低溫蛋白酶產生菌的篩選及其酶學性質的初步研究
2015-01-08 09:52:30莫清珊張會圖孫同韋馮士元路福平
天津科技大學學報 2015年3期
莫清珊 ,張會圖,田 耀,孫同韋,馮士元,孫 軍,路福平
(1.天津科技大學生物工程學院,天津 300457;2.天津科技大學海洋科學與工程學院,天津 300457)
低溫蛋白酶是指最適催化溫度在40,℃以下,并且在20~30,℃仍能保持較高酶活(50%以上)的一類蛋白水解酶[1].由于其最適催化溫度接近自然環境中的溫度,因此在應用過程中可省去加熱或冷卻過程,與中高溫蛋白酶相比具有節能、省時等特點,在食品及洗滌行業具有廣泛的應用前景.低溫蛋白酶多數來源于冰川、極地、高山、深海等低溫環境中的嗜低溫或耐低溫微生物[2–3].這些低溫微生物為了適應其所處的低溫環境,常可表達分泌一些在低溫條件下仍具有較高催化活性的胞外酶或胞內酶,因此從耐低溫或嗜低溫微生物中篩選獲得低溫蛋白酶已成為發掘新型工業用低溫酶制劑的主要方法[4].目前已發現的產低溫蛋白酶菌株有來源于冰川凍土中的地衣芽孢桿菌(Bacillus licheniformis)和短小桿菌(Curtobacterium luteum)、來源于南極地區的梭狀芽孢桿菌(Clostridium sp.)和嗜冷桿菌(Psychrobacter proteolyticus)、來源于海洋浮冰中的科爾韋爾氏菌屬(Colwellia sp.)、來源于寒漠地區的微小桿菌(Exiguobacterium sp.)以及來源于其他寒冷環境中的沙雷氏菌(Serratia sp.)、弧菌(Vibrio sp.)、黃單胞菌(Xanthomonas maltophilia)、希瓦氏菌屬(Shewanella sp.)產黃青霉(Penicillium chrysogenum)等.這些菌株所產低溫蛋白酶的最適催化溫度大部分均在30~40,℃,只有少數幾種的最適催化溫度低于20,℃,但穩定性很差.國內對低溫蛋白酶及其產生菌的研究起步較晚,研究對象多集中在來源于冰川及凍土中的假單胞菌屬 (Pseudomonas)、黃桿菌屬(Xanthomonas)、產氣單胞菌屬(Aeromonas)等.本研究則從深海沉積物中分離到1 株產低溫蛋白酶動性球菌(Planococcus sp.),并對該菌株的生長特性、產酶特性及其所產蛋白酶的酶學性質進行了研究.
1 材料與方法
1.1 菌株來源
嗜冷菌株由天津科技大學孫軍教授提供的南印度洋的深海沉積物中分離得到.
1.2 主要試劑和儀器
連接酶、Taq DNA 聚合酶、dNTP、T 載體、DNA Marker,寶生物工程(大連)有限公司;酪蛋白、瓊脂粉、緩沖液試劑,上海生工生物工程有限公司.
PCR 儀、凝膠成像儀、電泳系統,美國Bio-Rad公司;1500–201 型全波長酶標儀,美國熱電公司.
1.3 培養基
酪蛋白篩選培養基:酪蛋白20,g,瓊脂15,g,人工海水定容到1,L,調節pH 到7.5~8.0.
發酵培養基:ZoBell 2216E 培養基.
1.4 產低溫蛋白酶菌株的分離與鑒定
1.4.1 產低溫蛋白酶菌株的分離
取深海沉積物樣品1,g,加入20,mL 生理鹽水,渦旋振蕩混勻.按10 倍稀釋法稀釋成10-2、10-3、10-4、10-5、10-6、10-76 個稀釋度的稀釋液,每個稀釋度各取0.2,mL 稀釋液涂布于3 個酪蛋白篩選培養基平板上,并分別于4、20、25,℃培養72,h.選取菌落周圍有明顯蛋白水解圈的菌落于酪蛋白分離培養平板上進行三區劃線純化培養.根據水解圈直徑dH與菌落直徑dc的比值確定菌株低溫條件下產蛋白酶能力的大小.
