葎草幼苗光合生理特性對鉻脅迫的響應
2014-11-08 03:17:42王碧霞胥曉李霄鋒
草業學報 2014年4期
關鍵詞:植物
王碧霞,胥曉,李霄鋒
(1.西華師范大學生命科學學院 西南野生動植物資源保護教育部重點實驗室,四川 南充637009; 2.張家口市農業環境與農產品質量管理站,河北 張家口075000)
鉻在自然界中約占地殼總量的0.02%,常存在于土壤、大氣、水及各類生物體內[1]。然而隨著經濟全球化進程的加快,鉻被廣泛應用于化工、冶金、機械等行業,它在環境中的排放速度顯著增加,逐漸成為土壤污染的5種主要重金屬之一[2],據統計,我國每年新排放鉻渣約60萬t,歷年累積存鉻渣近600萬t,受鉻嚴重污染的土壤已達1250多萬t[3]。一般而言,自然土壤中適量濃度的鉻對種子萌發和植物生長是有利的[4-5],但含量過高卻會導致植物葉綠素分解,光合作用下降[4-6],細胞內活性氧累積,產生氧化脅迫[7],對植物生長具有抑制作用。因此,研究不同鉻濃度下植物的光合生理特性變化,對了解植物對不同程度鉻污染土壤的適應以及修復植物的篩選具有實際意義。目前,有關土壤鉻污染修復植物的研究僅集中在少數幾種超富集植物上,如李氏禾(Leersiahexandra)、Dicomaniccolifera和Suterafodina[8-9],但這類植物通常生長慢、植株小、周期長、經濟效益低,導致其在植物修復工程中應用較少。因此,選用生長快、周期短、具有經濟價值的普通植物用于重金屬鉻污染土壤的修復具有重要意義。
葎草(Humulusscandens)為桑科(Moraceae)葎草屬(Humulus),一年生或多年生草本,在我國青海、新疆、甘肅、西藏等省、市廣泛分布,其嫩莖和葉可用于畜禽飼料和添加劑,還可全草入藥,具有醫療、保健等功效。葎草生性強健、抗逆性強,不僅是工程礦地、裸露地和棄耕地的先鋒物種,而且在控制水土流失、改善生態環境方面也具有明顯優勢。鑒于葎草在中藥材方面的大量運用,近年來多數研究主要集中在其化學成分、藥用成分,遺傳育種等生理生化方面[10-12],同時也偶見葎草在水分、增溫脅迫下具有性別差異的文獻[13-14],但重金屬脅迫對葎草生理生態的影響尚未見報道。故本研究以常見的葎草為試材,研究不同鉻濃度對葎草幼苗葉片的氣體交換、丙二醛(MDA)含量、葉綠素含量和葉綠素熒光參數等生理生態指標的影響,以期揭示鉻脅迫環境下葎草的抗逆性及該類植物對脅迫環境的響應機制,為葎草進一步用于重金屬鉻污染土壤的修復植物研究提供理論依據。
1 材料與方法
1.1 試驗材料與設計
2012年10月中下旬采集南充市本地葎草野生種子,于2013年2月15日播種,待生長1個月后,選擇長勢基本一致的50株幼苗移栽到體積為10 L的塑料盆內,每盆裝入均質土壤7.5 kg,河沙1.5 kg,每盆1株,緩苗20 d。于4月8日進行鉻脅迫實驗,重金屬試劑為Cr3+。參照王愛云等[5]的方法,設Cr3+濃度為0,50,200,300,500 mmol/L共5個處理,將Cr3+按相應濃度配成1000 mL的溶液緩慢均勻澆灌于土壤中,每處理分別10株,對照組(10株)澆施等量清水。為防止溶液滲漏,對照組和處理組塑料盆內均套有塑料袋(土壤置于塑料袋內)。
1.2 測量方法
1.2.1氣體交換參數測定 于2013年4月28日,隨機選取不同處理下的植株各5株,采用LI-6400便攜式光合測定儀(Li-Cor公司,美國)測定氣體交換特征。每個處理隨機選擇5株,測定植株上部第5或6片全展向陽葉片。測定時間為上午8:00-11:30,測定指標為凈光合速率(Pn)、蒸騰速率(Tr)、氣孔導度(Gs)、胞間CO2濃度(Ci)等。瞬時水分利用效率(WUEi)為Pn與Tr之比,氣孔限制值(Ls)[Ls=(Ca-Ci)/Ca]采用Berry和Downton[15]的方法計算。測定時的環境溫度為(28.0±1.0)℃;大氣相對濕度為40%~43%;大氣CO2濃度為(370±10) mmol/mol;光照強度為(1400±50) mmol/(m2·s)。
