光強和二氧化碳濃度變化對滸苔幼苗生長及生理的影響
2015-01-05 05:13:12高秀秀李亞鶴段維軍徐年軍
海洋學報 2015年10期
高秀秀,李亞鶴,段維軍,徐年軍*
(1. 寧波大學 應用海洋生物技術教育部重點實驗室,浙江 寧波 315211; 2. 浙江省海洋生物工程重點實驗室,浙江 寧波 315211; 3. 寧波出入境檢驗檢疫局技術中心,浙江 寧波 315012)
光強和二氧化碳濃度變化對滸苔幼苗生長及生理的影響
高秀秀1,2,李亞鶴1,2,段維軍3,徐年軍1,2*
(1. 寧波大學 應用海洋生物技術教育部重點實驗室,浙江 寧波 315211; 2. 浙江省海洋生物工程重點實驗室,浙江 寧波 315211; 3. 寧波出入境檢驗檢疫局技術中心,浙江 寧波 315012)
大氣CO2濃度升高引起的海洋酸化如何在光變環境下影響大型海藻固碳量的問題,關系到未來海洋初級生產力的變化趨勢。為研究大型海藻對CO2濃度升高和光強變化的響應,本文選取滸苔(Ulvaprolifera)幼苗為實驗材料,探討其在不同光強下[80、260 μmol/(m2·s)]和兩種CO2濃度(正常CO2濃度:400 μL/L和高CO2濃度:1 000 μL/L)下的生理變化。研究發現,在正常CO2濃度、高光條件下,滸苔幼苗的生長最快,超氧化物特化酶(SOD)活性最高,而過氧化氫酶(CAT)活性在低光、高CO2處理下有最大值。光合色素含量和光系統Ⅱ的光化學效率在不同處理間沒有顯著性差異,但葉綠素a與類胡蘿卜素的比值在低光正常CO2處理下有最大值。同時,高光高CO2處理下,滸苔幼苗的可溶性蛋白含量最低。
CO2;光強;滸苔幼苗;葉綠素熒光效率;抗氧化活性
1 引言
化石燃料的燃燒和毀林的加劇導致大氣中CO2濃度逐漸升高,到21世紀末可增至800~1 000 μL/L(IPCC預測模型A1F1)[1],其中約48%被海洋吸收,導致海水中CO2濃度增加,海水pH值降低[2]。關于海水pH值降低對海洋生態系統影響的研究多集中于浮游植物及遠洋區域[3—4]。然而,有研究表明,對于近岸海域,盡管其pH值的變化較大,但由于富營養化及植物的呼吸作用,其酸化速率要快于大洋區域[5—6]。近年來,生活在近海海域的大型海藻,如褐藻門的巨藻[7]、毛孢藻[8]、綠藻門的石莼[9]以及紅藻門的珊瑚藻[10]和龍須菜[11],對海水pH值降低的響應也得到了廣泛關注。在已研究的大型海藻中,CO2濃度升高對其生長的影響呈現出增加、抑制和不受影響等特征,具有顯著的種間差異。
海水pH值降低是海洋環境變化中的一個重要影響因子,同時由于季節、云層厚度、水體混合、層化、透明度及光周期的影響,再加上生活在潮間帶的大型海藻,隨著海水潮汐變化,經歷著水生-干出-水生的循環,其所接受的光強也隨之變化。已有的研究表明,海水pH值降低對浮游植物的影響受溫度、光強的影響[12—14]。而作為重要經濟海藻的滸苔,廣泛分布在入海河口、海岸帶的潮間帶等的區域[15],其光合生理、生化特性是如何響應海水pH值降低的,又是如何受光變環境調控的,需要進一步的研究。
本研究選取滸苔(Ulvaprolifera)幼苗為材料,探討其對海水pH值降低和光強的復合響應,并分析了其響應機制。
2 材料和方法
2.1 材料
滸苔幼苗充氣培養于光照培養箱中,充氣速率為600 mL/min,培養光強為80 μmol/(m2·s),培養溫度為20℃,光周期為12 h∶12 h,培養基采用過濾滅菌的天然海水,并根據Provasoli培養基加富,每3 d更換一次在此環境條件下適應10 d,確保幼苗生長狀態良好。
2.2 實驗設計
適應狀態良好的滸苔接種于新鮮培養基中,接種濃度為0.25 g/L(經預實驗可知,該接種濃度可確保整個實驗過程中充氣狀態下培養體系的pH基本穩定)。設置兩個光強,分別為80、260 μmol/(m2·s),光周期為12 h∶12 h,培養溫度為20°C。兩個CO2濃度分別設置為400 μL/L(正常CO2濃度,記為LC)、1 000 μL/L(高CO2濃度,記為HC),LC、HC處理均在二氧化碳培養箱(RXZ-160B CO2培養箱、寧波江南儀器廠)中充氣培養(LC:直接充室外空氣;HC:CO2和空氣的混合氣體)。高光正常CO2、高光低CO2、低光高CO2和低光正常CO2處理分別記為HH、HL、LH和LL。
2.3 生長速率的測定
每2 d測定藻體的鮮質量,相對生長速率(RGR,單位:%·d-1)
RGR=100×(lnWn-lnW0)/(tn-t0)×100,
(1)
式中,Wn為tn天的藻體鮮質量。
2.4 色素的提取及測定
在光周期中期取0.02 g的藻體放入離心管中,加入100% 甲醇,4°C冰箱中過夜提取,之后離心,取上清液于酶標儀中進行全波段掃描(400~700 nm)。根據公式計算葉綠素a(Chla)、葉綠素b(Chlb)、類胡蘿卜素(Car,cartenoids)的含量[16]:
CChla=15.65 A666-7.34 A653,
(2)
CChlb=27.05 A653-11.21 A666,
(3)
CCar=(1000 A470-2.86 CChla-129.2 CChlb)/221.
