煙草靶斑病菌(Rhizoctonia solani)SRAP-PCR 體系建立及優化
2014-11-24 05:11:22趙艷琴吳元華趙秀香安夢楠陳建光
中國煙草學報 2014年3期
趙艷琴,吳元華,趙秀香,安夢楠,陳建光
1沈陽農業大學/植物保護學院,遼寧沈陽市東陵路120號 110866;
2 內蒙古民族大學/農學院,內蒙古通遼霍林河大街22號 028043
煙草靶斑病菌(Rhizoctonia solani)SRAP-PCR 體系建立及優化
趙艷琴1,2,吳元華1,趙秀香1,安夢楠1,陳建光1
1沈陽農業大學/植物保護學院,遼寧沈陽市東陵路120號 110866;
2 內蒙古民族大學/農學院,內蒙古通遼霍林河大街22號 028043
采用煙草靶斑病菌YC-9, LJT-8和QYS-7為DNA模板,初步篩選SRAP引物組合;采用L16(45)正交試驗設計,對煙草靶斑病菌的SRAP-PCR反應體系中的Mg2+、dNTPs, Taq DNA聚合酶、引物和DNA模板濃度等5個因素進行優化試驗。結果表明:共篩出13對擴增條帶清晰且多態性好的引物組合;煙草靶斑病菌的最佳SRAP反應體系為Mg2+濃度2.0 mmol/L、dNTP濃度200 μmol/L、Taq DNA聚合酶0.8 U、引物濃度140 mmol/L、模板DNA 20 ng及1×PCR buffer,反應總體積為20 μL;各因素對SRAP-PCR擴增反應結果影響的差異較大,依次為Taq DNA聚合酶>引物> Mg2+> dNTPs=模板DNA。
煙草靶斑病菌;正交試驗設計;反應體系優化;引物篩選
SRAP (Sequence related amplified polymorphism)技術是2001年Li和Quiros在蕓苔屬植物上開發出的新型分子標記技術[1]。該技術無需任何序列信息即可直接 PCR 擴增,從DNA 水平上直接檢測基因組DNA 的多態性[2],相比此前廣為應用的RAPD(Random amplification polymorphism DNA)技術以及AFLP (Amplified fragment length poly-morphism)等技術,SRAP技術穩定性和重復性好,并且成本低,簡單快速[3]。目前SRAP技術已成功在多種植物遺傳圖譜和多樣性分析的研究中應用[4-5],以及在植物病原物的遺傳多樣性研究中應用[6-7]。
