nPKC-PKD3信號(hào)傳導(dǎo)途徑介導(dǎo)Prostratin活化HIV-1轉(zhuǎn)錄
2014-12-01 08:15:58王會(huì)平劉潤忠
關(guān)鍵詞:信號(hào)
王會(huì)平,劉 敏,劉潤忠
(廈門大學(xué)生命科學(xué)學(xué)院,福建 廈門361102)
艾滋病在全球范圍內(nèi)流行多年,目前行之有效的治療方案稱之為高效抗逆轉(zhuǎn)錄病毒療法(HAART),即通常為大眾所熟知的“雞尾酒療法”[1-2].“雞尾酒療法”主要針對艾滋病毒的逆轉(zhuǎn)錄酶、蛋白酶以及外殼糖蛋白Gp41,因此它僅對患者體內(nèi)處于復(fù)制周期的活病毒療效顯著,用藥后患者血液中活病毒的滴度可降至無法檢測到的水平,無法徹底清除患者體內(nèi)的艾滋病毒;停藥后,患者的病情極易出現(xiàn)反復(fù)[2-3].近年來人們提出一種新的治療策略[4],用另外一類化合物刺激潛伏的艾滋病毒使其進(jìn)入復(fù)制周期,再聯(lián)用“雞尾酒療法”清除這些活病毒,以此來避免患者終生用藥的負(fù)擔(dān)并最終治愈艾滋病.雖然這種治療策略理論上存在可行性,但目前人們對于艾滋病毒潛伏和重新活化的理解還很有限,缺乏相應(yīng)的分子機(jī)制模型用于篩選這類活化潛伏病毒的化合物,從而造成臨床治療實(shí)踐的進(jìn)展緩慢[5-6].因此,深入探討艾滋病毒的潛伏以及重新活化的分子機(jī)制已經(jīng)成為當(dāng)下研究人員所關(guān)注的焦點(diǎn).
在業(yè)已發(fā)現(xiàn)的、為數(shù)不多的能激活潛伏艾滋病毒的化合物中,Prostratin是研究最多的一種,它作為潛在的抗艾滋病藥物已進(jìn)入II期臨床試驗(yàn)[6-10].Prostratin是不致癌的佛波酯類化合物,具備此類化合物的共性,即活化細(xì)胞內(nèi)蛋白激酶C(PKC)信號(hào)通路[11];該信號(hào)通路的核心蛋白PKC也被證實(shí)對活化免疫T細(xì)胞中潛伏的艾滋病毒起關(guān)鍵性的作用[9].PKC家族包含3個(gè)亞型:典型PKC(tPKC,包括α、βI、βII和γ)、新型PKC(nPKC,包括δ、ε、η和θ)以及非典型PKC(aPKC,包括ζ和λ).PKC家族所有成員的碳端都有相同的具催化活性的結(jié)構(gòu)域;氮端的差異則較大,便于結(jié)合不同的上游調(diào)控蛋白和下游效應(yīng)蛋白從而執(zhí)行不同的功能.當(dāng)胞外信號(hào)刺激細(xì)胞時(shí),胞內(nèi)的磷脂酶C(PLC)首先活化并誘導(dǎo)產(chǎn)生第二信使——甘油二酯(DAG),隨后PKC活化并將信號(hào)傳遞給下游效應(yīng)分子諸如蛋白激酶D(PKD)以執(zhí)行相應(yīng)的生物學(xué)功能[12-13].由于基因的轉(zhuǎn)錄活化是激活潛伏艾滋病毒的前提,因此Prostratin是否也是通過活化PKC信號(hào)途徑激活艾滋病毒的基因轉(zhuǎn)錄,進(jìn)而活化潛伏的艾滋病毒成為亟待解決的問題.
