菌劑對鉻(Ⅵ)污染土壤中坪草幼苗生理生化的影響
2014-11-08 03:20:44高海寧馬國泰李彩霞陳勇宋濤張勇焦揚
草業學報 2014年4期
高海寧,馬國泰,李彩霞,陳勇,宋濤,張勇,4,焦揚,4*
(1.河西學院農業與生物技術學院,甘肅 張掖 734000; 2.蘭州大學生命科學學院,甘肅 蘭州 730000;3.甘肅省鵬宇生態環保工程公司,甘肅 蘭州 730000;4.甘肅省高校河西走廊特色資源利用省級重點實驗室,甘肅 張掖 734000)
鉻鹽是重要的化工原料,廣泛應用于電鍍、化工、皮革、耐火材料等行業。鉻鹽生產及其產品應用的各個環節都可能產生含Cr廢水。由于Cr(Ⅵ)具有致癌性、致突變性、對生物體具有很強的毒性,被列為國際公認的3種致癌金屬物之一[1-2]。我國是鉻產品生產和使用大國,因此Cr(Ⅵ)污染治理是一個長期而艱巨的任務。我國人口壓力大,耕地資源緊張,安全有效地修復重金屬污染土壤,既可實現土地資源的再利用,同時,又可避免重金屬通過食物鏈遷移。相對于Cr(Ⅵ),Cr(Ⅲ)毒性較低,對環境影響較小,因此土壤Cr(Ⅵ)污染的修復機制通常有兩種[3-4]:一是改變Cr在土壤中的存在形態,將Cr(Ⅵ)還原為Cr(Ⅲ),降低其在環境中的遷移能力和生物可利用性;二是將Cr從被污染土壤中清除。
Cr(Ⅵ)污染土壤的微生物修復是利用土壤中的土著微生物或經馴化的特定微生物,通過將Cr(Ⅵ)還原為Cr(Ⅲ),達到降低Cr的移動性和毒性等目的[5]。鉻渣堆場鉻污染土壤中土著微生物對Cr(Ⅵ)具有很強的還原能力,直接向土壤中添加營養物質可刺激土著微生物活性,將Cr污染土壤中Cr(Ⅵ)還原成Cr(Ⅲ)進行Cr污染土壤的微生物修復[6]。但從機理上,微生物不利于收集或富集重金屬,因而,單獨應用微生物修復金屬污染土壤受到限制[7]。通過耐Cr植物對Cr污染場地的綠化與覆蓋,控制Cr向深層土壤移動是控制Cr(Ⅵ)污染的重要措施[8]。孟慶恒等[9]篩選出4株對Cr(Ⅵ)的清除率大于50%的土著菌株,并與玉米(Zeamays)幼苗組合進行微生物-植物聯合修復,具有明顯降低培養液中Cr(Ⅵ)濃度的效果,因此,研究微生物對重金屬污染修復具有重要的現實意義。