煙草靶斑病(Thanatephorus cucumeris(Frank)Donk)在許多國家都有發生[8-10],是一種危害嚴重的煙草葉部病害。我國煙草靶斑病主要發生在煙草旺長期和成熟期,潛育期短、流行性強,嚴重影響煙葉的品質和產量[11]。煙草靶斑病菌的無性態為立枯絲核菌(Rhizoctonia solani),目前針對我國煙草靶斑病菌的研究報道較少[12-14]。為明確煙草靶斑病菌的遺傳多樣性及進化背景,本文建立了煙草靶斑病菌SRAPPCR反應體系,并對引物進行了篩選,以期早日明確煙草靶斑病菌的遺傳本質,為該病害的防治提供理論依據。
1 材料與方法
1.1 試驗菌株
煙草靶斑病菌株YC-9,LJT-8和QYS-7,分別分離自鐵嶺市營廠鄉,李家臺鄉及丹東市青椅山煙草靶斑病菌組織,保存于沈陽農業大學煙草研究所。
1.2 試驗方法
1.2.1 基因組DNA提取與檢測
首先將菌株移植于PDA平板上,在28 ℃培養箱內培養3 d后,取5 mm直徑的菌餅至含有10 mL的PD培養液的培養皿(9 cm)中,于28 ℃下靜置培養3 d,過濾出菌絲,用無菌水沖洗,無菌濾紙吸干水分后于40℃干燥4 h后裝入1.5 mL的離心管中,-20℃保存備用。
DNA提取采用北京天根生物技術公司DNA提取試劑盒按照說明進行,并采用1%瓊脂糖凝膠電泳檢測DNA。
1.2.2 SRAP引物篩選
SRAP引物采用Li等已發表的序列Me1~Me10/Em1~Em10[1],由上海生工生物有限公司合成。選取菌株YC-9、LJT-8和QYS-7,對100對SRAP引物進行多態性初篩。
1.2.3 SRAP-PCR擴增反應體系及程序
基礎反應體積為20 μL:l0×PCR buffer (Mg2+)2 μL, Mg2+0.2 μL, 10 mmol/L dNTPs (10 mmol/L Each) 1.6 μL, 正 反 向 引 物 (10 μmol/L)各 0.5 μL, TaqDNApolymerase (2.5 U/μL) 0.32 μL,模板DNA1.0 μL,用ddH2O定容至20 μL。
反應程序參照Li等的方法略加修改[1], 94 ℃ 5 min; 94 ℃ 1 min, 35 ℃ 1 min, 72 ℃ l min, 4 個循環 ;94 ℃ 1 min, 56 ℃ 1 min, 72 ℃ 1 min, 39 個循環 ;72 ℃10 min, 4 ℃保存。
擴增結束后,取7.5 μL擴增產物,與1.5 μL 6×Loading Buffer(TaKaRa)混勻,點入含0.5 μg/mL Goldview 的1.5% 瓊脂糖凝膠中,以 DNA Marker DL2000(Tiangen)作為分子量標準,在 5 V/cm 電場強度下電泳 1~2 h,紫外凝膠成像儀下照相。
1.2.4 SRAP 反應體系優化