本文選用穩(wěn)定表達(dá)HIV-LTR-Luciferase報(bào)告基因的 HeLa細(xì)胞(HIV-LTR-Luc細(xì)胞)為研究對象,用Prostratin和多種PKC抑制劑聯(lián)合處理細(xì)胞,證實(shí)了nPKC亞型參與Prostratin誘導(dǎo)的HIV-1轉(zhuǎn)錄活化,并最終通過效應(yīng)蛋白PKD3激活HIV-1轉(zhuǎn)錄的分子信號(hào)傳導(dǎo)機(jī)制.
1 材料與方法
1.1 材 料
PKC抑制劑 G?6976、G?6983和BIMI(G?6850)購自美國Calbiochem公司;Prostratin購自美國LC Laboratories公司;轉(zhuǎn)染試劑PEI購自美國Polysciences公司;熒光素酶底物購自美國Promega公司.持續(xù)活化型PKCθ、PKCε和PKCα的表達(dá)質(zhì)粒受贈(zèng)于美國華盛頓大學(xué) Andrew Matthews Scharenberg教授[14-15];PKD3敲 低 質(zhì) 粒 (shPKD3)的 靶 序 列 為:5′-GTC CTA AGA CGG GAC TCT C[16].
1.2 方 法
1.2.1 細(xì)胞培養(yǎng)和藥物處理實(shí)驗(yàn)
HIV-LTR-Luc穩(wěn)定細(xì)胞株[17]培養(yǎng)在含10%(質(zhì)量分?jǐn)?shù))胎牛血清(FBS)的DMEM完全培養(yǎng)基中.藥物處理時(shí)細(xì)胞密度約為50%~70%,先用PKC抑制劑預(yù)處理1h,然后用2μmol/L的Prostratin繼續(xù)處理6h.藥物處理后,收集并裂解細(xì)胞用于熒光素酶活性分析.
1.2.2 熒光素酶活性分析
將HIV-LTR-Luc穩(wěn)定細(xì)胞株培養(yǎng)在六孔板里并用相應(yīng)的藥物處理.藥物處理后,細(xì)胞裂解液按照以前描述過的方案測量熒光素酶活性[18],根據(jù)3次不同的實(shí)驗(yàn)計(jì)算標(biāo)準(zhǔn)誤差.
1.2.3 細(xì)胞瞬時(shí)轉(zhuǎn)染
轉(zhuǎn)染前1d將HIV-LTR-Luc穩(wěn)定細(xì)胞株胞培養(yǎng)在六孔板里,細(xì)胞密度約為70%~90%.將2μg待轉(zhuǎn)染的質(zhì)粒和3μL PEI溶液混勻后加入細(xì)胞培養(yǎng)基中,繼續(xù)培養(yǎng)48h.細(xì)胞裂解液按照以前描述過的方案測量熒光素酶活性[18],根據(jù)3次不同的實(shí)驗(yàn)計(jì)算標(biāo)準(zhǔn)誤差.
2 實(shí)驗(yàn)結(jié)果
2.1 Prostratin刺激HIV-LTR-Luc的轉(zhuǎn)錄活化
為研究Prostratin激活HIV-1的轉(zhuǎn)錄活性進(jìn)而活化潛伏的艾滋病毒的分子機(jī)制,本文選用了HIVLTR-Luc穩(wěn)定細(xì)胞株.該細(xì)胞株構(gòu)建自HeLa細(xì)胞,它整合了由HIV-1長末端(LTR)啟動(dòng)子所調(diào)控的熒光素酶報(bào)告基因,其熒光素酶的表達(dá)水平直接反映HIV-1的轉(zhuǎn)錄活性[17-18].