高羊茅(Festucaarundinacea)和黑麥草(Loldiumperenne)是兩種常見的多年生冷季型草坪草,也是溫帶地區重要的禾草類牧草。發現高羊茅和黑麥草具有吸收不同重金屬的能力,楊卓等[10]對兩種坪草富集Zn、Cd、Pb的能力進行了研究,發現兩種坪草對Zn污染土壤的修復能力較強;張蕾等[11]通過盆栽試驗,研究黑麥草對復合污染河道浚底泥的修復效果,結果表明,黑麥草對Zn、Cd、Cu的富集能力強,是修復重金屬-有機復合物的良好植物;趙樹蘭和多立安[12]研究Cu2+與Zn2+對高羊茅生長的影響,結果顯示高羊茅根系和莖葉具有富集重金屬的能力;Zhu等[13]研究尾礦污染土壤對高羊茅和早熟禾(Poaannua)生長的影響,發現這些坪草對重金屬具有吸耐能力。以上研究主要集中在坪草對重金屬污染土壤的修復等方面,或者研究對象單一,并未涉及菌劑對鉻(Ⅵ)污染土壤中坪草幼苗生理方面的相關報道。
本研究以高羊茅和黑麥草為材料,以Cr(Ⅵ)污染土壤為基質,以期檢測菌劑處理對Cr(Ⅵ)污染土壤中高羊茅和黑麥草幼苗生理生化特性的影響,擬為Cr污染場地的微生物-植物聯合治理提供技術與理論依據。
1 材料與方法
1.1 實驗材料
1.1.1菌劑 從張掖周邊鹽堿化和Cr污染土壤中篩選到革蘭氏陰性芽孢桿菌Rs-2和K1,革蘭氏陽性芽孢菌Rs-4。3株菌具有在鹽堿環境中溶磷、硅的特性[14],且Rs-2、K1菌株可將Cr(Ⅵ)還原為Cr(Ⅲ)。將菌株發酵得到高密度菌液,以Rs-2∶Rs-4∶K1=9∶6∶4比例得到混合菌液,加入1.5%的載體材料和0.1%的甘油,制備得到復合菌劑,以培養基代替菌液再加入1.5%的載體材料和0.1%的甘油作為對照。
1.1.2供試土壤與植物 Cr(Ⅵ)污染土壤采自甘肅省張掖市民樂縣富源化工有限公司污水口周邊。Cr(Ⅵ)污染土壤采回后錘碎,挑除石子,備用。供試植物為高羊茅和黑麥草。
1.2 實驗方法
1.2.1培養基 菌液發酵培養基(g/L):蛋白胨10.0,牛肉膏5.0,NaCl 5.0,MgSO40.2,pH自然。
1.2.2實驗設計 實驗于2012年3-7月在河西學院植物生理實驗室進行。供試土壤按80∶10∶1的比例將Cr(Ⅵ)污染土壤∶有機肥(羊糞與玉米芯堆肥)∶菌劑混合均勻,加適量水后密封,每天攪拌一次,7 d后土壤處理完成。將處理后的Cr(Ⅵ)污染土壤均勻裝入育苗盤中。每穴播種經30% H2O2表面殺菌種子20粒,然后覆蓋1 cm左右處理過的土壤,澆透水至育苗盤底滲出水為止,每個處理重復3個育苗盤(4×9=36孔),每天進行管理,使土壤保持濕潤,待兩種坪草出芽后對植株進行間苗,每缽保留10株。當植物幼苗生長至3葉齡時,對照組(CK)澆灌稀釋5倍的對照液,處理組澆灌稀釋5倍的菌劑,在植物幼苗處理6 d后,測定幼苗生理指標,每項指標做3組平行。
1.3 分析方法