選取質量較好的菌株DNA作為模板及多態性好的一組SRAP引物進行反應體系優化試驗。首先按照表1進行單因素優化,以確定各因子的適宜濃度范圍,在此基礎上參照表2采用五因素四水平L16(45)進行正交優化。參考張麗等分析統計方法[15],對 Mg2+、dNTPs、Taq DNA 聚合酶、引物以及模板DNA濃度水平進行優化以獲得穩定的反應體系。

表1 單因子試驗設計表Tab.1 Single-factor experiment
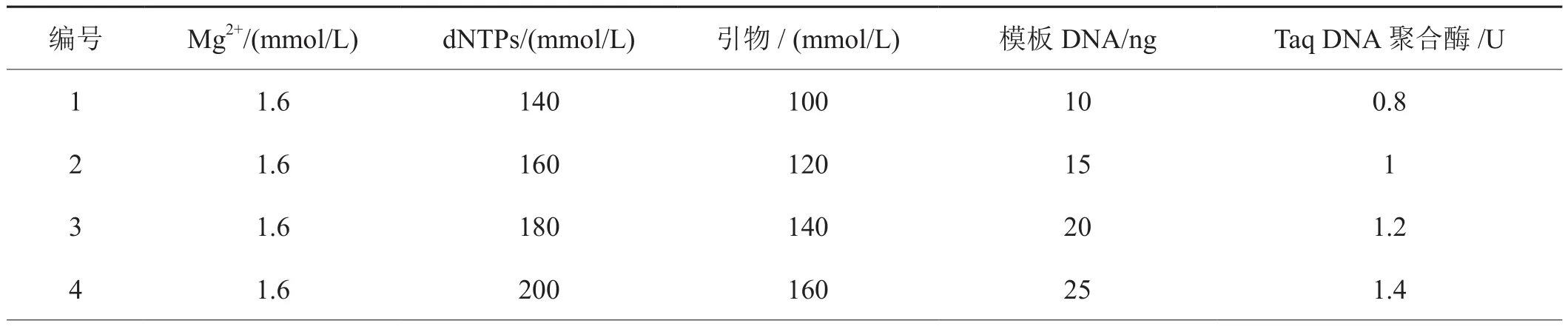
表2 煙草靶斑病菌SRAP-PCR反應體系的L16(45)正交試驗設計表Tab.2 An orthogonal design of L16(45) of SRAP-PCR reaction system for Rhizoctonia solani from tobacco target spot
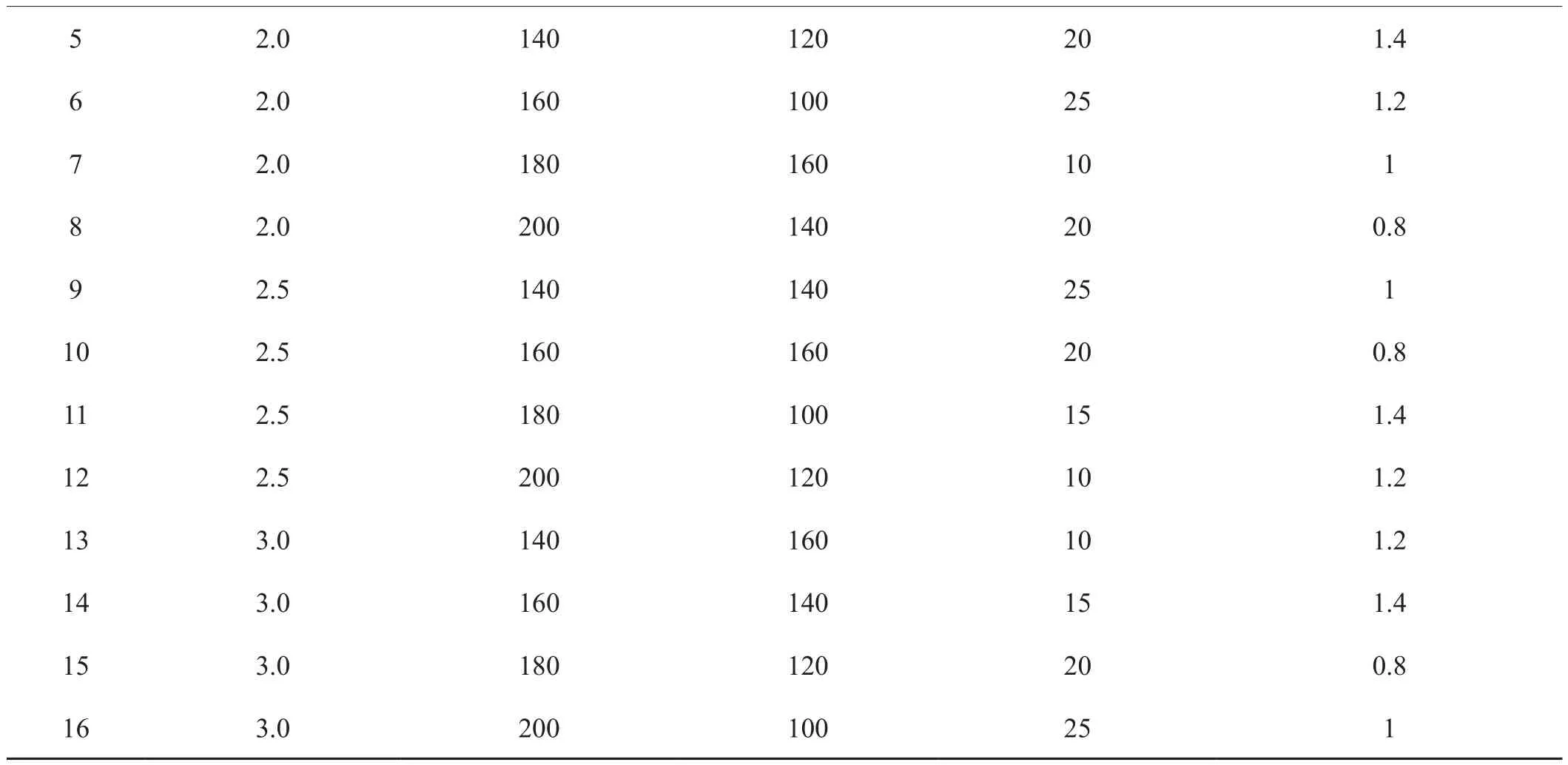
表2(續)
2 結果與分析
2.1 SRAP引物篩選
選取YC-9、LJT-8和QYS-7菌株對100對SRAP引物進行篩選,圖1結果表明:篩選獲得13組條帶清晰且多態性好的引物組合,分別是Me1/Em2、Me3/Em2、Me3/Em9、Me4/Em4、Me4/Em6、Me4/Em9、Me5/Em3、Me5/Em9、Me6/Em2、Me6/Em8 和Me7/Em10 Me8/Em7和 Me9/Em5。

圖1 優化SRAP反應體系部分引物中的擴增結果Fig.1 Amplified results of partial primer in the optimization of SRAP reaction system
2.2 SRAP反應體系優化
選擇煙草靶斑病菌YC-9 基因組DNA及SRAP引物組合Me7/Em10進行體系優化。
2.2.1 Mg2+濃度
圖2試驗結果表明:當Mg2+濃度為1.0 mmol/L時,僅有較少的擴增條帶數和擴增量;當Mg2+為1.5~2.5 mmol/L時,能獲得較清晰的譜帶且譜帶數量增多,其中以2.5 mmol/L 的Mg2+濃度最佳;當Mg2+為3.0~4.5 mmol/L時,隨著Mg2+濃度的增加擴增條帶數變化不明顯,但擴增量逐漸減少;至Mg2+為5 mmol/L時未擴增出譜帶。
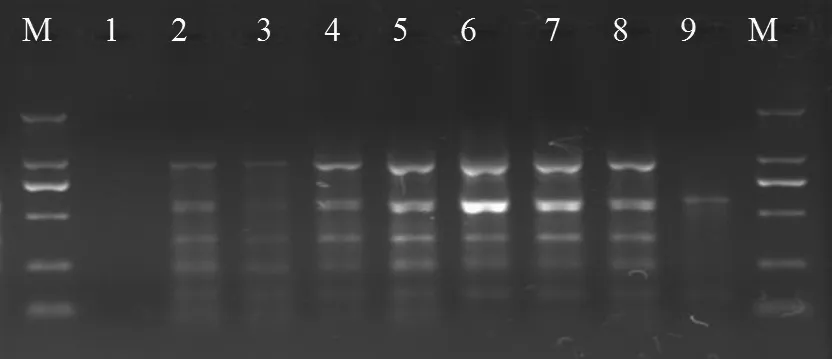
圖2 Mg2+濃度對PCR擴增的影響Fig.2 Effect of Mg2+ density on the amplification of PCR
