鼠傷寒沙門氏菌鞭毛蛋白可增強DnaJ蛋白的免疫反應①
2014-11-27 10:26:36劉宇思胥文春張雪梅
中國免疫學雜志 2014年8期
關鍵詞:小鼠
嚴 明 劉宇思 張 帥 胥文春 王 虹 張雪梅
(重慶醫科大學檢驗醫學院臨床檢驗診斷學教育部重點實驗室,重慶 400016)
佐劑是一種免疫增強劑可以擴大機體對抗原的免疫應答能力,自二十世紀20年代以來,許多物質被作為免疫佐劑進行研究。鋁佐劑是第一個被批準用于人的佐劑,也是目前人類疫苗最常用的佐劑,但其主要誘導體液免疫,抗體亞型以IgG1為主,對細胞免疫的促進能力弱[1]。弗氏佐劑能誘導細胞免疫,但是其具有組織損傷、致癌等毒性反應,不適合作為人類疫苗的組成成分[2]。隨著疫苗種類的不斷豐富,尤其是各種亞單位疫苗、蛋白疫苗的研發,新型佐劑的研究變得越來越重要[3,4]。
鞭毛蛋白是構成細菌的鞭毛纖維的粒狀蛋白質、具有較強的抗原性,它可被TLR5識別,并與其相結合誘導產生固有免疫應答和細胞因子的分泌,鞭毛蛋白作為天然免疫的誘導劑可以激活宿主免疫應答從而顯示出佐劑效應,目前已成為新型佐劑研究的熱點之一[5,6]。
肺炎鏈球菌是感染性疾病的重要病原體,主要威脅兒童和老年人的健康,疫苗是預防其感染的一個主要方式[7]。在本課題組前期研究發現,在CT佐劑存在的情況下,鼻腔免疫肺炎鏈球菌DnaJ蛋白可以抵抗致死劑量的肺炎鏈球菌感染[8]。但CT佐劑具有較強的細胞毒性,不能用于人類疫苗。因此,本研究將鼠傷寒沙門氏菌鞭毛蛋白與肺炎鏈球菌DnaJ蛋白聯合免疫,觀察其免疫效應及對小鼠感染肺炎鏈球菌的保護效果,為DnaJ蛋白疫苗的開發和鞭毛蛋白作為佐劑的應用提供依據。
1 材料與方法
1.1 材料
1.1.1 主要試劑與儀器 DNA純化試劑盒(Roche)、DNA提取試劑盒(Qiangen)、限制性內切酶NdeⅠ和SacⅠ(TaKaRa);T4-DNA連接酶(Progema公司產品);質粒抽提試劑盒(Omega公司產品);IPTG、丙烯酰胺(Sigma產品);咪唑、Tris、NaCl(BBI產品);His-Bind Column(Ni-NTA)柱(Novagen公司產品);HRP標記的山羊抗小鼠IgG或IgA(北京中杉金橋生物技術有限公司);IgG抗體亞型試劑盒(Santa公司);DMEM培養基(Invitrogen公司);細胞因子檢測試劑盒(Biolegend公司);哥倫比亞血平板(重慶龐統醫療器械有限公司);卡巴膽堿(山東博士倫福瑞達制藥有限公司);LB培養基及C+Y培養基由本實驗室配制。GST-谷胱甘肽-S-轉移酶[(是一種溶解性高的多肽,作為目的蛋白表達的載體,增加可溶性)購于Novagen公司]。