1.3.1土壤指標測定方法 土壤pH值的測定采用酸度計法;可溶性鹽的測定采用質量法;土壤Cr(Ⅵ)含量的測定采用二苯碳酰二肼比色法[15]。
1.3.2生化指標測定方法 稱取植物葉片0.2 g,加入50 mmol/L pH 7.8的磷酸緩沖液研磨,4℃、10000 r/min離心10 min,上清液定容至5 mL,得粗酶液備用。POD(peroxidase,過氧化物酶)活性測定采用愈創木酚法;SOD(superoxide dismutase,超氧化物歧化酶)活性測定采用氮藍四唑光還原法;CAT(catalase,過氧化氫酶)活性測定采用紫外分光光度法;葉綠素含量測定采用分光光度法;脯氨酸含量測定采用酸性茚三酮比色法;細胞膜透性測定采用電導法;丙二醛(MDA)的測定采用2-硫代巴比妥酸法;植物組織根部、地上部分Cr含量測定采用火焰原子吸收分光光度法[16]。
根據植物根、葉組織Cr的含量計算生物富集系數:生物富集系數=坪草鉻含量(mg/kg DW)/土壤鉻含量(mg/kg)。
1.4 數據處理
每個處理做3次重復,以平均值±標準偏差表示各指標的大小,采用SSR法檢驗差異顯著性。Origin Lab 7.5軟件進行數據作圖。
2 結果與分析
2.1 供試土壤處理前后的變化
由表1可見,供試土壤被菌劑處理后,其pH從7.38增長為8.25,而可溶性鹽和Cr(Ⅵ)分別從6.20%和53.9 mg/kg降為5.25%和23.6 mg/kg,

表1 供試土壤處理前后的指標變化
注:采用Duncan’s multiple range test 方法分析,同列不同字母表示顯著性差異(P<0.05,n=3),下同。Note: Analysis using Duncan’s multiple range test method, the different letters in the same column show significant difference (P<0.05,n=3), the same below.且供試土壤被菌劑處理前后Cr(Ⅵ)的變化差異顯著。
2.2 菌劑對坪草幼苗根和地上部分Cr含量的影響
由表2可知,污染土壤中Cr被坪草吸收后,大部分停留在根部,少量向地上部分遷移。CK組較處理組葉和根中的Cr含量差異均顯著。CK組坪草葉中Cr含量為根中Cr含量的11%~14%;而處理組葉中Cr含量為根中Cr含量的31%~39%。由生物富集系數可見,CK組兩種坪草葉Cr的生物富集系數為0.62和0.50,而處理組的生物富集系數增大至1.10和1.05,由此,處理前后根中Cr含量在降低,而葉中Cr含量在增加。且兩坪草根和葉對Cr的富集有差異,其中高羊茅對Cr富集略高于黑麥草,但差異不顯著。