2.2.2 dNTPs濃度優化
圖3表明:當dNTPs 為100 mmol/L時,獲得譜帶少且模糊不清,隨著濃度增加,條帶數增加,且清晰度增強,至dNTP的濃度為200 mmol/L時獲得達到最多,最清晰;而當濃度進一步升高時,擴增譜帶明顯變弱,并且穩定性降低。
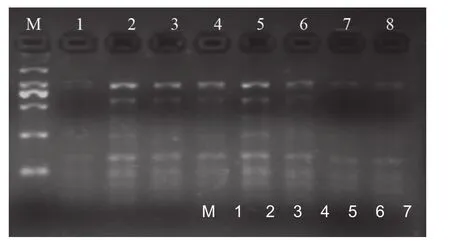
圖3 dNTPs濃度對PCR擴增的影響Fig.3 Effect of dNTPs density on the amplification of PCR
2.2.3 Taq DNA聚合酶用量優化
由圖4可知,當Taq DNA聚合酶為0.4 U時,擴增條帶數明顯少,且模糊不清;當用量為0.6~1.6 U時,譜帶數及亮度基本一致;當Taq DNA聚合酶1.8~2.0 U比1.6 U增加一條譜帶,清晰度有所增加。由于反應體系中 TaqDNA聚合酶的濃度過大時非特異性擴增可能增多,濃度過低又可能導致合成新鏈的效率的下降,最終導致擴增產物的減少,TaqDNA聚合酶的最佳用量為1.2 U即可。
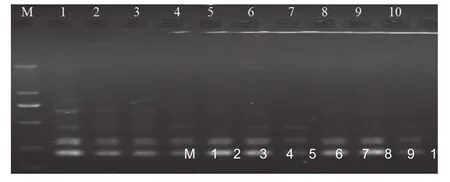
圖4 Taq DNA聚合酶用量對PCR擴增的影響Fig.4 Effect of Taq DNA polymerase on the amplification of PCR
由圖5結果表明:引物濃度的變化對反應體系的影響較大,當引物濃度為800和700 μmol/L時,擴增條帶較少且較弱,當引物濃度降低至100 μmol/L時,獲得條帶數量隨之增加且條帶清晰,增加多為250 bp以下的條帶,擴增效果最好;當引物濃度增加至為900 μmol/L條帶數和亮度也較700 μmol/L時增加,增加的多為1000 bp左右的大片段,但較密集。經比較認為引物的最佳濃度為100 μmol/L。
2.2.4 模板DNA用量
試驗結果表明(圖6),當模板的用量為10~60 ng時,均能擴增出清晰的帶型,隨著模板DNA用量的增加,擴增出譜帶數量基本一致,而譜帶的亮度逐漸增加。模板20 ng時,譜帶的亮度適于各條譜帶的區分,并且能節省模板DNA的用量,確定為最佳模板用量,即20 μL反應體系中的20 ng模板DNA。

圖6 模板DNA用量對PCR擴增的影響Fig.6 Effect of use of template DNA on the amplification ofPCR
2.2.5 SRAP反應體系的正交優化