1.1.2 菌種與實驗動物 肺炎鏈球菌標準菌株NCTC7466(D39)購買于英國國家菌種保藏管理中心(CMCC);鼠傷寒沙門氏菌標準菌株ATCC14208由重慶醫科大學附屬兒童醫院檢驗科惠贈;E.coli BL21(DE3)菌株、質粒 pET28(a)(本實驗室保存);重組 DnaJ蛋白(本實驗室保存[8]);SPF級C57BL/6小鼠、6~8周、雌性(由重慶醫科大學動物中心提供)。
1.2 方法
1.2.1 傷寒沙門菌鞭毛蛋白基因的擴增、克隆及序列鑒定 ATCC14208于LB培養基中增菌至對數生長期,按照試劑盒說明書提取基因組DNA,擴增鞭毛蛋白的基因編碼區。引物由華大基因公司合成,序列如下:上游引物:GGAATTCCATATGGCACAAGTAATCAAC(下劃線為NdeⅠ酶切位點);下游引物:CGAGCTCTTAACGTAACAGAGACAGCAC(下劃線為SacⅠ酶切位點)。取5 μl擴增產物進行電泳(1%的瓊脂糖凝膠)鑒定后,用DNA純化試劑盒對PCR產物進行純化。
1.2.2 鞭毛蛋白重組質粒的構建及鑒定 純化后的PCR產物和pET28(a)質粒分別用限制性內切酶NdeⅠ、SacⅠ進行雙酶切,并用DNA純化試劑盒回收酶切產物,然后用T4 DNA連接酶16℃連接過夜。連接產物轉化DE3感受態。挑取單個菌落行菌落PCR進行陽性篩選,陽性菌落用試劑盒抽提質粒后進行NdeⅠ、SacⅠ限制性內切酶雙酶切鑒定然后送往華大基因公司進行測序鑒定。
1.2.3 鞭毛蛋白的表達及純化 測序成功后根據本實驗課題組方法挑取陽性克隆接種LB培養液(含 50 μg/ml卡那霉素)37℃ 培養過夜[9,10]。次日將飽和培養物按1∶50接種至含相同卡那霉素濃度的LB培養液中,37℃、180 r/min培養至OD(600)=0.5時加不同濃度IPTG,20℃、120 r/min誘導培養5、8、10 h;4℃離心收集菌體。將菌體用Binding buffer(含 500 mmol/L NaCl、20 mmol/L Tris,pH=8.0)重懸后,進行超聲破菌,然后離心后分別收集上清和沉淀,通過SDS-PAGE電泳分析蛋白可溶性表達情況。收集超聲破菌后離心的上清液轉入經Binding Buffer(成分如前所述)平衡后的Ni-NTA樹脂進行純化。用含不同濃度咪唑的Washing Buffer梯度洗脫雜蛋白后用Elution Buffer(500 mmol/L咪唑,500 mmol/L NaCl,20 mmol/L Tris,pH=8.0)洗脫目的蛋白。純化后目的蛋白經SDS-PAGE鑒定后,用PBS對目的蛋白進行超濾除咪唑,待咪唑濃度超濾至1 mmol/L以下時進行蛋白定量,分裝保存于-80℃備用。