1.2.2丙二醛(MDA)含量及葉綠體色素含量測定 MDA含量的測定采用硫代巴比妥酸(TBA)比色法[16],葉綠素含量的測定采用丙酮提取法[17]。
1.2.3葉綠素熒光參數測定 采用便攜式調制葉綠素熒光儀(Junior-Pam,Heinz Walz,Effeltrich, Germany),測量前使功能成熟葉片經過充分的暗適應后,測定葎草成熟葉片的光下最大熒光(Fm′)、 光下最小熒光(Fo′)、初始熒光(Fo)、最大熒光(Fm)、表觀光合電子傳遞速率(ETR)、最大光化學效率(Fv/Fm)、光化學淬滅系數(qP)和非光化學淬滅系數(qN),均由儀器自動測得,依次計算出PSⅡ潛在活性(Fv/Fo),可變熒光(Fv)(Fv=Fm-Fo)。
1.2.4統計分析 利用SPSS 17.0統計軟件進行數據分析。平均值間的比較采用單因素方差分析方法(One-Way ANOVA);不同處理間的差異用Duncan多重比較檢驗(Duncan’s Multiple Range Test)。顯著性水平設定為P=0.05。
2 結果與分析
2.1 Cr3+脅迫下葎草幼苗氣體交換參數的變化
由圖1可以看出,Cr3+為50 mmol/L時,Pn和WUEi比對照顯著增高(圖1A和F),Gs、Ci、Tr、Ls與對照無顯著差異;而200 mmol/L時,Pn則下降至與對照相近,Gs、Ci和Tr也大幅度下降,但Ls和WUEi相反,比對照顯著增加;同時,300 mmol/L時,與對照相比,Pn、Gs、Ci、Tr下降幅度更加顯著,Ls和WUEi仍然上升;然而,500 mmol/L時,Ci、Ls和WUEi卻突然下降(圖1B、E和F),接近對照,但Pn、Gs、Tr仍大幅度降低(圖1A、C和D)。由此可見,葎草幼苗葉片的Pn、Gs、Ci、Tr、Ls、WUEi受到了Cr3+脅迫的顯著影響(P=0.000,P=0.000,P=0.003,P=0.000,P=0.000,P=0.000),Pn、Gs、Ci、Tr隨著Cr3+脅迫的加重而降低,呈下降趨勢,Ls和WUEi呈上升趨勢。
2.2 Cr3+脅迫下葎草幼苗葉綠體色素含量的變化
由表1可以看出,Chl a和Chl b含量隨Cr3+脅迫的增加而逐漸降低,具有顯著差異(P=0.013,P=0.011),其中,Cr3+為50 mmol/L時,二者分別比對照下降16.55%和15.09%,500 mmol/L時二者又減少40.00%和33.96%。同時,Chl a+b和Chl x含量也隨Cr3+脅迫的加重而降低,差異顯著(P=0.009,P=0.002);當Cr3+為50 mmol/L時,二者分別比對照下降16.67%和15.09%,500 mmol/L時二者下降幅度較大,分別為37.88%和43.43%。因此,Cr3+脅迫對葎草幼苗葉片的葉綠體色素含量具有較大影響,Chl a、Chl b、Chl a+b、Chl x含量隨Cr3+脅迫的加劇而逐漸降低,呈下降趨勢,尤其是Cr3+為500 mmol/L時葉綠體色素含量顯著下降。
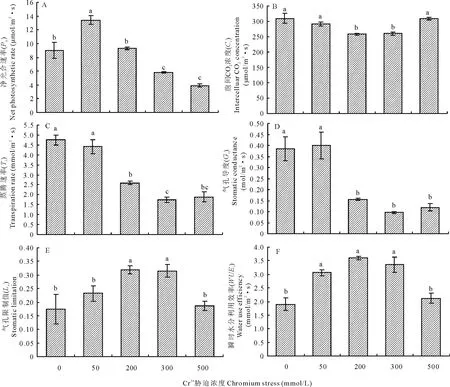
圖1 鉻脅迫下葎草幼苗葉片的氣體交換特征

表1 鉻脅迫對葎草幼苗葉片葉綠體色素和丙二醛(MDA)含量的影響
注:測定值以平均值±標準誤表示,同列不同字母表示差異顯著(P<0.05),下同。Note: Each value is the mean±standard error, different letters in the same column mean significant difference atP<0.05, the same below.