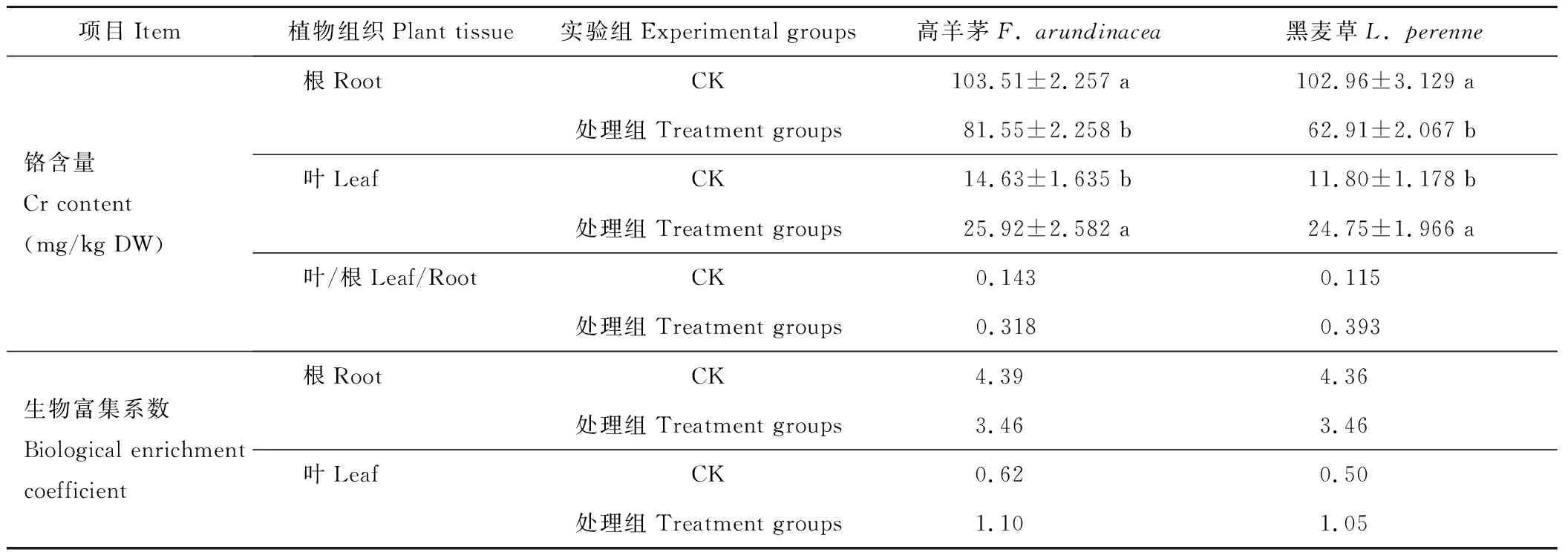
表2 坪草幼苗根系、地上部分鉻的含量
2.3 菌劑對坪草幼苗葉綠素含量的影響
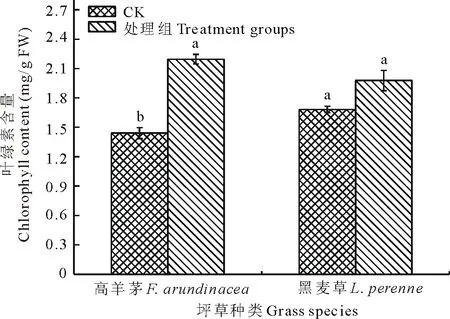
圖1 Cr(Ⅵ)污染土壤中坪草幼苗葉綠素含量
從圖1可見,CK組兩坪草幼苗葉綠素的含量是1.44和1.68 mg/g FW,經菌劑處理后,高羊茅和黑麥草幼苗總葉綠素含量增加至2.19和1.98 mg/g FW。兩坪草處理組葉綠素含量均高于CK組,而且高羊茅的這種變化差異顯著,可見,在坪草幼苗生長過程中輔以菌劑處理,可使幼苗葉綠素含量增加。
2.4 菌劑處理對坪草幼苗保護酶系的影響
由表3可見,菌劑處理后,高羊茅和黑麥草的SOD和CAT都高于CK組,處理組是CK組的1.13,1.47倍和1.19,1.04倍,但處理組較CK組,兩坪草的POD、CAT以及高羊茅的SOD差異均不顯著,而黑麥草的SOD處理前后差異顯著。加菌劑處理后,高羊茅的POD活性較CK組降低,處理組是CK的0.85倍。
2.5 菌劑處理對坪草幼苗脯氨酸、丙二醛和相對電導率的影響
由表4可知,處理組較CK組,黑麥草的MDA、脯氨酸和相對電導率均有一定程度的降低,且差異顯著。高羊茅的處理組較CK組,脯氨酸變化差異顯著,雖MDA和相對電導率有一定程度降低,但差異不顯著。由此,經菌劑處理后兩坪草的脯氨酸含量降低較明顯,黑麥草的MDA和相對電導率較高羊茅的降低更為顯著。
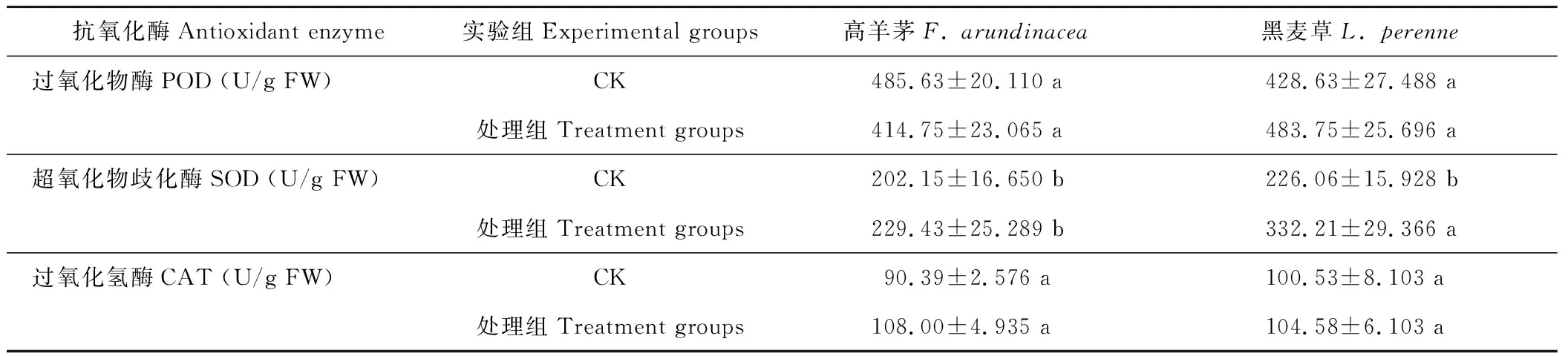
表3 菌劑對Cr(Ⅵ)污染土壤坪草幼苗細胞保護酶活性的影響
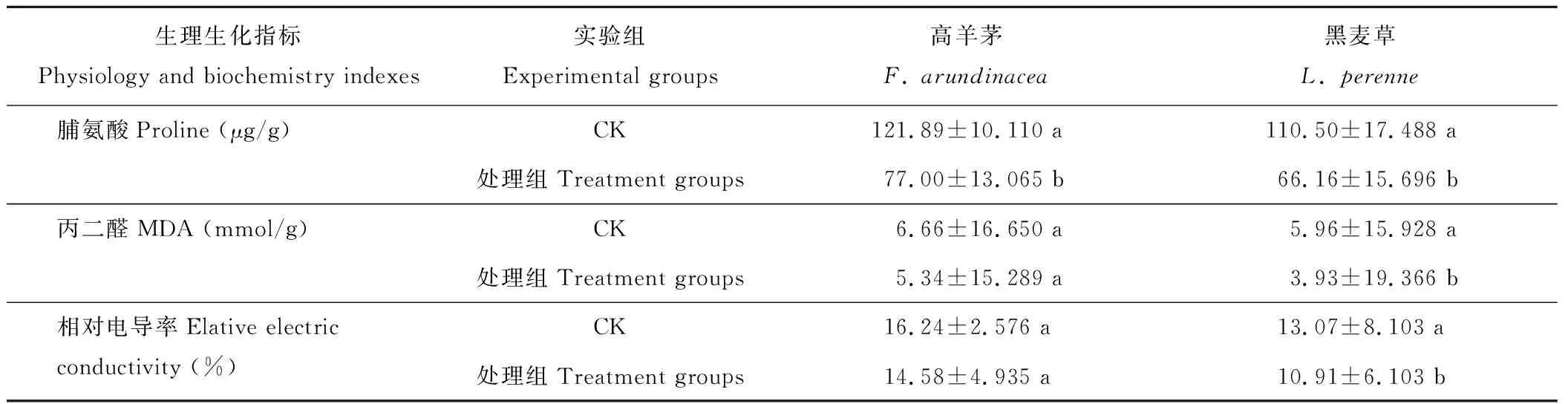
表4 菌劑對Cr(Ⅵ)污染土壤坪草幼苗脯氨酸、丙二醛和相對電導率的影響
3 討論
常文越等[2]在25℃、投加2%的還原菌,在施用菌劑1個月后,土壤浸出液Cr(Ⅵ)濃度范圍從10~55 mg/L降低至0.75 mg/L以下。在實驗中對供試土壤采用菌劑處理后,污染土壤中Cr(Ⅵ)的濃度從53.9 mg/kg降為23.6 mg/kg,且土壤pH略升而含鹽量略降,可能是菌劑的還原作用降低了Cr(Ⅵ)在污染土壤中的含量,因此,菌劑對Cr(Ⅵ)污染土壤具有一定的改良效果。