比較單因素篩選結果,選出正交試驗各因子較好的4個水平,進行正交試驗體系優化。結果如圖7所示,在16個正交組合中,各因素組合不同,擴增結果明顯不同。16個處理均能擴出譜帶,其中組合8的擴增譜帶數量最多,最清晰。因此確定其為最佳反應體系。即模板DNA 20 ng,Mg2+濃度2.0 mmol/L,dNTP濃度200 mmol/L,Taq酶0.8 U,引物濃度140 mmol/L,反應總體積為20 μL。
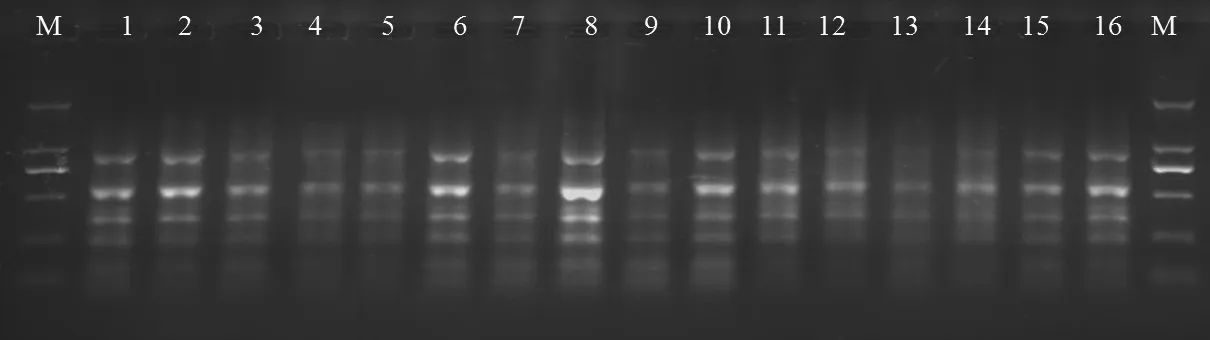
圖7 正交設計擴增的結果Fig.7 Results of amplified orthogonal design
對16個正交組合試驗圖譜進行統計分析,K代表因子在某水平下獲得總條帶數,k代表所對應K值的平均值,R代表k的最大值與最小值之差,對比R值的大小以反映因子對擴增結果的影響,即R越大影響越顯著[15]。由表3可知,Mg2+以2水平的2.0 mmol/L最好,dNTPs 以4水平的200 mmol/L最好,引物則以3水平的140 mmol/L最好,模板DNA用量以4水平的20 ng用量最好,Taq DNA 聚合酶以1水平的0.8 U最好。該數據剛好與通過圖譜觀察最佳組合8的結果一致。進一步對比R大小,反應體系中對擴增結果的影響強弱順序為Taq DNA聚合酶>引物 > Mg2+> dNTPs= 模板 DNA。
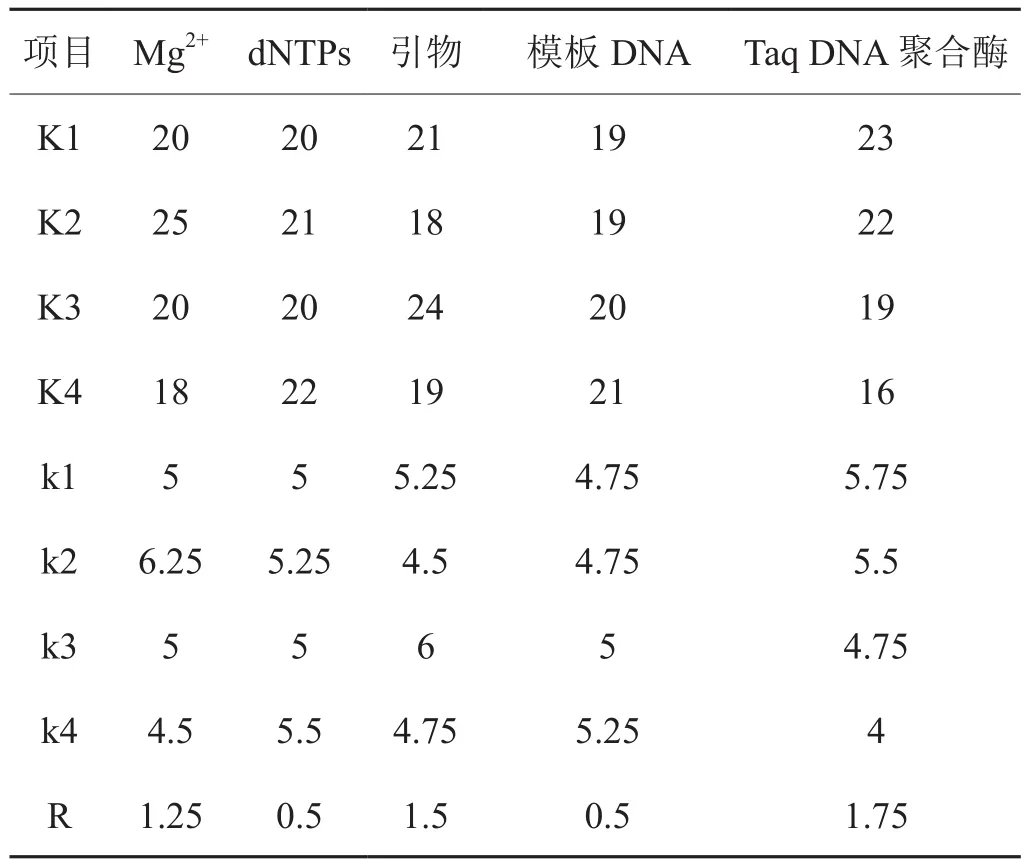
表3 正交試驗結果的統計分析Tab.3 Analytical results of orthogonal experiments
3 結論與討論
在PCR反應體系中各個因素間都會發生相互作用,如dNTPs會與Mg2+發生拮抗作用,Mg2+濃度又會對Taq DNA聚合酶的活性產生直接影響,而Taq DNA聚合酶在引物與模板DNA結合后的延伸中起重要作用[16]。因此,反應體系中無論哪個因素發生改變都會導致擴增結果的變化。采用PCR技術擴增多態性位點及遺傳多樣性研究,易受到這些因素的影響,因此一個適宜的PCR反應體系對于遺傳多樣性研究來說是尤為重要的。目前,PCR體系優化的方法中以單因素法和正交試驗設計應用最為廣泛,其中單因素分析法直觀快速,但是忽略各因素間的互相作用[17],而正交試驗設計較好的避免了這一點,但是正交試驗的各因素水平范圍寬泛,缺乏確定各因素水平的標準。將單因素法與正交試驗設計結合能夠彌補這些缺點。