1.4.2 16S,rDNA 的克隆及系統進化分析
菌體基因組提取參照文獻[5]進行;以上述基因組為模板,以細菌 16S rDNA 通用引物 27,F(5'-AGAGTTTGATCCTGGCTCAG-3'),1492,R(5'-AGT AAGGAGGTGATCCAACCGCA-3')對16S rDNA 進行PCR 擴增.PCR 反應條件為:94,℃預變性5,min,然后94,℃ 60,s,55,℃ 90,s,72,℃ 120,s,循環30 次,72,℃ 延伸10,min.擴增產物經0.8%瓊脂糖凝膠電泳回收后克隆至T 載體,并委托北京華大基因公司測序.將所得序列在NCBI 網站進行 Blastn 比對(www.ncbi.nlm.nih.gov/BLAST),確定與其相關的種屬特性.
1.4.3 菌株形態及生理生化特性鑒定
最適生長溫度、菌落形態、革蘭氏染色、氧化酶活性檢測、過氧化氫酶活性檢測、明膠液化實驗參照文獻[6]進行.
1.5 蛋白酶粗酶液的制備及酶活測定
將種子培養液以2%的體積比接種至50,mL 發酵培養基,分別在不同溫度下(15、20、25,℃),200,r/min振蕩培養52,h,定時取樣,考察不同培養溫度對產酶的影響.
收集菌液于4,℃、10,000,r/min 離心30,min,收集上清液.
以酪蛋白為底物的酶活的測定方法參照文獻[7]進行;根據QB/T 1803—1993《工業酶制劑通用試驗方法》,以1,mL 酶液在37,℃、pH 8.0 條件下,每分鐘反應產生1,μg 酪氨酸所需要的酶量為1 個酶活力單位(U/mL).
粗酶液用緩沖溶液稀釋至適當濃度,作為待測酶液.緩沖液配制1%的酪素溶液作為底物并將粗酶液稀釋適當的倍數.取1,mL 稀釋的酶液,37,℃保溫2,min,加入同樣溫度的底物 1,mL,于 37,℃反應10,min,加入2,mL 質量分數10%三氯乙酸終止反應.靜置離心,取1,mL 上清液,加入5,mL 0.4,mol/L Na2,CO3溶液、1,mL 福林酚試劑,混勻,40,℃保溫20,min,測定吸光度A680.以滅活酶液為空白對照.
1.6 酶學性質分析
1.6.1 溫度對蛋白酶酶活的影響
分別將1,mL 粗酶液與1,mL 水解酪蛋白溶液(1%,pH 8.0)混勻,分別在4~50,℃ 測定酶活力,以酶活力最高時為100%,計算其他條件下的相對酶活力.
1.6.2,pH 對蛋白酶酶活的影響
用不同pH 的乳酸鈉緩沖液(pH 2.0、3.0、4.0、5.0)、磷酸鹽緩沖液(pH 6.0、7.0、7.5、8.0)、硼砂緩沖液(pH 9.0、10.0)將待測酶液進行適當的稀釋,在37,℃及相應pH 條件下測定酶活,酶活數值最大者計為100%,所對應的pH 即為待測酶最適反應pH,其他pH 下的酶活與最高酶活的比值即為其相對酶活.
1.6.3 蛋白酶的熱穩定性檢測
將粗酶液分別在10、37、45,℃保溫30、60、90、120,min 后測定殘余酶活,以未進行保溫處理的酶活力值作為對照,設定其相對酶活力為100%,得出酶活力隨保溫時間變化的曲線.