(4)
2.5 葉綠素熒光參數的測定
葉綠素熒光參數采用氙燈脈沖調制熒光儀(water-PAM)進行測定。
在光周期中期快速取樣(1~2 min)進行快速光響應曲線和誘導曲線測定。樣品暗處理15 min后測定葉綠素熒光誘導曲線,光化光強度設置為培養光強。
快速光響應曲線(RLC)的測定使用8個光強梯度[226、337、533、781、1 077、1 593、2 130和2 854 μmol/(m2·s)]的光化光,每個光強處理時長設定為10 s。 相對電子傳遞速率(rETR)通過以下的公式計算:
rETR=yield×0.5×PFD,
(5)
式中,yield代表光系統Ⅱ的有效光化學效率,系數0.5代表光系統Ⅱ吸收的光量子占總量的50%,PFD(photon flux density)代表光化光的強度[μmol/(m2·s)]。快速光響應曲線根據Webb 等[17]進行擬合,公式如下:
rETR=rETRmax×(1-eαx/rETRmax),
(6)
Ik=rETRmax/α.
(7)
2.6 抗氧化活性測定
SOD活性測定采用氮藍四唑法[18],CAT的測定采用過氧化氫法(H2O2)。取0.1 g樣品洗凈后置于預冷的研缽中,加1.6 mL 50 mmol/L預冷的磷酸鹽緩沖液(pH 7.8)在冰浴上研磨成勻漿。轉入離心管在4℃、10 000 g下離心20 min,上清液即為酶液。SOD測定取3 mL反應混合液[甲硫氨酸溶液、EDTA-Na2、磷酸緩沖液(pH 7.8)和氮藍四唑溶液]和30 μL酶液于試管中,將試管置于光照培養箱中在4 000 lx光照下反應20 min,測定OD560值,通過公式計算SOD活性;CAT的測定取3 mL反應液[磷酸緩沖液(pH 7.0)和30%的H2O2]加入0.1 mL酶液,測定OD240(紫外)值(測定40 s),通過公式計算CAT酶活。
2.7 可溶性蛋白含量測定
考馬斯亮藍G250法測定。處理方法同上,獲得滸苔酶液提取液。取20 μL提取液加入80 μL,0.05 mol/L,pH 7.8磷酸緩沖液,再加入2.9 mL考馬斯亮藍G250溶液,充分混合,反應2 min后測定OD595,并通過標準曲線計算出蛋白質含量。可溶性蛋白的標準曲線:y=6.025x-0.023 5,R2=0.999 5。
2.8 可溶性糖的測定
蒽酮硫酸比色法測定。取0.1 g的新鮮滸苔,用純水定容至10 mL,沸水浴1 h,過濾,殘渣再沸水浴兩次,每次30 min,合并濾液,取1 mL待測液,緩慢加入3 mL蒽酮試劑(2.0 g/L),立即沸水浴10 min,然后冷卻至室溫,檢測其OD620值,并通過標準曲線計算可溶性糖的量。葡萄糖標準曲線:y=0.035 9x-0.005 8,R2=0.997 6(含量范圍0~80 μg)。
2.9 數據分析
數據處理使用Origin 7.0軟件,采用One-Way ANOVA,Turkey進行顯著性分析,設顯著水平為p<0.05,通過在圖中或者表格中標注不同的小寫字母來表示差異顯著性。
3 結果和分析
3.1 不同處理下的生長和色素含量
正常CO2條件下,光強增加顯著提高了滸苔幼苗的生長(p=0.008),表現為相對生長速率增加58%,但在高CO2條件下,光強對滸苔幼苗的生長影響不顯著(p=0.809)。低光條件下,CO2濃度升高顯著增加了滸苔幼苗的生長(p=0.006),增幅約為45%,而在高光條件下,CO2濃度升高抑制了滸苔幼苗的生長,約降低11%,但其差異性不顯著(p=0.392)(圖1)。