首先,用不同濃度的Prostratin處理細(xì)胞6h.如圖1所示,熒光素酶的表達(dá)水平隨Prostratin濃度上升而升高,即 HIV-1的轉(zhuǎn)錄活性也在不斷升高;當(dāng)Prostratin濃度為5μmol/L時(shí),HIV-1轉(zhuǎn)錄水平提高100倍以上.為避免高劑量藥物衍生的毒理作用,后續(xù)實(shí)驗(yàn)中Prostratin的處理濃度限定為2μmol/L.緊接著,在這一濃度條件下,測試了不同處理時(shí)間對HIV-1轉(zhuǎn)錄活性的影響.發(fā)現(xiàn)隨著處理時(shí)間的延長,HIV-1轉(zhuǎn)錄活性逐漸增強(qiáng);用Prostratin處理細(xì)胞6h左右已經(jīng)能高效地激活HIV-1的轉(zhuǎn)錄(圖2).由于細(xì)胞在轉(zhuǎn)錄水平應(yīng)答外界信號(hào)刺激可在較短的時(shí)間內(nèi)實(shí)現(xiàn),而長時(shí)間藥物處理會(huì)帶來信號(hào)傳導(dǎo)過程中的次級(jí)應(yīng)答使得問題變得過于復(fù)雜,因此后續(xù)實(shí)驗(yàn)中藥物處理時(shí)間限定為6h.
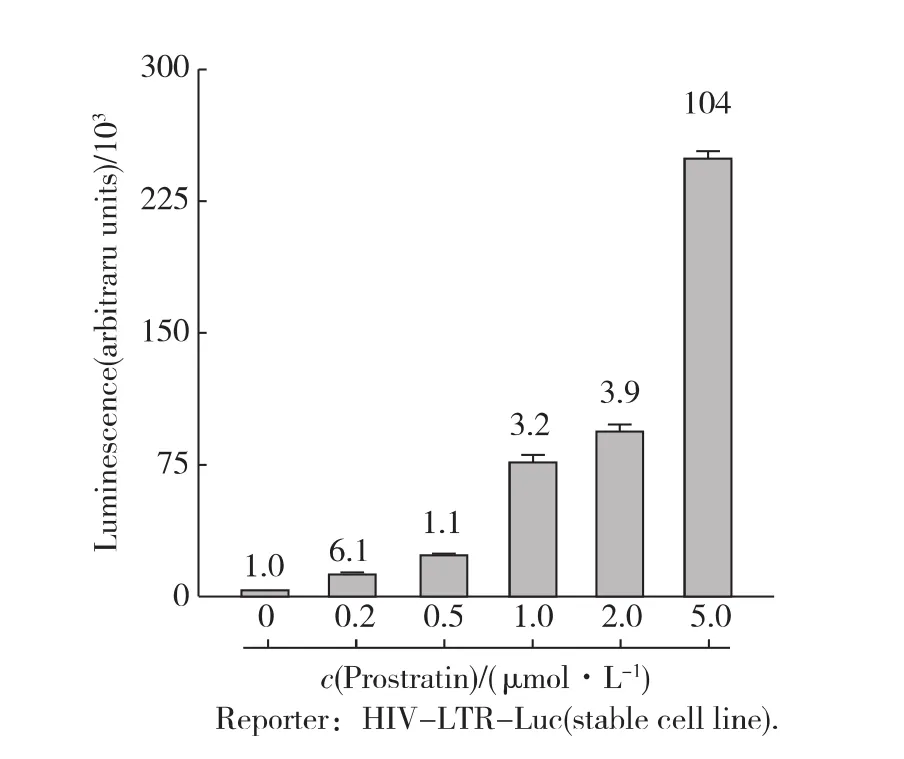
圖1 不同濃度的Prostratin處理后HIV-1的轉(zhuǎn)錄活性Fig.1 Prostratin activated gene expression from stably integrated HIV-LTR-luciferase reporter in a dose-dependent manner
2.2 Prostratin通過nPKC活化HIV-1的轉(zhuǎn)錄
此前其他實(shí)驗(yàn)室在Jurkat免疫T細(xì)胞中證實(shí)蛋白PKC參與Prostratin活化潛伏艾滋病毒的過程[9],那么Prostratin激活HIV-1轉(zhuǎn)錄是否有蛋白PKC的參與呢?首先,我們采用3種不同的蛋白PKC的抑制劑預(yù)處理細(xì)胞再偶聯(lián)Prostratin刺激來觀察HIV-1的轉(zhuǎn)錄活性.如圖3所示,廣譜的PKC抑制劑G?6983和BIM I均能削弱Prostratin刺激引起的HIV-1的轉(zhuǎn)錄活化,而另1種PKC抑制劑G?6976卻沒有這個(gè)作用.仔細(xì)分析這3種PKC抑制劑的特異性,發(fā)現(xiàn)G?6983和 BIM I主 要 抑 制 tPKC 和 nPKC[19],而G?6976只能抑制tPKC;這意味著nPKC可能參與Prostratin刺激的HIV-1的轉(zhuǎn)錄活化.接下來為進(jìn)一步證實(shí)這個(gè)推論,在HIV-LTR-Luc細(xì)胞中瞬時(shí)過表達(dá)了持續(xù)活化型的PKCθ、PKCε和PKCα.如圖4所示,其中2種nPKC均能直接激活HIV-1的轉(zhuǎn)錄活性,而tPKC則無此功能.因此,nPKC直接介導(dǎo)了Prostratin刺激引起的HIV-1的轉(zhuǎn)錄活化.