初步篩選出13對SRAP引物可用于煙草靶斑病菌基因組的多態性分析;并結合了單因素與正交試驗設計對煙草靶斑病菌SRAP-PCR反應體系進化優化,建立了適合該病菌的反應體系:模板DNA 20 ng,Mg2+濃度2.0 mmol/L,dNTP濃度200 mmol/L,Taq酶0.8 U,引物濃度140 mmol/L,反應總體積為20 μL。反應體系中對擴增結果的影響強弱順序為Taq DNA聚合酶>引物> Mg2+> dNTPs=模板DNA。
[1]Li G, Quiros C F.Sequence-related amplified polymorphism (SRAP), a new marker system based on a simple PCR reaction: Its application to mapping and gene tagging in Brassica [J].Theoretical and Applied Genetics, 2001,103(2/3): 455-461.
[2]柳李旺, 龔義勤, 黃浩, 等.新型分子標記-SRAP與TRAP 及其應用[J].遺傳, 2004, 26(5): 777-781.
[3]徐操, 趙寶華.SRAP 分子標記的研究進展及其應用 [J].生命科學儀器, 2009, 7(4): 24-27.
[4]Ruiz J J, García-Martínez S, Picó B, et al.Genetic variability and relationship of closely related Spanish traditional cultivars of tomato as detected by SRAP and SSR markers [J].Journal of the American Society for Horticultural Science, 2005, 130(1): 88-94.
[5]Tang L H, Xiao Y, Li L, et al.Analysis of genetic diversity among Chinese Auricularia auricula cultivars using combined ISSR and SRAP markers [J].Current Microbiology, 2010, 61(2): 132-140.
[6]陳碧云, 胡瓊, Dixelius C, 等.利用 SRAP 分析核盤菌遺傳多樣性 [J].生物多樣性, 2010, 18(5): 509-515.
[7]王輝, 劉長遠, 趙奎華, 等.遼寧省辣椒疫病菌多態性及致病力分化研究初探 [J].微生物學通報, 2012,39(2):180-190.