1.2.4 小鼠鼻腔免疫 將C57BL/6小鼠隨機分成三組,分別為鞭毛蛋白組、DnaJ組及DnaJ與鞭毛蛋白混合組。用乙醚吸入麻醉小鼠后分別通過鼻腔滴入30 μl PBS(含10 μg相應蛋白),每周免疫一次,連續免疫三次。
1.2.5 黏膜免疫后小鼠特異性體液免疫及細胞免疫反應的檢測 末次免疫1周后,取小鼠尾靜脈血(每組4只),同時腹腔注射30 μl卡巴膽堿使小鼠產生唾液并收集;分離血清存于-20℃。用DnaJ蛋白(濃度為5 μg/ml)作為抗原包被96孔板小鼠血清作為一抗從1∶1 000開始稀釋到1∶5 120 000,HRP標記的山羊抗小鼠IgG及其個亞型作為二抗,通過間接ELISA的方法測血清總IgG及其亞型的效價。同理檢測唾液中特異性分泌型IgA的效價,具體如本實驗室所做[9]。
末次免疫1周后,分別取三組小鼠各3只,分離獲取脾細胞。重懸于含10%FBS的DMEM培養基,并加至24孔細胞培養板,調整每孔細胞量為5×105個,實驗孔加入5 μg的DnaJ蛋白,陰性對照孔加入同體積的 PBS,37℃,5%CO2,孵育;72 h后取細胞培養的上清液,ELISA法檢測細胞因子IL-17A、IL-4、IL-10、IFN-γ水平,操作按照試劑盒說明書進行。
1.2.6 小鼠的主動保護實驗 在末次免疫兩周后,鼻腔感染5×107CFU的D39菌株,連續監測小鼠生存狀態至21 d,計算生存率。
1.3 統計學分析 用GraphPad Prism 5軟件進行作圖和分析。組間抗體滴度及細胞因子的比較用完全隨機設計資料的兩樣本Student's t檢驗,組間生存率比較用Log-rank(Mantel-Cox)檢驗,以P<0.05為差異有統計學意義。
2 結果
2.1 傷寒沙門鞭毛蛋白基因的擴增 以鼠傷寒沙門氏菌DNA為模板擴增鞭毛蛋白目的基因,進行1%瓊脂糖凝膠電泳分析,可見大小約1 500 bp左右的一條清晰條帶,與預期相符(圖1A)。
2.2 pET28a-Flagellin重組表達質粒的構建 擴增產物與表達載體pET-28a(+)連接轉化DE3感受態,所得陽性克隆經菌落PCR及質粒雙酶切鑒定,1%瓊脂糖電泳出現的條帶與目的片段大小一致(圖1B)。陽性克隆送往公司測序,測序結果與Gen-Bank公布的序列一致,證明重組質粒構建成功。