圖1 不同CO2和光強處理對滸苔幼苗生長的影響Fig.1 Effects of different CO2 and light intensity on the growth of U. prolifera seedling數值為平均值±標準偏差,n=3;橫坐標代表不同的處理,其中,HH、HL、LH和LL分別代表高光高CO2、高光正常CO2、低光高CO2和低光正常CO2處理,不同字母表示不同處理間差異顯著(p < 0.05)Data=mean+SD,n=3;the X axis represents different treatments,HH,HL,LH,LL represent high light and high CO2,high light and normal CO2,low light and high CO2,low light and normal CO2 treatment respectively. Different letters indicated significant difference between treatments(p<0.05)
在整個實驗過程中,滸苔幼苗的Chla、Chlb及類胡蘿卜素的含量分別維持在988 μg/g、751 μg/g、174 μg/g,不同處理間差異不顯著(見圖2),相比于其他處理,低光正常CO2條件下,Chla與Chlb含量的比值(CChl a/CChl b)降低,而CChl a/CCar的值升高,同時,CChl b/CCar的值也略有增加(表1)。

表1 不同CO2和光強處理對滸苔幼苗Chl a、Chl b與Car含量比值的影響Tab.1 Effects of different CO2 and light intensity on the ratios of chlorophyll a、b and cartenoids of U. prolifera seedling
注:數值為平均值±標準偏差,n=3。
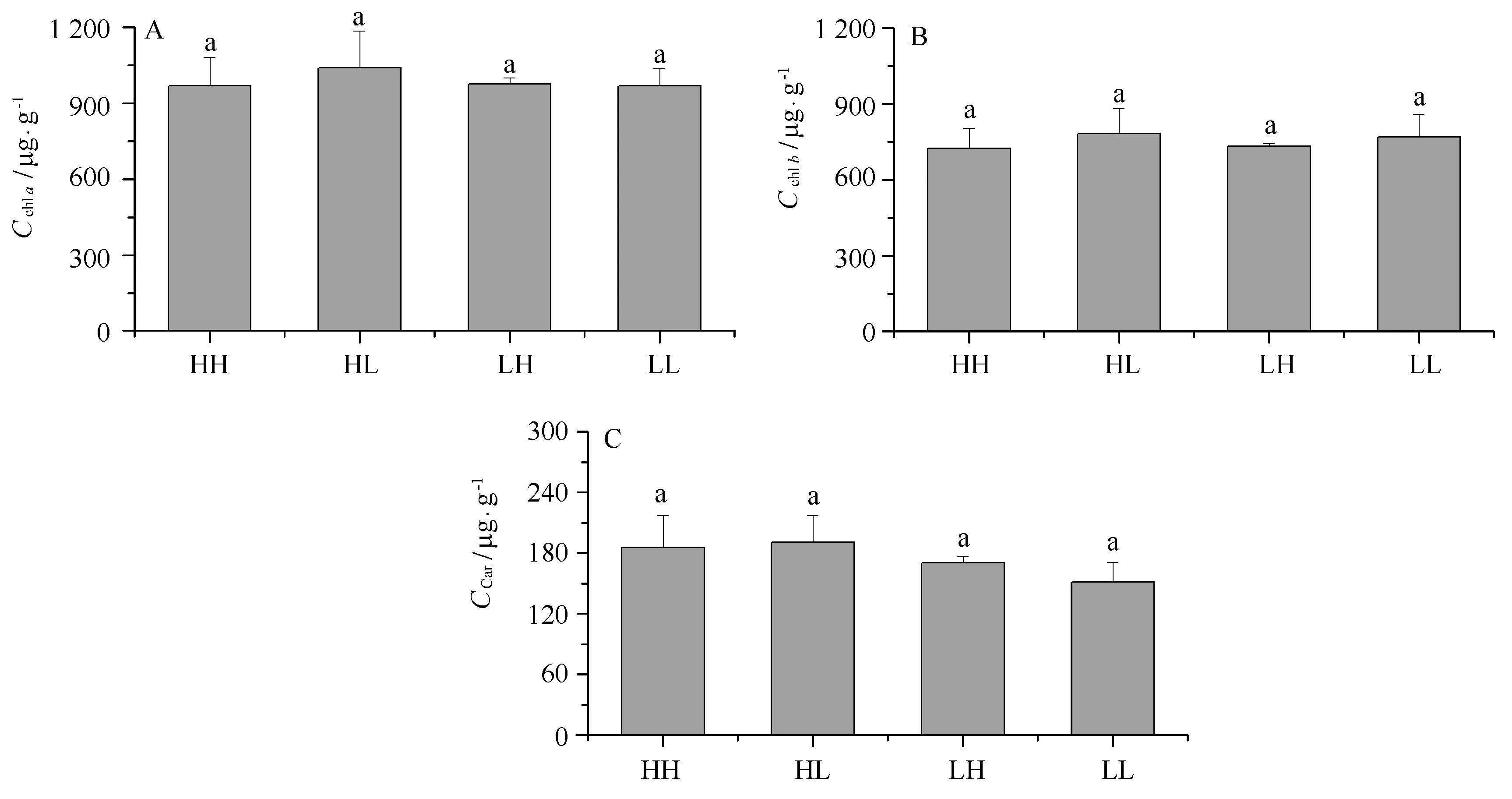
圖2 不同CO2和光強處理對滸苔幼苗光合色素的影響Fig.2 Effects of different CO2 and light intensity on the photosynthesis pigments of U. prolifera seedling數值=平均值±標準偏差,n=3; HH、HL、LH和LL和不同字母表示的含義同圖1Data=mean+SD,n=3; HH,HL,LH,LL and all letters showed the same meaning as that in Figure 1
3.2 不同處理下的光化學參數
相比于低光處理,高光處理下的滸苔幼苗最大光合效率略有增加,但差異不顯著(p>0.06),而有效光化學效率在低光下的值顯著高于高光處理(低CO2:p=0.012;高CO2:p=0.019 )。相比于光強變化,CO2濃度變化對滸苔幼苗光合效率的影響差異不顯著(圖3)。滸苔幼苗的相對電子傳遞速率在高光高CO2條件下較高(見圖4)。根據圖4計算出的相對電子傳遞速率、飽和光強、光能利用效率如表2所示。相對于低光處理,高光處理的滸苔幼苗在高、低CO2條件下相對電子傳遞速率(rETR)增強,但差異不顯著;同時飽和光強(Ek)也增強;相比于正常CO2處理,高CO2條件下滸苔幼苗的rETRmax、Ek均升高,而表觀光能利用效率(α)則呈現相反趨勢。

圖3 不同CO2和光強處理對滸苔幼苗葉綠素熒光效率的影響Fig.3 Effects of different CO2 and light intensity on the chlorophyll fluorescence efficiency of U. prolifera seedlingA為最大光合效率(Fv/Fm);B為有效光合效率(Fv′/Fm′);數值為平均值±標準偏差,n=3;HH、HL、LH和LL和不同字母表示的含義同圖1A is Fv/Fm and B is Fv′/Fm′; data=mean+SD,n=3; HH,HL,LH,LL and all letters showed the same meaning as that in Figure 1