1.6.4 不同金屬離子及化學試劑對蛋白酶酶活的影響
將待測酶液置于不同濃度(5、1,mmol/L)的Ca2+、Mg2+、Cu2+、Ni2+、Co2+、Zn2+、Mn2+、Ba2+緩沖液中,4,℃保溫1,h,在pH 10.0、37,℃條件下測定殘留的酶活.將待測酶液置于EDTA、PMSF、β-ME 緩沖液中,4,℃保溫1,h,在pH 10.0、37,℃條件下測定殘留的酶活.
2 結果與分析
2.1 產低溫蛋白酶菌株的篩選
低溫培養條件下不同菌株的蛋白水解圈形成情況如圖1 所示.在低溫培養條件下(4~10,℃),經透明圈篩選以及劃線純化后復篩,共獲得3 株具有分泌表達蛋白酶能力的菌株,分別將其編號為11813、11815 以及11816.其中菌株11815 的蛋白酶分泌表達能力較強,在培養溫度為10,℃的條件下,24,h 內,其蛋白水解圈直徑與菌落直徑的比值(dH/dc)最高可達2.77;而在相同條件下,菌株11813 及11816 的dH/dc則分別僅有1.78 和1.60.

圖1 低溫培養條件下不同菌株的蛋白水解圈形成情況Fig.1 Cold protease transparent zone formed by strains
在不同培養溫度下,分別對上述3 株產蛋白酶菌株進行液體發酵培養,菌株11815 在4~37,℃均可生長,其最適生長溫度為20,℃左右,當溫度大于等于40,℃時,則生長極為緩慢甚至停止生長;菌株11813及11816 的最適生長溫度與菌株11815 相似,因此均屬于典型的耐冷菌.由于菌株11815 的產蛋白酶能力較強,因此作為下一步重點研究對象.
2.2 菌株11815的分析鑒定
菌株11815 屬革蘭氏陽性菌,呈球形或卵圓形,直徑約1.0~1.2,μm,呈雙球狀或連珠狀排列;菌落形態圓形,顏色橙黃色,表面光滑、邊緣齊整.該菌屬好氧性或兼性厭氧菌,可較好利用葡萄糖、果糖、淀粉等碳源,能夠利用硫酸銨、蛋白胨、水解酪蛋白等氮源;具有氧化酶、過氧化氫酶活性,明膠液化實驗呈陽性.
該菌16S rDNA 全長序列為1,450,bp,與動性球菌(Planococcus antarcticus) DSM 14505 的相似度為99%,結合形態特征和生理生化特性,將其鑒定為動性球菌屬.
2.3 溫度和pH對菌株11815產酶的影響
在搖瓶發酵條件下,研究不同培養溫度(15、20、25,℃)對菌株11815 產酶的影響,結果如圖2 所示.結果表明:該菌株的最佳生長溫度與最適產酶溫度相一致,均為20,℃,發酵培養48,h 可達到產酶高峰,產酶量約為280,U/mL.在菌體培養的初級階段,菌體的生長量與產酶量呈正比;當菌體生長達到平衡期后,菌體量不再增加,而粗酶液中的蛋白酶活性則開始下降,這可能與蛋白酶的自身降解有關.從上述實驗結果推斷:該蛋白酶的表達應為組成型表達,只與菌體量有關,而與其他誘導因素無關.因此在發酵過程中設法提高菌體濃度,同時在發酵液中加入適量蛋白酶抑制劑,可有效提高蛋白酶產量.

圖2 培養溫度對產酶的影響Fig.2 Effect of culture temperature on enzyme production
pH 對菌株產酶的影響較小,初始pH 在6~8 之間,菌株11815 均可穩定生長并產酶.
2.4 酶學性質分析
2.4.1 酶的最適作用溫度及熱穩定性檢測
在不同溫度下分別對菌株11815 發酵液中的蛋白酶活性進行了檢測,結果如圖3 所示.該菌株所產蛋白酶的最適作用溫度為37,℃左右,并且在25~30,℃均有較好的催化活性(最高催化酶活的50%以上),與其他低溫蛋白比較發現,蛋白酶SKPB5 以及焦曲霉(Aspergillus ustus)、假單胞菌(Pseudomonas)strain DY-A 分泌的蛋白酶等,最適溫度在 40~42,℃[8–10],11815 分泌的蛋白酶的最適作用溫度更低.因此,該菌株所產蛋白酶的最適作用溫度與自然環境中的溫度基本一致,在使用過程中可以省去加熱及冷卻的過程,具有節能環保等應用屬性.