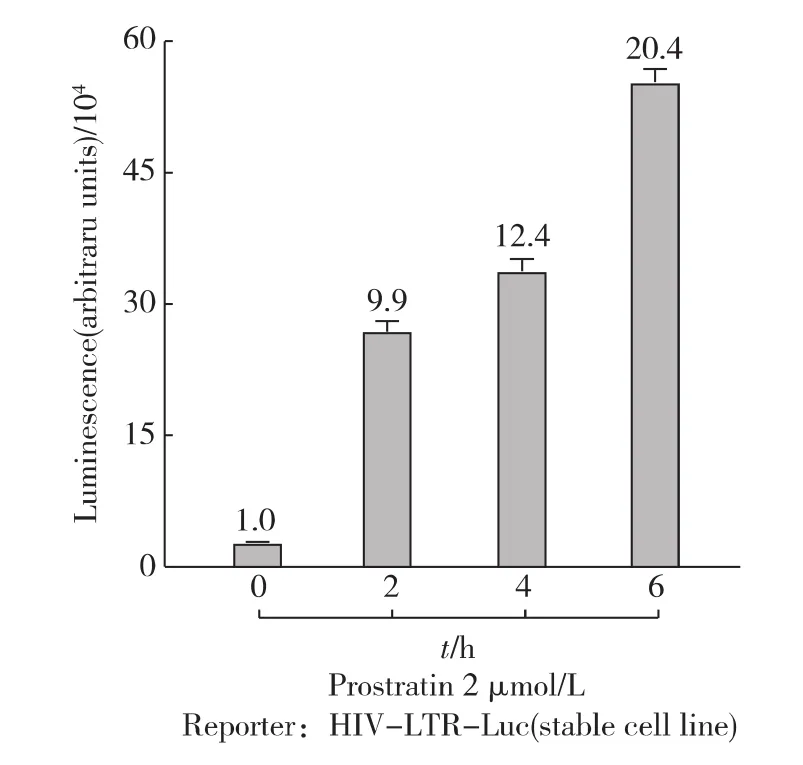
圖2 Prostratin處理不同時(shí)間后HIV-1的轉(zhuǎn)錄活性Fig.2 Time course of Prostratin-stimulated HIV-LTR-Luc expression

圖3 PKC抑制劑阻斷Prostratin活化的HIV-1轉(zhuǎn)錄Fig.3 Effect of PKC inhibitors on Prostratin-activated HIV-1expression
2.3 敲低效應(yīng)蛋白PKD3拮抗Prostratin刺激活化的HIV-1轉(zhuǎn)錄
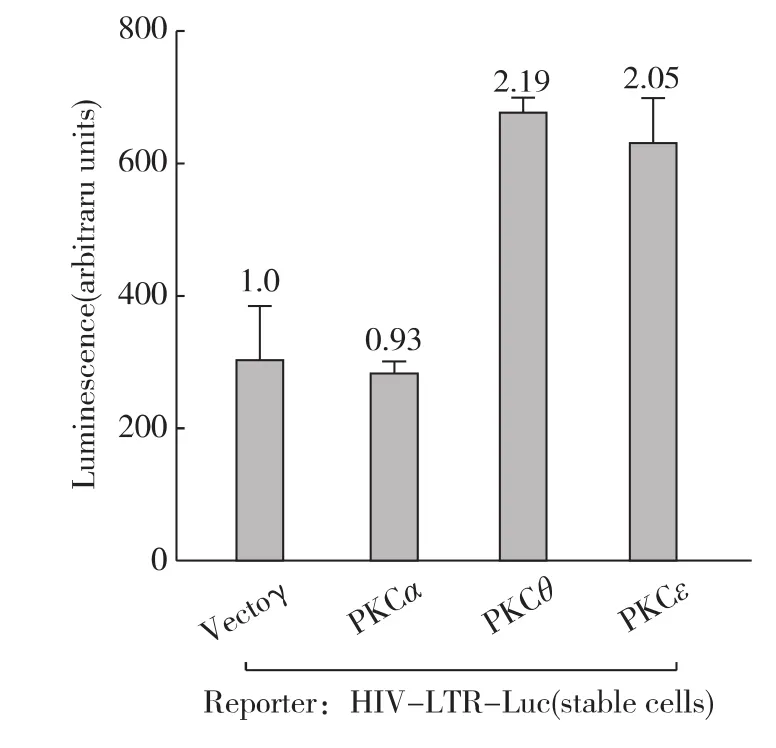
圖4 3種PKC對HIV-1轉(zhuǎn)錄活性的影響Fig.4 Effect of overexpression of selected PKC isoforms on HIV-1expression
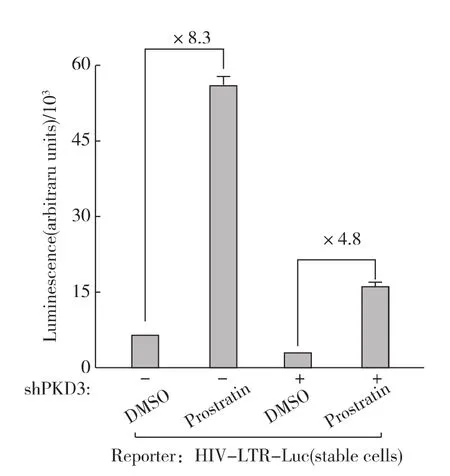
圖5 敲低PKD3削弱Prostratin活化的HIV-1轉(zhuǎn)錄Fig.5 The effect of shRNA knock-down on Prostratin-activated HIV-1expression
下游效應(yīng)蛋白是信號(hào)傳導(dǎo)過程中特定功能的執(zhí)行者.我們之前的實(shí)驗(yàn)證實(shí),PKC信號(hào)途徑的效應(yīng)蛋白PKD最終影響HIV-1的轉(zhuǎn)錄活性[16].PKD家族共有3個(gè)成員,前期的實(shí)驗(yàn)表明PKD3而非PKD1、PKD2介導(dǎo) HIV-1的轉(zhuǎn)錄活化[16].因此,我們先用特異的shRNA敲低PKD3,然后再用Prostratin處理細(xì)胞觀察HIV-1的轉(zhuǎn)錄活性.PKD3被敲低以后(內(nèi)源性PKD3減少約50%)[16],Prostratin刺激所引起的HIV-1轉(zhuǎn)錄活化倍數(shù)也相應(yīng)的減少一半左右(圖5).這進(jìn)一步證明,Prostratin處理激活了nPKC-PKD3信號(hào)傳導(dǎo)途徑從而活化HIV-1的轉(zhuǎn)錄.