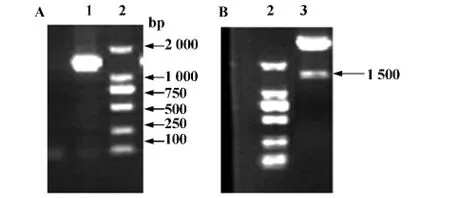
圖1 pET28a-Flagellin重組表達質粒的構建與重組質粒雙酶切鑒定Fig.1 Construction and double enzyme identification of recombinant expression plasmid
2.3 重組鞭毛蛋白的誘導表達與純化 確定pET-28a(+)-Flagellin/DE3的最適合表達條件是:IPTG終濃度為0.1 mmol/L時,20℃、120 r/min誘導8 h、SDS-PAGE結果顯示誘導后全菌和上清均在分子量為52 kD左右出現一粗的蛋白條帶(與預期相符),而未誘導全菌未出現(如圖2);將此上清經Ni-NTA柱純化后,得到單一的目的蛋白,其純度經凝膠掃描可見達95%以上,可用于后續實驗。
2.4 小鼠免疫后產生的特異性抗體及亞型 為排除總蛋白量增加以及重組蛋白殘留內毒素可能帶來的影響,我們采用GST作為無關蛋白對照,設置了單獨DnaJ蛋白組和DnaJ蛋白+GST蛋白組免疫小鼠,分別作為陰性對照1和2。間接ELISA結果顯示(如圖3),陰性對照1和2免疫組小鼠的血清特異性IgG水平無顯著差異,而鞭毛蛋白與DnaJ蛋白聯合免疫組小鼠血清中特異的IgG抗體效價較二者顯著升高(P<0.01)。抗體亞型檢測顯示,鞭毛蛋白與DnaJ蛋白聯合免疫小鼠后,血清中的IgG抗體亞型以IgG1為主,其次為IgG2a和IgG2b,未檢測到IgG3(如圖4)。
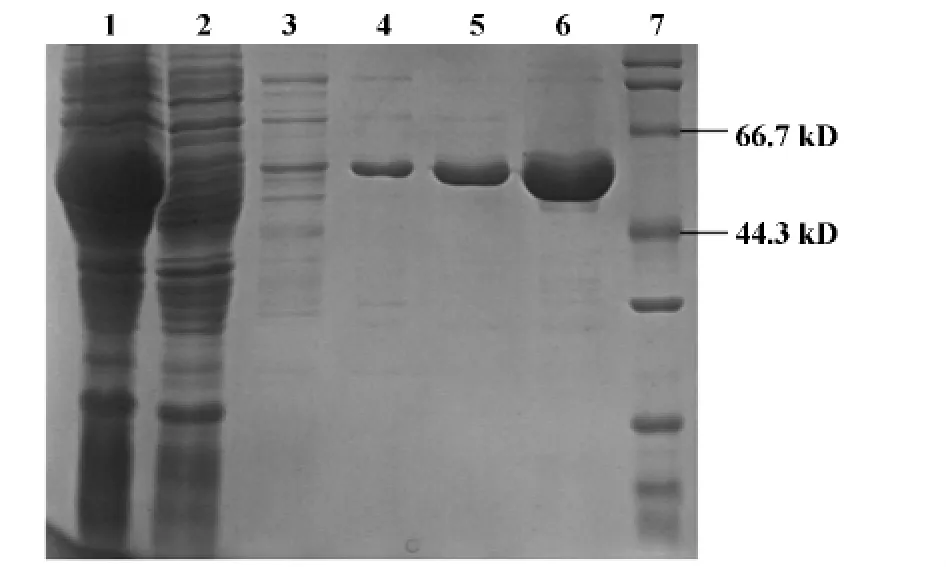
圖2 鞭毛蛋白純化前后SDS-PAGE的電泳結果Fig.2 Electrophoresis results of SDS-PAGE for flagellin
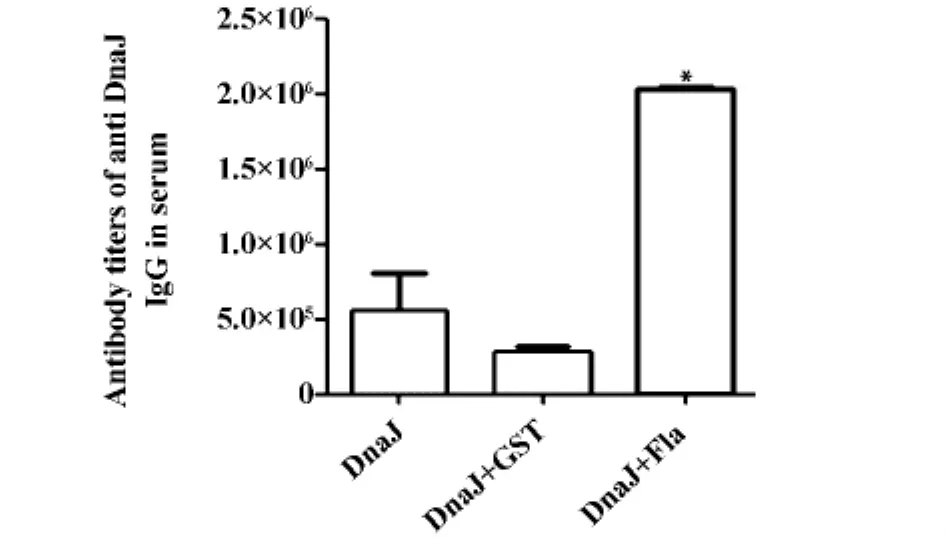
圖3 小鼠血清中特異抗體效價Fig.3 Special serum antibody titers of mouse
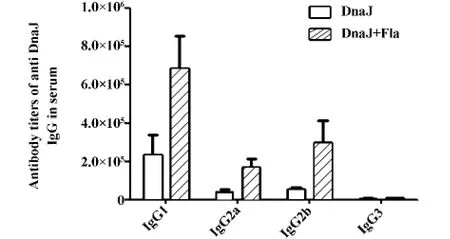
圖4 小鼠血清中抗體亞型效價Fig.4 Serum subtype antibody titers of mouse
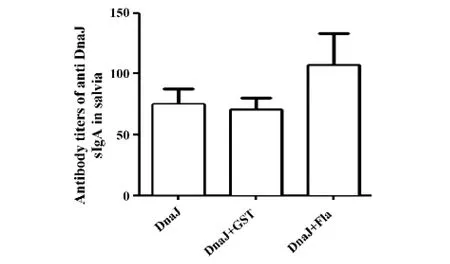
圖5 小鼠唾液中特異性抗體效價Fig.5 Special antibody titers in saliva of mice