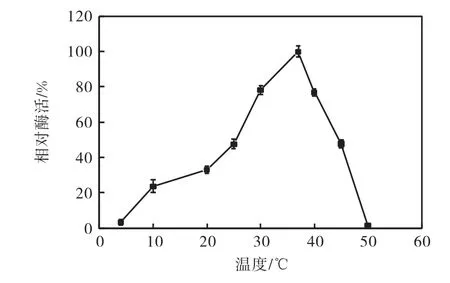
圖3 蛋白酶的作用溫度與相對酶活的關系Fig.3 The relationship between protease temperature and the relative enzyme activity
酶的熱穩定性實驗結果如圖4 所示.該菌株所產蛋白酶的熱穩定性較低,37,℃保溫 20,min 或45,℃保溫60,min 后,酶活僅剩原來的10%;50,℃保溫10,min 后,則催化活性完全喪失;該蛋白酶只有在10,℃以下才能保存較長的時間;這一特性與大多數低溫蛋白酶[7–9]相似,主要與低溫蛋白酶本身較為松散的蛋白結構及其較高的分子柔性有關.
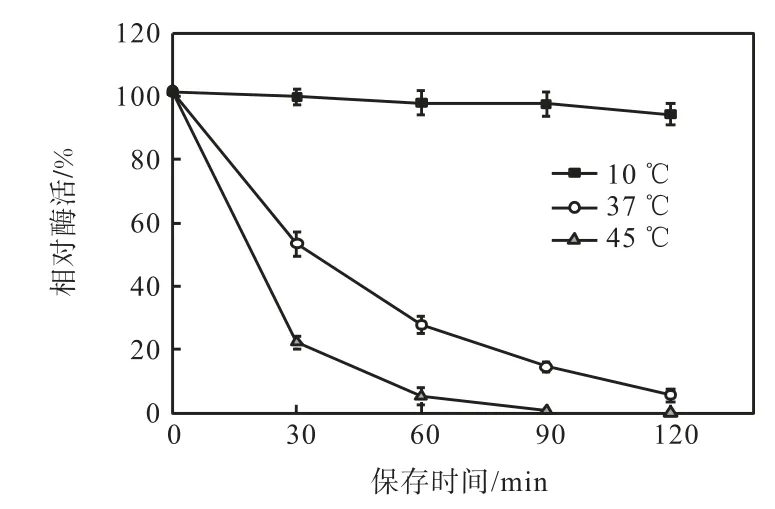
圖4 蛋白酶的熱穩定性Fig.4 Effects of temperature on the stability of protease activity
2.4.2 酶的最適作用pH
分別在不同pH 條件下測定菌株11815 所產蛋白酶活力,酶活力隨pH 變化的曲線如圖5 所示.該酶在pH 4.0~10.0 均具有催化活力,其最適作用pH為8.0 左右;當pH 升至9.0 時,其酶活約為最高酶活的70%左右,當pH 升至10.0 時,酶活大幅下降,僅為最高酶活的25%;當pH 降至7.0 時,酶活力約為最高活力的82%.因此,該菌株所產蛋白酶為低溫蛋白酶.