3 討 論
盡管Prostratin在臨床上的個(gè)別應(yīng)用已取得了一定的療效[20-21],但具體的藥用機(jī)理仍不清楚,這顯著延緩了臨床廣泛的使用[6].本文從分析HIV-1轉(zhuǎn)錄活性入手,以HIV證實(shí)nPKC-PKD3信號(hào)傳導(dǎo)途徑參與Prostratin刺激引起的HIV-1轉(zhuǎn)錄活性增強(qiáng).本文所發(fā)現(xiàn)的信號(hào)傳導(dǎo)機(jī)制,為提速Prostratin大范圍應(yīng)用于臨床治療提供了可靠的基礎(chǔ)理論依據(jù),同時(shí)也為日后構(gòu)建新的抗艾滋病藥物篩選模型以及藥物作用靶點(diǎn)提供了新的線索.
在本研究前,研究人員以攜帶潛伏HIV-1的免疫細(xì)胞為模型,初步將蛋白PKC與活化潛伏的艾滋病毒的現(xiàn)象相聯(lián)系[9].然而,事實(shí)上該潛伏HIV-1細(xì)胞模型與真實(shí)的艾滋病毒潛伏過程存在明顯的區(qū)別[22].首先,真實(shí)的艾滋病毒潛伏過程中,93%的HIV-1基因組會(huì)整合在宿主處于高度轉(zhuǎn)錄活化狀態(tài)的基因的內(nèi)含子區(qū),因此可能是轉(zhuǎn)錄過程中相伴的RNA剪輯導(dǎo)致RNA聚合酶II直接滑過HIV-1的基因從而引起 HIV-1基因未被轉(zhuǎn)錄[23];而潛伏 HIV-1細(xì)胞模型中,HIV-1基因更多整合在異染色體區(qū),由于染色體空間結(jié)構(gòu)未被打開,該區(qū)的基因處于轉(zhuǎn)錄抑制狀態(tài)[24].其次,真實(shí)的艾滋病毒潛伏多發(fā)生在免疫T細(xì)胞記憶細(xì)胞中,此類細(xì)胞處于細(xì)胞周期的靜息期,不生長也不分裂[25-26];而潛伏 HIV-1細(xì)胞模型為常規(guī)的免疫T細(xì)胞,處于正常的細(xì)胞增殖周期,細(xì)胞快速生長并進(jìn)行有絲分裂[24].因此,為規(guī)避細(xì)胞模型與真實(shí)過程之間的差異,同時(shí)將復(fù)雜的多因素協(xié)同實(shí)現(xiàn)的艾滋病毒潛伏過程簡化為簡單的實(shí)驗(yàn)?zāi)P停x取了專門為研究HIV-1轉(zhuǎn)錄活性而設(shè)計(jì)的 HIV-LTR-Luc細(xì)胞模型.因?yàn)闈摲麳IV-1活化的源頭就是潛伏的HIV-1退出靜息狀態(tài),開始第1輪HIV-1基因的轉(zhuǎn)錄.當(dāng)最初的轉(zhuǎn)錄以及翻譯完成后,新生的病毒蛋白如Tat會(huì)強(qiáng)烈刺激后續(xù)的轉(zhuǎn)錄過程,從而最終促使艾滋病迅猛復(fù)發(fā).
