出版(2020年3期)2020-06-20 07:10:34
福利中國(2015年4期)2015-01-03 08:03:38