唾液IgA抗體水平檢測結果顯示,聯合免疫組小鼠唾液IgA抗體水平較陰性對照組升高,但無統計學差異(如圖5)。
2.5 細胞因子檢測 研究顯示,Th1、Th2和Th17型細胞免疫反應可能參與肺炎鏈球菌疫苗的保護作用[10],本研究檢測了相關的細胞因子:IFN-γ(Th1型)、IL-4、IL-10(Th2型)、IL-17A(Th17 型)。結果如圖6顯示,鞭毛蛋白與DnaJ蛋白聯合免疫組較DnaJ蛋白+GST蛋白免疫組和單獨DnaJ蛋白組產生了更高水平的 IFN-γ(P<0.01)、IL-17A(P<0.01)和IL-4(P<0.01),提示鞭毛蛋白可以增強DnaJ誘導的特異性Th1、Th2和Th17型細胞免疫應答。
2.6 小鼠肺炎鏈球菌D39攻毒保護試驗 末次免疫兩周后對三組小鼠鼻腔滴注致死劑量的肺炎鏈球菌D39(5×107CFU),連續觀察21 d小鼠的保護效果。結果顯示,鞭毛蛋白與DnaJ混合組的生存率為60%,與單獨鞭毛蛋白組相比有統計學差異(P<0.01),也高于單獨 DnaJ蛋白免疫組的生存率(50%),提示鞭毛蛋白可以增強DnaJ蛋白誘導的保護性免疫效應(如圖7)。
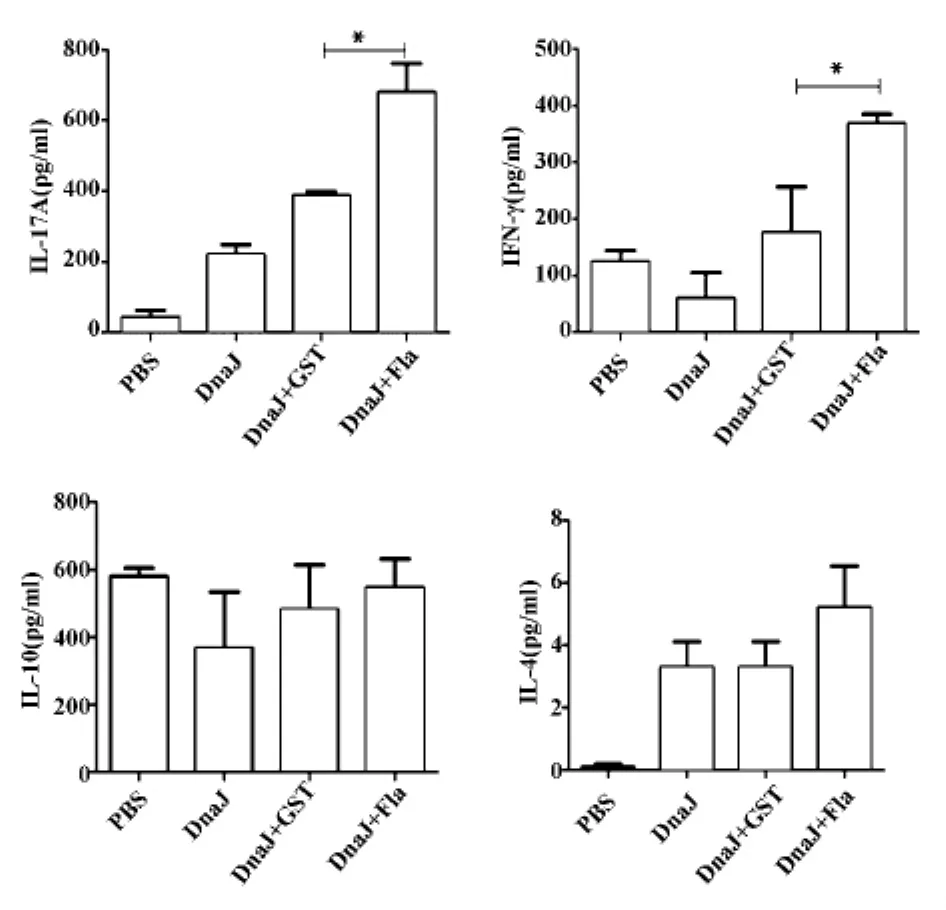
圖6 免疫后小鼠脾細胞產生的細胞因子Fig.6 Cytokine levels in spleen cells of immunized mice
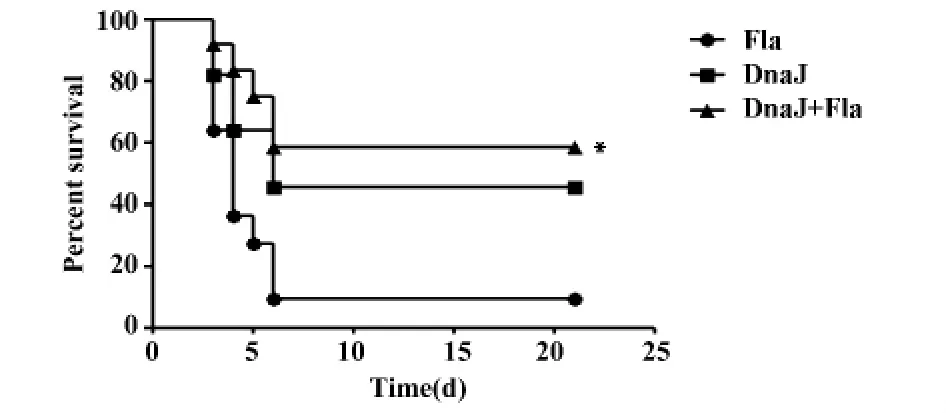
圖7 小鼠感染肺炎鏈球菌D39的生存率Fig.7 Survival rate against D39 infection
3 討論
肺炎鏈球菌是一種嚴重危害人們健康的病原體,而疫苗接種是預防其感染的一個有效方法[11]。隨著人們對不同類型的疫苗的研究,原有的佐劑也顯示出其局限性,因此新型佐劑的研究也變得十分迫切[12]。鞭毛蛋白是傷寒沙門氏菌細菌鞭毛的主要蛋白,是目前已知的TLR5唯一激動劑,可以通過TLR5途徑激活宿主免疫應答,研究表明鞭毛蛋白與抗原以不同方式融合后可以發揮佐劑作用[13]。但是融合蛋白可能對鞭毛蛋白本來的構象產生影響,本研究主要將鞭毛蛋白與DnaJ蛋白聯合免疫小鼠,通過研究其與DnaJ蛋白聯合免疫對肺炎鏈球菌感染的保護作用來對鞭毛蛋白佐劑效應進行評價。