在過去10余年的研究中,我們采用HIV-LTRLuc細(xì)胞模型已經(jīng)成功詮釋過處于復(fù)制周期中的活病毒的轉(zhuǎn)錄機(jī)制,即憑借病毒蛋白Tat綁架宿主轉(zhuǎn)錄系統(tǒng)中的核心分子正性轉(zhuǎn)錄延伸因子b(P-TEFb),并進(jìn)一步利用P-TEFb能刺激基因轉(zhuǎn)錄延伸的性能,方便HIV-1可以順利合成完整的全基因產(chǎn)物[27-29].對于潛伏的艾滋病毒而言,由于第1輪轉(zhuǎn)錄都沒有啟動(dòng),活病毒轉(zhuǎn)錄所倚重的病毒蛋白Tat還沒有產(chǎn)生,潛伏的HIV-1只能更多依賴于轉(zhuǎn)錄因子 NFκB[9].轉(zhuǎn)錄因子NFκB會(huì)募集Brd4-P-TEFb蛋白復(fù)合體(由蛋白Brd4與P-TEFb相互作用形成的蛋白復(fù)合體)到HIV-1啟動(dòng)子區(qū)的NFκB結(jié)合位點(diǎn)附近,通過P-TEFb激活RNA聚合酶II完成第1輪HIV-1基因的轉(zhuǎn)錄.有報(bào)道指出PKD可以調(diào)控NFκB.活化的PKD通過磷酸化激活I(lǐng)KK復(fù)合體,活化了的IKK會(huì)促進(jìn)NFκB抑制蛋白IκB的泛素化降解,從而通過去抑制作用激活轉(zhuǎn)錄因子NFκB.因此當(dāng)Prostratin刺激所產(chǎn)生的應(yīng)答信號(hào)傳遞到效應(yīng)蛋白PKD3以后,PKD3作為PKC信號(hào)傳導(dǎo)途徑與NFκB信號(hào)傳導(dǎo)途徑的整合點(diǎn),最終改變了潛伏HIV-1的轉(zhuǎn)錄活性.
[1]Lewin S R,Evans V A,Elliott J H,et al.Finding a cure for HIV:will it ever be achievable[J].J Int AIDS Soc,2011,14:4.
[2]Stevens M,De Clercq E,Balzarini J.The regulation of HIV-1transcription:molecular targets for chemotherapeutic intervention[J].Med Res Rev,2006,26(5):595-625.
[3]Siliciano R F.Scientific rationale for antiretroviral therapy in 2005:viral reservoirs and resistance evolution[J].Top HIV Med,2005,13(3):96-100.
[4]Archin N M,Margolis D M.Emerging strategies to deplete the HIV reservoir[J].Curr Opin Infect Dis,2014,27(1):29-35.
[5]Han Y,Wind-Rotolo M,Yang H C,et al.Experimental approaches to the study of HIV-1latency[J].Nat Rev Microbiol,2007,5(2):95-106.
[6]Liu M,Hsu J,Chan C,et al.The ubiquitin ligase Siah1 controls ELL2stability and formation of super elongation complexes to modulate gene transcription[J].Mol Cell,2012,46(3):325-334.
[7]Korin Y D,Brooks D G,Brown S,et al.Effects of prostratin on T-cell activation and human immunodeficiency virus latency[J].J Virol,2002,76(16):8118-8123.
[8]Kulkosky J,Culnan D M,Roman J,et al.Prostratin:activation of latent HIV-1expression suggests a potential inductive adjuvant therapy for HAART[J].Blood,2001,98(10):3006-3015.
[9]Williams S A,Chen L F,Kwon H,et al.Prostratin antagonizes HIV latency by activating NF-kappaB[J].J Biol Chem,2004,279(40):42008-42017.
[10]Beans E J,F(xiàn)ournogerakis D,Gauntlett C,et al.Highly potent,synthetically accessible prostratin analogs induce latent HIV expressionin vitro and ex vivo[J].Proc Natl Acad Sci USA,2013,110(29):11698-11703.
[11]Gulakowski R J,McMahon J B,Buckheit R W,Jr,et al.Antireplicative and anticytopathic activities of prostratin,a non-tumor-promoting phorbol ester,against human immunodeficiency virus(HIV)[J].Antiviral Res,1997,33(2):87-97.
[12]Steinberg S F.Structural basis of protein kinase C isoform function[J].Physiol Rev,2008,88(4):1341-1378.
[13]Zugaza J L,Sinnett-Smith J,Van Lint J,et al.Protein kinase D (PKD)activation in intact cells through a protein kinase C-dependent signal transduction pathway[J].EMBO J,1996,15(22):6220-6230.
[14]Matthews S A,Dayalu R,Thompson L J,et al.Regulation of protein kinase Cnu by the B-cell antigen receptor[J].J Biol Chem,2003,278(11):9086-9091.