2.3 Cr3+脅迫下葎草幼苗丙二醛(MDA)含量的變化
從表1可以看出,Cr3+為50 mmol/L時,MDA含量比對照下降7.49%,而200和300 mmol/L時MDA含量顯著上升,比對照高14.97%和15.51%,500 mmol/L時卻降低,雖比對照略高,但差異不顯著。從總體上來看,在不同Cr3+脅迫下,葎草幼苗葉片MDA含量存在差異(P=0.020),隨Cr3+濃度的加劇呈上升的變化趨勢。
2.4 Cr3+脅迫下葎草幼苗葉綠素熒光參數的變化
從表2可以看出,在不同Cr3+脅迫下,葎草幼苗葉片葉綠素熒光參數存在顯著差異。50 mmol/L時,Fo′、Fm′、Fo、Fm和Fv均比對照顯著降低,在200和300 mmol/L脅迫時其變化不顯著,但仍然大幅度下降,且出現最小值,而500 mmol/L時,Fo′、Fm′、Fo、Fm和Fv雖比對照仍顯著降低,但均有所增高。另外,ETR截然相反,50 mmol/L時最低(3.10 μmol/m2·s),隨著Cr3+脅迫的加重,ETR逐漸升高,而在300 mmol/L濃度時出現最大值(27.35 μmol/m2·s)。從總體上來看,Cr3+脅迫導致葎草幼苗的Fo′、Fm′、Fo、Fm、Fv和ETR具有顯著影響(P=0.000,P=0.000,P=0.013,P=0.011,P=0.000,P=0.009),Cr3+濃度越高,葎草葉片的Fo′、Fm′、Fo、Fm、Fv逐漸降低,呈下降趨勢,ETR卻逐漸增高,呈上升趨勢。
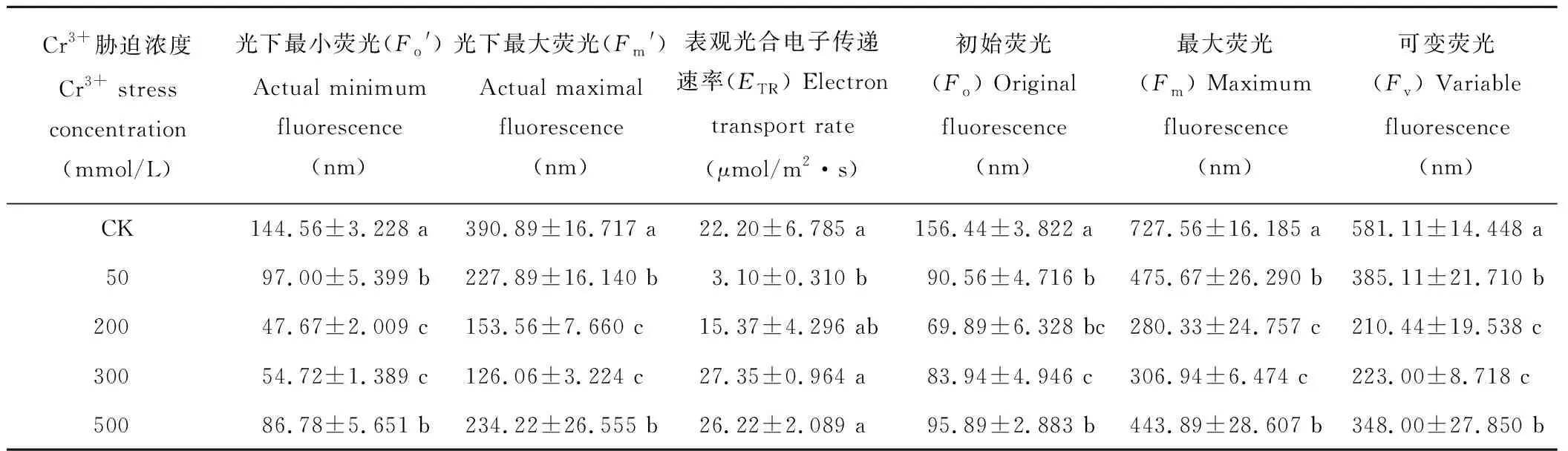
表2 鉻脅迫對葎草幼苗葉片葉綠素熒光參數的影響
2.5 Cr3+脅迫下葎草幼苗光系統Ⅱ反應中心的變化
從圖2可以看出,Cr3+為50 mmol/L時,Fv/Fo、Fv/Fm、qP、qN均比對照增高,但Fv/Fo、Fv/Fm增幅不顯著,而200 mmol/L時,四者卻均比對照降低,其中Fv/Fo和Fv/Fm下降顯著(圖2C和D),且300 mmol/L時,Fv/Fo和Fv/Fm仍然大幅度減少,qP也顯著降低,但qN增幅明顯(圖2A和B);然而500 mmol/L時卻與之相反,Fv/Fo、Fv/Fm、qP、qN均與對照相近,沒有顯著差異,其中Fv/Fo、Fv/Fm、qP突然升高,而qN則又下降。由此可以看出,Cr3+脅迫對葎草幼苗葉片光系統Ⅱ反應中心具有較大影響,其Fv/Fo、Fv/Fm、qP、qN差異顯著(P=0.002,P=0.003,P=0.003,P=0.006),Fv/Fo、Fv/Fm、qP隨Cr3+脅迫的加劇呈下降趨勢,qN變化無規律,但重度脅迫(500 mmol/L)時,Fv/Fo、Fv/Fm、qP、qN均與對照接近。