3.3 不同處理下的抗氧化活性
相比于高光處理,滸苔幼苗的超氧化物歧化酶(SOD)的活性在低光條件下顯著增加,高、正常CO2處理下,增幅約為57%、42%。相比于正常CO2濃度處理,低光下,高CO2處理的滸苔幼苗具有略高的活性,但差異不顯著(p=0.502)(見圖5A)。過氧化氫酶(CAT)的活性在不同處理間差異顯著,低光條件下,CO2濃度升高,其活性顯著提高(p=0.008)。而高光條件下,盡管差異不顯著(p=0.124),但CO2濃度升高提高了CAT的活性。總體來講,其活性隨著CO2濃度的增加而升高(圖5B)。

表2 滸苔幼苗正常CO2和高CO2及不同光強下的光合作用速率(rETR)與光強關系的最佳擬合參數Tab.2 The fitted parameters derived from rapid light curves of U. prolifera seedling grown under both normal,high CO2and different light intensities
注:Ek為飽和光強 [μmol/(m2·s)],rETRmax為最大相對電子傳遞速率,α為表觀光能利用效率;數據為平均值±標準偏差,n=3;HH、HL、LH和LL表示的含義同圖1。

圖4 不同CO2和光強處理對滸苔幼苗相對電子傳遞速率的影響Fig.4 Effects of different CO2 and light intensity on the rETR of U. prolifera seedling數值為平均值±標準偏差,n=3; HH、HL、LH和LL表示的含義同圖1Data=mean+SD,n=3; HH,HL,LH,LL showed the same meaning as that in Figure 1

圖5 不同CO2和光強處理對滸苔幼苗SOD (A) 和CAT (B) 活性的影響Fig.5 Effects of different CO2 and light intensity on the SOD (A) and CAT (B) of U. prolifera seedling數值為平均值±標準偏差,n=3;HH、HL、LH和LL和不同字母表示的含義同圖1Data=mean+SD,n=3;HH,HL,LH,LL and all letters showed the same meaning as that in Figure 1
3.4 不同處理下的可溶性糖、可溶性蛋白等含量
相比于高光處理,低光條件下滸苔可溶性蛋白含量顯著增加,在高、正常CO2條件下,增長幅度分別為:47%、13%。相比于高CO2處理,高光下正常CO2滸苔的可溶性蛋白含量增加顯著,而低光下含量變化較小(p=0.108)(見圖6A)。相比之下,可溶性糖的含量在各處理條件下基本保持穩定,各組之間并無顯著差異。
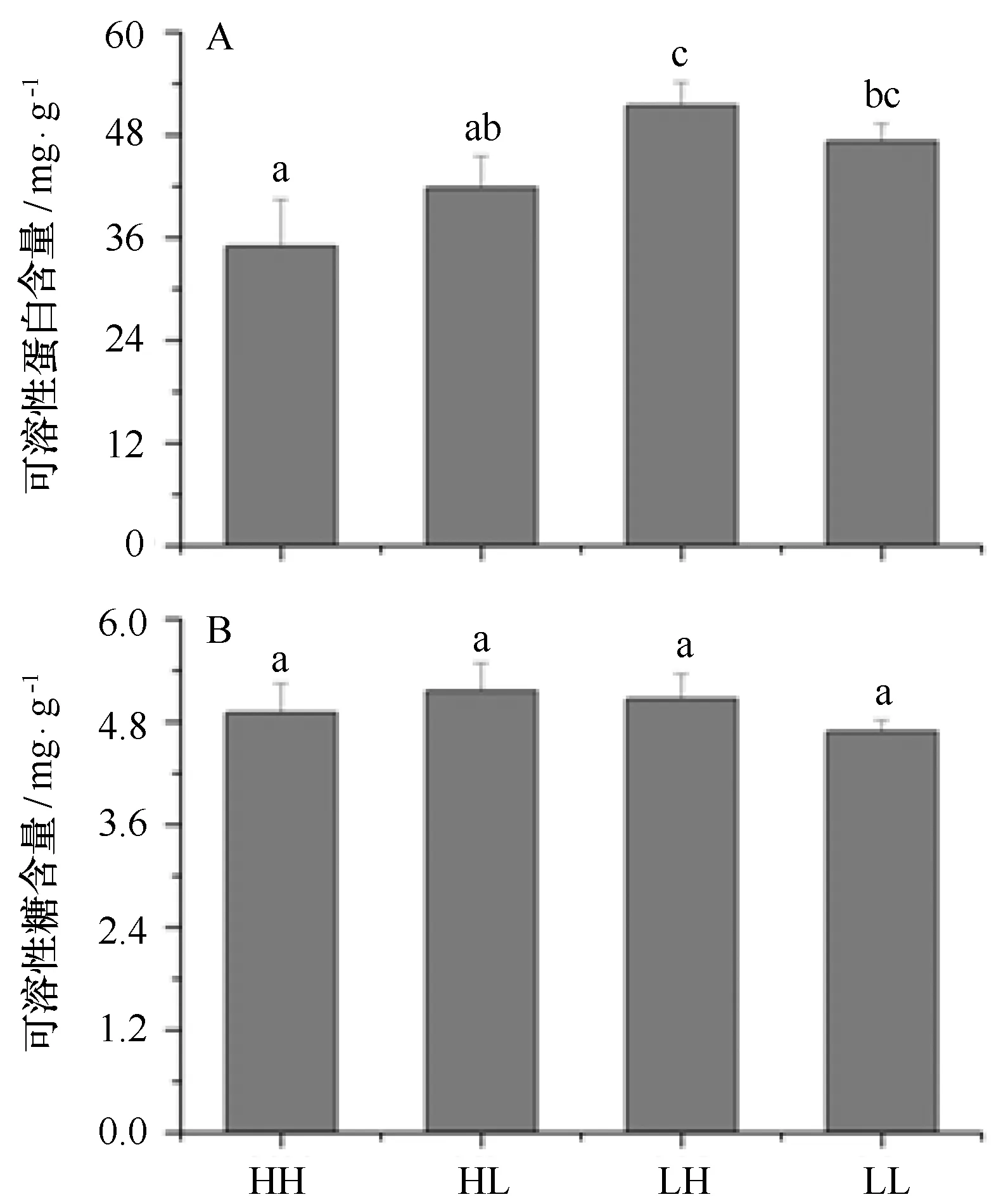
圖6 不同CO2和光強處理對滸苔幼苗可溶性蛋白 (A) 和可溶性糖 (B) 含量的影響Fig.6 Effects of different CO2 and light intensity on the dissolved protein (A) and carbohydrate (B) of U. prolifera seedling數值為平均值±標準偏差,n=3;HH、HL、LH和LL和不同字母表示的含義同圖1Data=mean+SD,n=3;HH,HL,LH,LL and all letters showed the same meaning as that in Figure 1