重金屬元素能否向植物地上部分,特別是向葉片部分的遷移是判斷超積累植物的重要標準之一[17]。重金屬在植物體內的積累有很大的差異,不同植物對重金屬的策略也不一致,有的采用富集策略,也有的采用排斥策略[18]。Zayed等[1]研究發現,在使用Cr(Ⅵ)培養液后,供試作物根中含量為160~350 mg/kg DW,莖中含量為1.6~2.0 mg/kg DW,即重金屬Cr在植物體內新陳代謝旺盛的器官中蓄積量較大[9]。實驗中,當供試植物在Cr(Ⅵ)污染土壤中生長時,在幼苗期輔以菌劑處理,Cr在高羊茅和黑麥草根中的積累從103.51和102.96 mg/kg降為81.55和62.91 mg/kg,但兩種坪草葉對Cr的生物富集系數從0.62和0.50增長為1.10和1.05。由此可得,菌劑處理對緩解Cr在新陳代謝旺盛的器官中的蓄積有明顯效果,而且可以提高Cr在坪草地上部分,尤其是葉中的生物富集量。
葉綠體是植物細胞所特有的能量轉換細胞器,它受重金屬的影響比較明顯。王愛云和黃珊珊[19]以外源Cr(Ⅵ)濃度在0~300 mg/kg范圍內模擬Cr污染土壤,高羊茅幼苗總葉綠素含量從3.43 mg/g降為2.81 mg/g。本實驗中菌劑處理后,高羊茅幼苗葉中總葉綠素含量從1.44 mg/g FW增加至2.19 mg/g FW,葉綠素總量低于報道值的3.43 mg/g,可能源于Cr(Ⅵ)污染土壤成分更為復雜,且有5.25%的鹽含量,但菌劑處理后將幼苗葉綠素總量提高了52.1%。因此,幼苗期輔以菌劑處理可以緩解坪草在Cr(Ⅵ)脅迫條件下幼苗葉綠素的降低程度,但不同坪草提高幅度有差異。
POD、SOD和CAT是植物體內酶促防御系統的3個重要保護酶,當植物處于逆境條件下,POD、SOD、CAT保護酶通過協調作用能有效清除OH-,O2-等自由基,防御膜脂過氧化,使植物細胞膜免受傷害。SOD和CAT活性的同時提高十分重要,由于SOD在清除自由基時,同時生成H2O2自由基,它可以通過Fenton型Haber-Weiss反應生成更多的自由基,CAT活性提高能與SOD協同作用更好地保護細胞膜免受自由基的傷害[20]。本研究結果表明,菌劑處理后,高羊茅和黑麥草的SOD和CAT活性都高于CK組,處理組是CK的1.13,1.47倍和1.19,1.04倍,表明在植物幼苗期輔以菌劑處理,可增加高羊茅和黑麥草的SOD和CAT活性來清除由Cr而引起的氧化傷害,與高羊茅相比較,黑麥草SOD活性增幅更高。