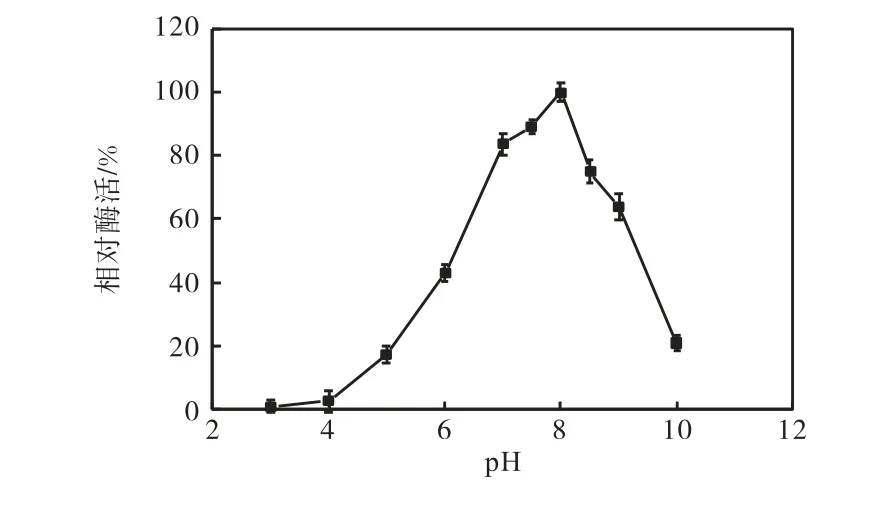
圖5 蛋白酶的作用pH與相對酶活的關系Fig.5 The relationship between pH of protease and the relative enzyme activity
2.4.3 不同金屬離子及化學試劑對蛋白酶酶活的影響
不同金屬離子及化學試劑對蛋白酶酶活的影響見表1.
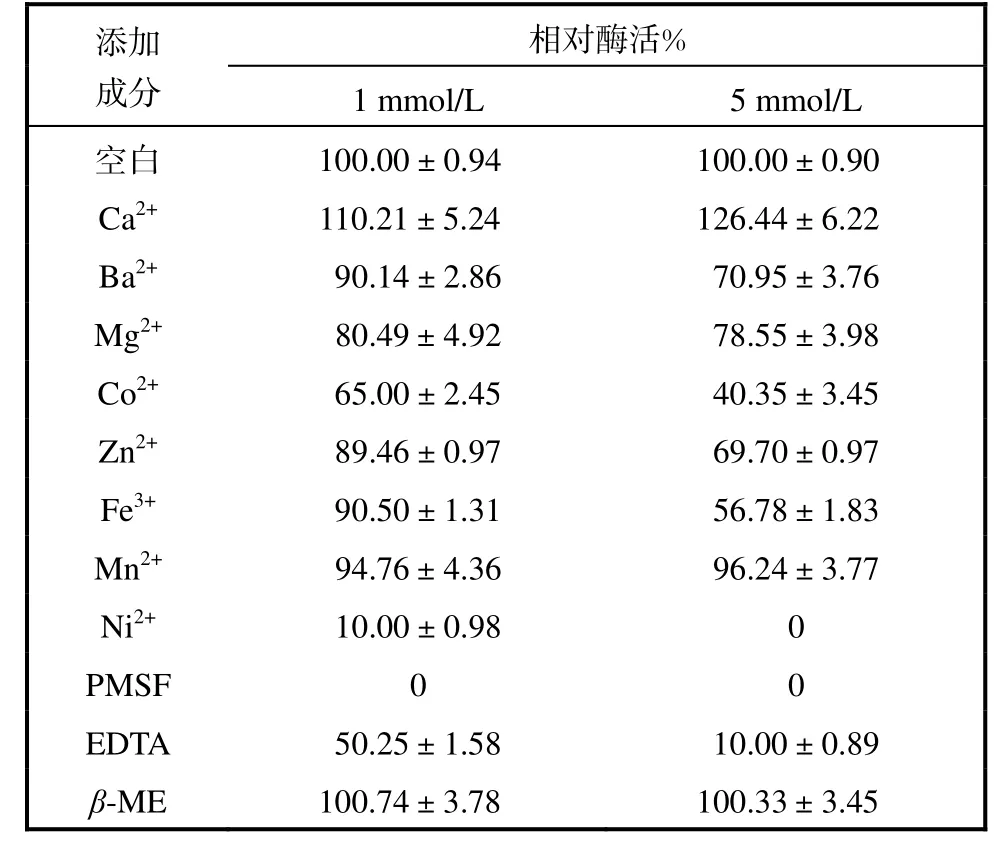
表1 不同的金屬離子、抑制劑對蛋白酶酶活的影響Tab.1 Effect of various metal ions and inhibitors on protease activity
金屬離子Ca2+對該菌株所產蛋白酶有激活作用,Mn2+對其基本沒有影響;Mg2+、Ba2+、Fe3+、Zn2+、Co2+、Ni2+則對其有一定的抑制作用,Ni2+對其抑制作用最強,濃度為1.0,mmol/L 的Ni2+可抑制90%以上的酶活,因此組氨酸可能是該酶的活性中心之一;EDTA 對該酶有一定抑制作用,某些金屬離子可能對維持酶分子的三維結構有重要作用;另外,該酶被PMSF 強烈抑制,因此該菌株所產蛋白酶的活性位點應包括His、Ser 以及羧基氨基酸.