[15]Matthews S A,Liu P,Spitaler M,et al.Essential role for protein kinase D family kinases in the regulation of class II histone deacetylases in B lymphocytes[J].Mol Cell Biol,2006,26(4):1569-1577.
[16]Zhu Y,Ding F,Juventus A J,et al.PKD3is required for prostratin-activated HIV-1transcription,in IT in Medicine and Education (ITME)[C]∥IT in Medicine and Education (ITME),2011International Symposium on[S.l.]:IEEE,2011:606-610.
[17]Chen R,Liu M,Li H,et al.PP2Band PP1alpha cooperatively disrupt 7SK snRNP to release P-TEFb for transcription in response to Ca2+signaling[J].Genes Dev,2008,22(10):1356-1368.
[18]Yang Z,Zhu Q,Luo K,et al.The 7SK small nuclear RNA inhibits the CDK9/cyclin T1kinase to control transcription[J].Nature,2001,414(6861):317-322.
[19]Way K J,Chou E,King G L.Identification of PKC-isoformspecific biological actions using pharmacological approaches[J].Trends Pharmacol Sci,2000,21(5):181-187.
[20]Reuse S,Calao M,Kabeya K,et al.Synergistic activation of HIV-1expression by deacetylase inhibitors and prostratin:implications for treatment of latent infection[J].PLoS One,2009,4(6):e6093.
[21]Burnett J C,Lim K I,Calafi A,et al.Combinatorial latency reactivation for HIV-1subtypes and variants[J].J Virol,2010,84(12):5958-5974.
[22]Ruelas D S,Greene W C.An integrated overview of HIV-1latency[J].Cell,2013,155(3):519-529.
[23]Han Y,Lassen K,Monie D,et al.Resting CD4+T cells from human immunodeficiency virus type 1(HIV-1)-infected individuals carry integrated HIV-1genomes within actively transcribed host genes[J].J Virol,2004,78(12):6122-6133.
[24]Jordan A,Bisgrove D,Verdin E.HIV reproducibly establishes a latent infection after acute infection of T cells in vitro[J].EMBO J,2003,22(8):1868-1877.
[25]Chun T W,F(xiàn)inzi D,Margolick J,et al.In vivo fate of HIV-1-infected T cells:quantitative analysis of the transition to stable latency[J].Nat Med,1995,1(12):1284-1290.
[26]Chun T W,Carruth L,F(xiàn)inzi D,et al.Quantification of latent tissue reservoirs and total body viral load in HIV-1 infection[J].Nature,1997,387(6629):183-188.
[27]Zhu Y,Pe′ery T,Peng J,et al.Transcription elongation factor P-TEFb is required for HIV-1tat transactivation in vitro[J].Genes Dev,1997,11(20):2622-2632.
[28]Mancebo H S,Lee G,F(xiàn)lygare J,et al.P-TEFb kinase is required for HIV Tat transcriptional activation in vivo and in vitro[J].Genes Dev,1997,11(20):2633-2644.
[29]Zhou Q,Chen D,Pierstorff E,et al.Transcription elongation factor P-TEFb mediates Tat activation of HIV-1 transcription at multiple stages[J].EMBO J,1998,17(13):3681-3691.
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評(píng)價(jià)·高一版(2020年6期)2020-11-02 02:45:24
媽媽寶寶(2019年10期)2019-10-26 02:45:34
中國生殖健康(2019年3期)2019-02-01 06:12:26
鐵道通信信號(hào)(2018年11期)2019-01-19 01:15:08
電子制作(2018年11期)2018-08-04 03:25:42
鐵道通信信號(hào)(2018年2期)2018-04-18 12:18:10
鐵道通信信號(hào)(2016年11期)2016-06-01 12:11:32
鑿巖機(jī)械氣動(dòng)工具(2016年3期)2016-03-01 04:00:25
中國病理生理雜志(2015年8期)2015-12-21 12:38:06