孫健等[21]報道重金屬脅迫能誘導植物組織中POD總活性升高,這是對所有污染脅迫的共同響應。POD利用H2O2來催化對自身有害過氧化物的氧化和分解[22];許長成等[23]報道大豆(Glycinemax)在干旱條件下,大豆葉片H2O2含量增加,CAT活性降低,POD活性隨著升高,由此POD活性的變化與干旱條件的H2O2有關。本研究發現在Cr處理下,菌劑對高羊茅的POD活性的影響有所不同,高羊茅在不加菌劑時,其葉片中POD活性達到了485.6 U/g FW,而加菌劑后使高羊茅POD活性下降,但是CAT活性增加,推測Cr(Ⅵ)污染土壤高羊茅幼苗體內CAT活性強于POD,H2O2的積累量減少,從而使POD活性出現下降。而黑麥草加入菌劑后POD、CAT、SOD活性均高于CK組,三者酶協同作用,降低Cr的氧化損傷,使得MDA含量和相對電導率顯著低于對照組,保護了膜的完整性,但相比較而言,高羊茅幼苗體內MDA含量和相對電導率降低幅度較小。
脯氨酸在植物滲透調節中起著重要的作用。脯氨酸的合成、累積及代謝是一個受非生物脅迫和細胞內脯氨酸濃度調控的生理生化過程[24-25]。脯氨酸含量升高,是植物受到重金屬毒害的一個重要特征。張小艾等[26]認為,對于脯氨酸含量在逆境下增加是一種傷害反應的觀點不一,其機理有待進一步研究。本實驗中,CK組的Cr(Ⅵ)污染土壤中兩種坪草脯氨酸含量顯著高于菌劑處理組,說明在Cr脅迫下脯氨酸含量的升高是維持細胞滲透勢,對植物細胞具有保護作用,輔以菌劑后脯氨酸含量降低,而保護酶活性升高,MDA和細胞電導率的處理組/CK均小于1,降低了Cr(Ⅵ)污染土壤對黑麥草和高羊茅幼苗細胞膜不飽和脂肪酸的破壞程度和氧化損傷,減緩了Cr(Ⅵ)對兩種坪草的毒害作用[27]。
4 結論
在坪草幼苗期輔以菌劑處理,對緩解Cr在新陳代謝旺盛器官中的蓄積有明顯效果,而且可以提高Cr在坪草地上部分,尤其是葉中的生物富集量;并能夠降低Cr(Ⅵ)污染土壤坪草幼苗的質膜透性和MDA含量,提高POD、SOD和CAT保護酶活性。采用菌劑處理Cr(Ⅵ)污染土壤,可降低Cr(Ⅵ)污染土壤中Cr(Ⅵ)的含量,對Cr(Ⅵ)污染土壤具有一定的修復和改良效果。
猜你喜歡
小學科學(學生版)(2021年7期)2021-07-28 06:44:32
小學科學(學生版)(2020年6期)2020-08-13 07:47:12
當代陜西(2019年7期)2019-04-25 00:22:18
領導決策信息(2018年26期)2018-10-12 02:18:26
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