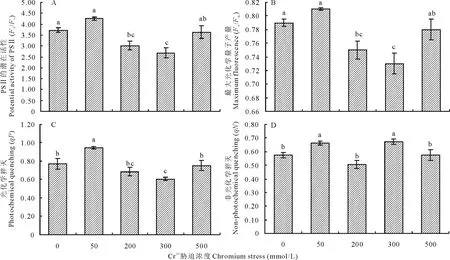
圖2 鉻脅迫下葎草幼苗葉片的光系統Ⅱ反應中心特征
3 討論
3.1 低濃度Cr3+脅迫對葎草幼苗光合生理特性的影響
植物葉片凈光合速率(Pn)的大小是衡量植物光合作用能力強弱的重要指標。本試驗結果表明,葎草幼苗在低濃度(50 mmol/L)Cr3+下Pn和WUEi顯著增高,而Gs、Ci、Tr和Ls均沒有變化。Ci和Gs是反映氣孔開張程度的重要指標,其值的變化反映了葉肉細胞內外CO2濃度的變化狀況。在本研究中,低濃度Cr3+(50 mmol/L)處理下葎草幼苗的光合速率顯著增加,而Gs和Ci均無顯著變化。因此,我們推測低濃度Cr3+脅迫下葎草凈光合速率的增高與氣孔行為無關,而極可能與光合色素、光系統中的電子傳遞能力和酶的活性增強有關。進而,我們對葉片色素含量以及葉綠素熒光參數的測定結果發現,低濃度Cr3+脅迫下葎草葉綠素a、b以及總色素含量均無顯著變化,而qP和qN均比對照顯著增加。根據陳建明等[18]的解釋,光化學淬滅系數qP反映了PSⅡ原初電子受體QA的還原狀態,qP愈大,PSⅡ的電子傳遞活性愈大;而qN反映的是P天線色素吸收的不能用于光合電子傳遞而以熱形式耗散掉的光能部分,是反映光合機構自我保護機制的指標。由此證實了低濃度Cr3+脅迫可能通過促進了PSⅡ電子傳遞活性來增加了凈光合速率。因此,我們的研究結果表明了低濃度的Cr3+對植物的生長具有一定的刺激作用。該結論與許建光等[19]認為微量鉻能提高植物體內某些酶的活性而促進植物生長的觀點完全一致。
3.2 中濃度Cr3+脅迫對葎草幼苗光合生理特性的影響
在中濃度(200~300 mmol/L)Cr3+脅迫下,我們發現隨著Cr3+脅迫的加重,Pn、Gs、Ci和Tr均下降,Ls上升。根據Farquhar和Sharkey[20]的觀點:當Pn下降,Ci也減小,而Ls增大,可以認為Pn的下降主要是由于Gs的降低;如果Pn下降,Ci升高,而Ls減少,便可以肯定葉肉細胞光合活性的降低是Pn下降的決定因素。因此,我們可以確定在中度Cr3+脅迫下Pn顯著下降的原因可能與氣孔限制所引起CO2進入葉片受阻有關。