4 討論
生長狀況是體現藻類的最直觀指標,每一種藻都有適宜的光強范圍,當光照超過光補償點時,藻體積累有機物才會生長[19—20]。葉綠素含量與藻類光合參數的變化是判斷藻類是否正常生長的重要指標,決定著藻類吸收和轉化光能的高低,在一定程度上反映了對光強的耐受能力[21]。光強提高在一定程度上為藻體的光合作用提供更多的光能,從而促進藻體生長。CO2濃度升高能促進大型海藻光合作用,這在過去的研究中多有體現[22—25]。其原因主要是,CO2濃度的升高,能夠提高培養水體中的無機碳濃度,促進光合固碳作用,同時下調了藻體的無機碳濃縮機制,節省了能量[26],從而更利于藻體的生長。這與本研究中低光強處理的結果一致。而在高光條件下,高CO2濃度抑制了滸苔幼苗的生長。該現象在微藻中已有報道[12,27—28],然而在大型藻類中鮮有研究。本研究的結果顯示,光強增加并未降低滸苔幼苗的Chla含量,其原因可能是光強處理的時間較短,對藻體葉綠素含量的響應沒有得到及時的體現,也正因為如此,相比于低光強處理,CO2濃度增加為高光生長的滸苔幼苗提供了過多的能量。同時,SOD和CAT參與組成植物體內重要的抗氧化酶系統,起到減弱或清除自由基和過氧化物的毒害作用,可提高逆境脅迫下藻體對環境的耐受能力。本研究中,高光條件下,SOD和CAT活性較低,即表明適當提高光強,可增強藻體對光能的利用效率,促進藻體的生長[29],此時,CO2濃度升高,藻體無機碳濃縮機制下調,節省的部分能量加速了高光下的光抑制,進而在一定程度上抑制了滸苔幼苗的生長。
可溶性蛋白和可溶性糖含量的改變是植物感應所受脅迫環境的指標,在逆境脅迫環境下,通過改變糖和蛋白的含量水平來感應外界環境變化或提供必要的信號傳導,調節植物的滲透壓[29]。而碳水化合物含量的變化也可能直接或間接的影響光合作用器中的有關組分[30—31],并有可能阻礙CO2擴散進入葉綠體[32],進而影響光合作用。本研究結果顯示,可溶性糖的變化趨勢不明顯,其含量維持在相對穩定的水平,主要是因為可溶性糖變化與滲透壓調節有關,光照和CO2對滸苔滲透壓影響較小,且處理時間較短,并未引起可溶性糖的變化。而可溶性蛋白含量的變化趨勢在高光條件下降低,表明適當提高光強可提高滸苔的光合作用速率,在一定程度上進一步導致其可溶性蛋白的降低。
目前,世界范圍內對海洋酸化的研究還處于起步階段,盡管已經評估的物種約有100種[33],但有關海洋酸化的長期效應和對漁業資源的全部影響還尚不清楚。同時,大型綠藻種類豐富,分布廣泛,且不同地區的優勢種和關鍵種差異顯著,故盡可能的選擇多地區多種類的大型綠藻優勢種,研究其對包括海水pH值變化在內的環境變化的響應,只有系統全面地認識海洋酸化對海洋各區域海洋生物及生態系統所產生的影響,才能找到有效的解決辦法,減少環境變化帶給人類的巨大損失。
[1] Haines A. Climate change 2001: the scientific basis. contribution of working group 1 to the third assessment report of the intergovernmental panel on climate change[J]. International Journal of Epidemiology,2003,32(2): 321-321.