本研究成功表達與純化鞭毛蛋白,并將其與DnaJ蛋白聯合免疫C57BL/6小鼠(設置不同條件的對照組),所有小鼠免疫3次后取鼠尾靜脈血和唾液。ELISA結果表明鞭毛蛋白與DnaJ蛋白聯合免疫可以誘導宿主產生較高劑量的IgG抗體,且IgG抗體亞型以IgG1為主同時產生一定量的IgG2a和IgG2b,說明鞭毛蛋白與DnaJ蛋白混合免疫時鞭毛蛋白有增強DnaJ蛋白免疫效應的作用。
脂多糖(Lipopolysaccharide,LPS)是大腸桿菌細菌內毒素的主要毒性成分,具有致熱作用。通過大腸桿菌表達外源蛋白時,破菌收集蛋白時會混入大量的LPS,后續的親和層析純化步驟不能保證完全去除溶液中混有的LPS。有報道提示,LPS可以刺激機體免疫反應,具有黏膜佐劑的能力[14]。因此,本實驗采用了GST蛋白作為無關蛋白對照,以排除LPS可能產生的干擾作用[15]。該對照同時也排除了蛋白量增多而產生的影響。抗體效價檢測結果顯示,DnaJ+GST對照組誘導的特異性IgG水平與單獨DnaJ蛋白組無顯著差別,而DnaJ+鞭毛蛋白組的特異性IgG效價較二者顯著增高,這表明鞭毛蛋白確實能夠增強DnaJ的免疫原性,具有佐劑效應。
為了進一步研究其免疫增強效應的類型,我們對不同免疫組的脾細胞分泌的細胞因子水平進行檢測,結果發現與PBS對照組相比,鞭毛蛋白與DnaJ蛋白聯合與單獨DnaJ蛋白免疫組小鼠均檢測到了顯著升高的IL-4、IFN-γ和IL-17A;而聯合免疫組較單獨免疫組的細胞因子水平顯著增高,提示鞭毛蛋白發揮佐劑效應可增強宿主的Th1、Th2及Th17免疫應答。主動性保護實驗提示,鞭毛蛋白與DnaJ蛋白聯合免疫小鼠的生存率(60%)比DnaJ蛋白免疫組(50%)小鼠高,與單一的鞭毛蛋白免疫組相比存在統計學差異。實驗表明鞭毛蛋白可以增強DnaJ蛋白誘導的保護性免疫反應,提高宿主肺炎鏈球菌感染抵抗能力。
綜上所述,我們的研究結果表明:傷寒沙門氏菌的表面鞭毛蛋白具有佐劑作用,與DnaJ蛋白聯合免疫可以增強其誘導的保護性免疫反應并提高宿主抵抗肺炎鏈球菌感染的能力。本研究為鞭毛蛋白作為蛋白疫苗佐劑的使用提供實驗依據。
[1]Jazani NH,Sohrabpour M,Mazloomi E,et al.A novel adjuvant,a mixture of alum and the general opioid antagonist naloxone,elicits both humoral and cellular immune responses for heat-killed Salmonella typhimurium vaccine[J].FEMS Immunol Med Microbiol,2011,61(1):54-62.