然而,Ls與WUEi變化趨勢卻相似,隨著Cr3+濃度的增加而增加,其原因可能是氣孔減少對H2O分子的影響比CO2更大,受到脅迫時氣孔部分關閉,導致水分利用效率提高。此外,光合色素以及葉綠素熒光參數的測定結果表明,在中濃度(200~300 mmol/L)Cr3+脅迫下,與對照相比葎草葉片的葉綠素a、葉綠素b、總色素含量、類胡蘿卜素含量以及光系統Ⅱ的潛在活性(Fv/Fo)、最大光化學量子產量(Fv/Fm)、光化學淬滅系數(qP)和非光化學淬滅系數(qN)均顯著下降。Fv/Fm是開放的PSⅡ反應中心捕獲激發能的效率,是反映植物PSⅡ受傷害程度的重要參數[21]。在中濃度Cr3+脅迫下,Fm、Fv、Fv/Fo下降反映了葎草幼苗的PSⅡ反應中心開放程度減弱,光合電子傳遞速度減慢,不僅碳同化速率降低,而且Rubisco和PSI與PSⅡ間的電子傳遞位點還可能遭到破壞[22]。
此外,Pandey和Sharma認為[23],Cr3+在植物體內沒有一定的轉運途徑,而是通過和鐵競爭轉運蛋白才能到達植株上部,這樣葉片中的鐵含量會減少,造成葉綠素合成量減少[19]。所以,我們的研究結果進一步證實了中濃度的Cr3+脅迫可以對葉綠素合成產生嚴重影響,使得葉綠素含量顯著降低,導致光化學反應過程中PSⅡ的酶和電子傳遞也受到顯著影響。
3.3 高濃度Cr3+脅迫對葎草幼苗光合生理特性的影響
在高濃度(500 mmol/L)Cr3+脅迫下,我們發現葎草植株的Pn、Gs、Tr與對照相比顯著降低,尤其是光合色素Chl a、Chl b、Chl a+b和Chl x的含量隨濃度越高其值顯著降到最低,而氣孔限制值(Ls)與胞間CO2濃度(Ci)卻無顯著變化。由此反映出隨Cr3+濃度的加重,Cr3+與鐵競爭轉運蛋白的現象加劇[23],葉綠素合成量不僅受到影響,甚至影響其光合功能,從而導致了Pn的顯著下降。葎草光合色素含量的這種變化趨勢,與二月蘭(Orychophragmusviolaeeus)、玉米(Zeamays)的葉綠素含量的變化規律完全吻合[24-25]。另一方面,我們卻發現葉綠素熒光參數Fv/Fo、Fv/Fm、qP和qN以及反映膜脂過氧化程度的MDA含量與對照相比卻無顯著變化(表1,圖2),這可能是大量Cr3+干擾原有離子間的平衡系統,致使PSⅡ反應中心不可逆失活,引起Fo增加,QA處于氧化還原狀態,Fv隨光合活性而變化,為了保護PSⅡ免遭破壞,非輻射能量耗散也隨之增強,因此qP上升,qN下降,我們猜測也許是防御PSⅡ過量激發或結構嚴重損傷的某種機制,或是細胞內積累的Cr3+通過其他方式影響植物的生理生化活動,與部分大分子物質結合,取代了某些必需的特定元素,使其變性或活性降低,這也可從較低水平的MDA得到印證。