[2] Sabine C L,Feely R A,Gruber N,et al. The oceanic sink for anthropogenic CO2[J]. Science,2004,305(5682): 367-371.
[3] Beardall J,Stojkovic S,Gao Kunshan. Interactive effects of nutrient supply and other environmental factors on the sensitivity of marine primary producers to ultraviolet radiation: implications for the impacts of global change[J]. Aquat Biol,2014,22: 5-23.
[4] H?der D P,Villafae V E,Helbling E W. Productivity of aquatic primary producers under global climate change[J]. Photochemical & Photobiological Sciences,2014,13(10): 1370-1392.
[5] Cai Weijun,Hu Xinping,Huang Weijen,et al. Acidification of subsurface coastal waters enhanced by eutrophication[J]. Nature Geoscience,2011,4(11): 766-770.
[6] Zhai W D,Zheng N,Huo C,et al. Subsurface pH and carbonate saturation state of aragonite on the Chinese side of the North Yellow Sea: seasonal variations and controls[J]. Biogeosciences,2014,11(4): 1103-1123.
[7] Roleda M Y,Morris J N,McGraw C M,et al. Ocean acidification and seaweed reproduction: increased CO2ameliorates the negative effect of lowered pH on meiospore germination in the giant kelpMacrocystispyrifera(Laminariales,Phaeophyceae)[J]. Global Change Biology,2012,18(3): 854-864.
[8] Bender D,Diaz-Pulido G,Dove S. The impact of CO2emission scenarios and nutrient enrichment on a common coral reef macroalga is modified by temporal effects[J]. Journal of Phycology,2014,50(1): 203-215.
[9] Olischl?ger M,Bartsch I,Gutow L,et al. Effects of ocean acidification on growth and physiology ofUlvalactuca(Chlorophyta) in a rockpool-scenario[J]. Phycological Research,2013,61(3): 180-190.