3 結語
從南印度洋深海沉積物中分離得到1 株產低溫蛋白酶菌株,經菌種鑒定為動性球菌(Planococcus sp.).該菌株的最適生長溫度為 20,℃左右,可在10~25,℃進行快速生長代謝;并在20,℃以下具有表達分泌低溫蛋白酶的能力,最適的培養溫度為20,℃,搖瓶發酵條件下,48,h 內的蛋白酶產量可達280,U/mL.酶學性質分析結果顯示:該菌株所產蛋白酶的最適作用pH 為8.0,最適作用溫度為37,℃,當催化溫度降至30,℃時,酶活可保持在80%以上,并且在4,℃條件下仍具有水解蛋白的能力.EDTA 對該酶有一定抑制作用,PMSF 對該酶具有強烈抑制作用,因此該菌株所產蛋白酶的活性位點應包括His、Ser 以及羧基氨基酸.該酶熱穩定性較差,50,℃僅保溫1.0,min,則酶活完全喪失.由該菌株所產蛋白酶的上述特性表明,該酶屬于典型的低溫蛋白酶,具有一定的研究及開發價值.本課題組還將對該低溫蛋白酶的編碼基因進行分離與克隆,并進一步嘗試在其他宿主菌中實現其高效異源表達.
[1]Joshi S,Satyanarayana T.Biotechnology of cold-active proteases[J].Biology,2013,2(2):755–783.
[2]Pawar R,Zambare V,Barve S,et al.Application of protease isolated from Bacillus sp.158 in enzymatic cleansing of contact lenses[J].Biotechnology,2009,8(2):276–280.
[3]Kuddus M,Ramteke P W.Recent developments in production and biotechnological applications of cold-active microbial proteases[J].Critical Reviews in Microbiology,2012,38(4):330–338.
[4]林永成,周世寧,樂長高.海洋微生物活性代謝產物化學[J].大學化學,1996,11(6):1–7.
[5]薩姆布魯克 J,拉塞爾D W.分子克隆實驗指南[M].黃培堂,譯.3 版.北京:科學出版社,2002.
[6]東秀珠,蔡妙英.常見細菌鑒定系統手冊[M].北京:科學出版社,2001.
[7]張樹政.酶制劑工業:下冊 [M].北京:科學出版社,1984:446–447.
[8]Kasana R C,Yadav S K.Isolation of a psychrotrophic Exiguobacterium sp.SKPB5(MTCC 7803)and characterization of its alkaline protease[J].Current Microbiology,2007,54(3):224–229.
[9]Damare S,Raghukumar C,Muraleedharan U D,et al.Deep-sea fungi as a source of alkaline and cold-tolerant proteases[J].Enzyme and Microbial Technology,2006,39(2):172–181.
[10]Zeng R,Zhang R,Zhao J,et al.Cold-active serine alkaline protease from the psychrophilic bacterium Pseudomonas strain DY-A:Enzyme purification and characterization[J].Extremophiles,2003,7(4):335–337.