[2]Batista-Duharte A,Lindblad EB,Oviedo-Orta E.Progress in understanding adjuvant immunotoxicity mechanisms[J].Toxicol Lett,2011,203(2):97-105.
[3]Blaas SH,Stieber-Gunckel M,Falk W,et al.CpG-oligodeoxy nucleotides stimulate immunoglobulin A secretion in intestinal mucosal B cells[J].Clin Exp Immunol,2009,155:534-540.
[4]Brito LA,Malyala P,O'Hagan DT.Vaccine adjuvant formulations:A pharmaceutical perspective[J].Seminars in Immunology,2013,25(2):130-145.
[5]Pino O,Martin M,Michalek SM.Cellular mechanisms of the adjuvant activity of the flagellin component FljB of Salmonella enterica serovar Typhimurium to potentiate mucosal and systemic responses[J].Infect Immun,2005,73(10):6763-6770.
[6]Atif SM,Uematsu S,Akira S,et al.CD103-CD11b+dendritic cells regulate the sensitivity of CD4 T-cell responses to bacterial flagellin[J].Mucosal Immunol,2014,7(1):68-77.
[7]Gentile A,Bazán V.Prevention of pneumococcal disease through vaccination[J].Vaccine,2011,29:C15-C25.
[8]Cui Y,Zhang X,Gong Y,et al.Immunization with DnaJ(hsp40)could elicit protection against nasopharyngeal colonization and invasive infection caused by different strains of Streptococcus pneumoniae[J].Vaccine,2011,29(9):1736-1744.
[9]Gong Y,Xu W,Cui Y,et al.Immunization with a ZmpB-based protein vaccine could protect against pneumococcal diseases in mice[J].Infect Immunol,2011,79(2):867-878.
[10]Kadioglu A,Gingles NA,Grattan K,et al.Host cellular immune response to pneumococcal lung infection in mice[J].Infect Immunol,2000,68(2):492-501.
[11]Fasche S,Van Hoek AJ,Sheasby E,et al.Effect of pneumococcal conjugate vaccination on serotype-specific carriage and invasive disease in England:a cross-sectional study [J].Plos Med,2011,8(4):e1001017
[12]Reed SG,Bertholet S,Coler RN,et al.New horizons in adjuvants for vaccine development[J].Trends Immunol,2009,30(1):23-32.
[13]Rumyantsev AA,Zhang ZX,Gao QS,et al.Direct random insertion of an influenza virus immunologic determinant into the NS1 glycoprotein of a vaccine flavivirus[J].Virology,2010,396:329-338.
[14]Lambrecht BN,Kool M,Willart MA et al.Mechanism of action of clinically approved adjuvants[J].Curr Opin Immunol,2009,21:23-29.
[15]Lee SE,Kim SY,Jeong BC,et al.A bacterial flagellin,Vibrio vulnificus FlaB,has a strong mucosal adjuvant activity to induce protective immunity [J].Infect Immunol,2006,74(1):694-702.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