[10] Hofmann L C,Straub S,Bischof K. Elevated CO2levels affect the activity of nitrate reductase and carbonic anhydrase in the calcifying rhodophyteCorallinaofficinalis[J]. Journal of Experimental Botany,2013,64(4): 899-908.
[11] 徐軍田,高坤山. 二氧化碳和陽光紫外輻射對龍須菜生長和光合生理的影響[J]. 海洋學報,2010,32(5): 144-151.
Xu Juntian,Gao Kunshan. The influence of carbon dioxide and solar UVR on the growth,photosynthesis and pigments contents ofGracilarialemaneiformis[J]. Haiyang Xuebao,2010,32(5): 144-151.
[12] Gao Kunshan,Xu Juntian,Gao Guang,et al. Rising CO2and increased light exposure synergistically reduce marine primary productivity[J]. Nature Climate Change,2012,2(7): 519-523.
[13] Li Y,Gao K,Villafae V E,et al. Ocean acidification mediates photosynthetic response to UV radiation and temperature increase in the diatomPhaeodactylumtricornutum[J]. Biogeosciences,2012,9(10): 3931-3942.
[14] Xu Juntian,Gao Kunshan,Li Yahe,et al. Physiological and biochemical responses of diatoms to projected ocean changes[J]. Marine Ecology Progress Series,2014,515: 73-81.
[15] 呂永林,張永普,李凱,等. 浙江洞頭大竹嶼島潮間帶大型底棲生物多樣性[J]. 生態學雜志,2011,30(4): 707-716.
Lv Yonglin,Zhang Yongpu,Li Kai,et al. Macrobenthos diversity in intertidal zone of Dazhuyu Island,Dongtou County of Zhejiang Province[J]. Chinese Journal of Ecology,2011,30(4): 707-716.
[16] Wellburn A R. The spectral determination of chlorophyllsaandb,as well as total carotenoids,using various solvents with spectrophotometers of different resolution[J]. Journal of Plant Physiology,1994,144(3): 307-313.
[17] Webb W L,Newton M,Starr D. Carbon dioxide exchange ofAlnusrubra[J]. Oecologia,1974,17(4): 281-291.
[18] 李合生. 植物生理生化實驗原理和技術[M]. 北京: 高等教育出版社,2000: 267-268.
Li Hesheng. Experimental principle and technology of plant physiology and biochemistry[M]. Beijing:Higher Education Press, 2000:267-268.
[19] Lapointe B E,Tenore K R,Dawes C J. Interactions between light and temperature on the physiological ecology ofGracilariatikvahiae(Gigartinales: Rhodophyta)[J]. Marine Biology,1984,80(2): 161-170.
[20] 劉雅萌,徐軍田,李信書,等. 不同增殖方式來源的綠潮藻滸苔藻體生長及光合生理特性差異[J]. 水產學報,2014,38(5): 691-696.
Liu Yameng,Xu Juntian,Li Xinshu,et al. The effects of different proliferative ways for the green tide algaUlvaproliferaon their growth and photophysiological performances[J]. Journal of Fisheries of China,2014,38(5): 691-696.
[21] Havaux M,Tardy F. Thermostability and photostability of photosystem Ⅱ in leaves of theChlorina-f2 barley mutant deficient in light-harvesting chlorophylla/bprotein complexes[J]. Plant Physiology,1997,113(3): 913-923.
[22] Zou Dinghui. Effects of elevated atmospheric CO2on growth,photosynthesis and nitrogen metabolism in the economic brown seaweed,Hizikiafusiforme(Sargassaceae,Phaeophyta)[J]. Aquaculture,2005,250(3/4): 726-735.
[23] Wu Hongyan,Zou Dinghui,Gao Kunshan. Impacts of increased atmospheric CO2concentration on photosynthesis and growth of micro-and macro-algae[J]. Science in China Series C: Life Sciences,2008,51(12): 1144-1150.
[24] Xu Zhiguang,Zou Dinghui,Gao Kunshan. Effects of elevated CO2and phosphorus supply on growth,photosynthesis and nutrient uptake in the marine macroalgaGracilarialemaneiformis(Rhodophyta)[J]. Botanica Marina,2010,53(2): 123-129.
[25] 鄒定輝,高坤山. 高CO2濃度對大型海藻光合作用及有關過程的影響[J]. 生態學報,2002,22(10): 1750-1757.
Zou Dinghui,Gao Kunshan. Effects of elevated CO2concentration on the photosynthesis and related physiological processes in marine macroalgae[J]. Acta Ecologica Sinica,2002,22(10): 1750-1757.
[26] 鄧亞運,鄒定輝. 大氣CO2濃度升高對不同氮生長條件下的兩種大型海藻光合作用的影響[J]. 生態學雜志,2014,33(6): 1520-1527.
Deng Yayun,Zou Dinghui. Effects of elevated atmospheric CO2on photosynthesis ofGracilarialemaneiformisandUlvaconglobatagrown at low and high N supplies[J]. Chinese Journal of Ecology,2014,33(6): 1520-1527.