[8]Costa A S.Mancha aureolada erequeima do fumo causades por corticium solani [J].Biologicol, 1948,14:113-114.
[9]Vargas E.Infection by basidiospores ofThanatephoruscucumeriscausing a foliar disease in tobacco [J].Turrialba,1973, 23:357-359.
[10]Shew H D, Main C E.Rhizoctonialeaf spot of flue-cure tobacco in North Carolina [J].Plant disease, 1985, 69: 901-90.
[11]吳元華, 王左斌, 劉志恒,等.我國煙草新病害—靶斑病[J].中國煙草學報, 2006, 12(6): 22-23.
[12]伏穎, 吳元華, 穆凌霄,等.煙草靶斑病菌基因組DNA提取及RAPD反應體系的優化 [J].煙草科技, 2011, 11:71-75,78.
[13]伏穎, 趙秀香,趙艷琴, 等.煙草靶斑病侵染特性研究 [J].中國煙草學報, 2012, 18(5): 56-59.
[14]吳元華, 趙艷琴, 趙秀香,等.煙草靶斑病原鑒定及生物學特性研究 [J].沈陽農業大學學報,2012,43(5):521-527.
[15]張麗, 周蘭英, 肖千文, 等.正交試驗設計在建立杜鵑花RAPD-PCR反應體系中的應用 [J].北方園藝,2007(5):124-126.
[16]任小巍, 王瑜, 袁慶華.正交設計優化草地早熟禾SRAP-PCR反應體系及引物篩選 [J].草業科學,2012(3):411-416.
[17]張平湖, 劉冠明.橄欖SRAP-PLR體系的建立和優化[J].中國農學通報, 2010,26(15):86-88.
Establishment and optimization of SRAP-PCR reaction system forRhizoctonia solanifrom tobacco target spot
ZHAO Yanqin1,2, WU Yuanhua1, ZHAO Xiuxiang1, AN Mengnan1, CHEN Jianguang1
1 College of Plant Protection, Shenyang Agricultural University, Shenyang 110866, China;
2 College of Agriculture, Inner Mongolia University for the Nationalities, Tongliao, 028043 China
SRAP primer pairs were screened for polymorphism using DNA ofRhizoctonia solaniisolates YC-9, LJT-8 and QYS-7 as templates.An orthogonal design of L16(45)was used to optimize SRAP-PCR reaction system forR.solaniof tobacco with 5 factors, namely Mg2+, dNTPs, primers, Taq DNA polymerase and template DNA.Results showed that a total of 13 polymorphic SRAP primer pairs were screened out of 100 SRAP primer pairs, and a suitable SRAP-PCR reaction system forRhizoctonia solanifrom tobacco target spot was 2.0 mmol.L-1 Mg2+, 200 mmol.L-1 dNTPs, 0.8U Taq DNA polymerase, 140 mmol.L-1 primer pairs, 20 ng template DNA and 1×PCR buffer.In addition, each factor in SRAP-PCR reaction system had different effects on amplified patterns in descending order of Taq DNA polymerase> primer>Mg2+> dNTPs=DNA.
Rhizoctonia solanifrom tobacco target spot; orthogonal experiment design; optimization of reaction system; primer screening
10.3969/j.issn.1004-5708.2014.03.016
S432.41; Q78 文獻標志碼:A 文章編號:1004-5708(2014)03-0096-06
國家煙草專賣局科技攻關項目 [國煙辦綜 (2010)182號];遼寧省煙草專賣局科技攻關項目[遼煙計(2010)86號]
趙艷琴(1978—),博士研究生,植物病原真菌學,Email:zhaoyanqin782828@qq.com
吳元華(1963—),博士,教授,植物病理學及生物農藥,Email:wuyh09@vip.sina.com
2013-04-23
猜你喜歡
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
房地產導刊(2022年5期)2022-06-01 06:20:14
建材發展導向(2021年12期)2021-07-22 08:06:48
建材發展導向(2021年7期)2021-07-16 07:07:52
中學生數理化(高中版.高二數學)(2021年12期)2021-04-26 07:43:48
新世紀智能(數學備考)(2020年11期)2021-01-04 00:38:16
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
當代化工研究(2016年9期)2016-03-20 16:22:08
新高考·高一物理(2014年1期)2014-09-18 01:26:07
聲屏世界(2014年6期)2014-02-28 15:18:09