作為檢測植物受逆程度的重要指標,多數的研究結果表明,脅迫環境下植物的MDA含量常常高于正常生境中的植物。然而,在極高脅迫環境下 MDA含量的變化規律研究尚少。在本實驗過程中,我們發現當Cr3+脅迫下的濃度升至500 mmol/L時,MDA含量與對照組的含量相比并沒有顯著變化,這與前人的結果似乎并不一致[4-6]。
另一方面,部分研究也發現了植株MDA含量在高脅迫下呈現降低現象。如高麗萍等[26]在獼猴桃(Actinidiachinensis)的衰老試驗中,觀察到MDA 含量先升后降。此外,覃鵬等[27]的研究也證實了在干旱脅迫下各個煙草(Nicotianatabacum)品種(系)的MDA含量總體上隨著干旱脅迫的不斷加重而呈現出先升后降的趨勢,其原因估計是高脅迫程度下植株嚴重受害,葉片全部萎蔫而不能進行正常的生物合成,反而不斷消耗干物質,導致MDA不僅不能產生,甚至處于持續的降解之中,所以隨著干旱處理的加重而呈現下降趨勢。多數的研究認為,鉻可引起永久性的質壁分離并使植物組織失水,高濃度的鉻對線粒體的傷害是不可逆轉的,將造成呼吸作用降低,甚至植物細胞死亡[28-30]。因此,我們推測高濃度鉻脅迫導致植株根部細胞發生永久性的質壁分離并使植物組織失水,植物體內的代謝過程發生紊亂,化學成分發生改變,引起部分細胞的死亡,最終導致MDA含量不僅沒有隨脅迫加強而繼續增高,而是相應地發生了降解,造成含量降低。在實驗中我們觀測到高濃度鉻脅迫下植株在7 d內很快出現萎蔫的現象,這也間接地證明了推測的合理性。
綜上所述,低濃度Cr3+脅迫可以通過促進PSⅡ電子傳遞活性來增加葎草幼苗的凈光合速率,從而在一定程度上促進植株生長。中等濃度Cr3+脅迫則嚴重影響了植株氣孔的擴張行為,降低了光合色素含量,同時減弱了PSⅡ反應中心開放程度以及光合電子的傳遞速度,從而導致葎草幼苗的凈光合速率顯著下降,植株生長受到抑制。高濃度Cr3+脅迫則能導致植物體內的代謝過程發生紊亂,引起部分細胞的死亡,光合色素含量下降,凈光合速率降到最低,植株生長受到嚴重影響。此外,由于Cr3+毒害是多方面的,其抗性機理十分復雜,Cr3+脅迫影響植株生長的關鍵途徑及因素還有待進一步深入研究。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