[27] Li Yahe,Xu Juntian,Gao Kunshan. Light-modulated responses of growth and photosynthetic performance to ocean acidification in the model diatomPhaeodactylumtricornutum[J]. PLoS One,2014,9(5): e96173.
[28] Li Gang,Campbell D A. Rising CO2interacts with growth light and growth rate to alter photosystem Ⅱ photoinactivation of the coastal diatomThalassiosirapseudonana[J]. PLoS One,2013,8(1): e55562.
[29] 朱政,蔣家月,江昌俊,等. 低溫脅迫對茶樹葉片SOD、可溶性蛋白和可溶性糖含量的影響[J]. 安徽農業大學學報,2011,38(1): 24-26.
Zhu Zheng,Jiang Jiayue,Jiang Changjun,et al. Effects of low temperature stress on SOD activity,soluble protein content and soluble sugar content inCamelliasinensisleaves[J]. Journal of Anhui Agricultural University,2011,38(1): 24-26.
[30] Nie G,Hendrix D L,Webber A N,et al. Increased accumulation of carbohydrates and decreased photosynthetic gene transcript levels in wheat grown at an elevated CO2concentration in the field[J]. Plant Physiology,1995,108(3): 975-983.
[31] Gesch R W,Boote K J,Vu J C V,et al. Changes in growth CO2result in rapid adjustments of ribulose-1,5-bisphosphate carboxylase/oxygenase small subunit gene expression in expanding and mature leaves of rice[J]. Plant Physiology,1998,118(2): 521-529.
[32] Makino A,Mae T. Photosynthesis and plant growth at elevated levels of CO2[J]. Plant and Cell Physiology,1999,40(10): 999-1006.
[33] Kroeker K J,Kordas R L,Crim R,et al. Impacts of ocean acidification on marine organisms: quantifying sensitivities and interaction with warming[J]. Global Change Biology,2013,19(6): 1884-1896.
The effects of light and increased CO2on the growth and physiological performances in marine green algae Ulva prolifera seedling
Gao Xiuxiu1,2,Li Yahe1,2,Duan Weijun3,Xu Nianjun1,2
(1.KeyLaboratoryofAppliedMarineBiotechnologyofMinistryofEducation,NingboUniversity,Ningbo315211,China; 2.KeyLaboratoryofMarineBiotechnologyofZhejiangProvince,Ningbo315211,China; 3.NingboEntry-ExitInspectionandQuarantineBureauTechnologyCentre,Ningbo315012,China)
Ocean acidification due to increasing anthropogenic CO2is expected to influence marine primary productivity. In order to investigate the interactive effects of increased light and CO2changes on macroalgae,we grewUlvaproliferaseedling under the normal CO2level (400 μL/L) and elevated CO2level (1 000 μL/L) conditions,the light levels set as 80 and 260 μmol/(m2·s),and measured its physiological performance. Research finding that the specific growth rate and activity of SOD was highest under high light and normal CO2level . However,the activity of CAT showed the highest value under the high CO2and low light conditions. Under the different treatments,there was no significant effect on the chlorophyllaand maximal photochemical yield of PSⅡ (Fv/Fm),while the largest ratios of chlorophylla to carotenoid occurred at nomal CO2and low light conditions. Meanwhile,the content of soluble protein was decreased by high light and high CO2.
CO2; light;Ulvaproliferaseedling; chlorophyll fluorescence efficiency; Antioxidant activity
10.3969/j.issn.0253-4193.2015.10.008
2015-02-11;
2015-07-21。
國家自然科學基金項目(40876073, 41276122);國家教育部博士點基金博導基金(20123305110002); 寧波大學學科項目(XKL14D2085);寧波大學科研啟動項目(F01259144702)。
高秀秀(1989—),女,山東省濟南市人,主要研究藻類生理與分子生物學。E-mail:ggaoxiuxiu@163.com
*通信作者:徐年軍(1973—),男,湖北省赤壁市人,博士,博士生導師,研究員,主要研究方向為海洋天然藥物,海洋生物資源綜合利用等。E-mail:xunianjun@nbu.edu.cn
S917.3
A
0253-4193(2015)10-0080-08
高秀秀,李亞鶴,段維軍,等. 光強和二氧化碳濃度變化對滸苔幼苗生長及生理的影響[J].海洋學報,2015,37(10):80—87,
Gao Xiuxiu,Li Yahe,Duan Weijun,et al. The effects of light and increased CO2on the growth and physiological performances in marine green algaeUlvaproliferaseedling[J]. Haiyang Xuebao,2015,37(10):80—87,doi:10.3969/j.issn.0253-4193.2015.10.008
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
媽媽寶寶(2017年3期)2017-02-21 01:22:28
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
中國塑料(2016年3期)2016-06-15 20:30:00
