不同腎上腺能受體激動劑對冷暴露中緬樹鼩產熱能力的影響
2015-01-16 07:54:31朱萬龍王政昆
綠色科技 2015年12期
朱萬龍,王政昆
(云南省高校西南山地生態系統動植物生態適應進化及保護重點實驗室/云南師范大學 生命科學學院,云南 昆明 650500)
不同腎上腺能受體激動劑對冷暴露中緬樹鼩產熱能力的影響
朱萬龍,王政昆
(云南省高校西南山地生態系統動植物生態適應進化及保護重點實驗室/云南師范大學 生命科學學院,云南 昆明 650500)
研究了冷馴化條件下不同腎上腺能受體激動劑對中緬樹鼩產熱能力的影響,測定了其靜止代謝率和非顫抖性產熱。結果表明:冷馴化組中緬樹鼩的靜止代謝率(Resting metabolic rate,RMR)較對照組增加,β3-NST(注射β3-腎上腺素激動劑BRL37344)和NST(注射NE)較對照組均極顯著增加。冷馴化組和對照組在注射NE和BRL37344后其耗氧量均有極顯著的增加。對照組中緬樹鼩的NST高于β3-NST,但兩者之間差異不顯著。冷馴化組中緬樹鼩NST與β3-NST差異極顯著。以上結果說明冷馴化可使中緬樹鼩的產熱能力顯著增強,其BAT中可能存在有β3-腎上腺能受體(β3-adrenergic receptor,β3-AR),推測β3-AR可能并不是中緬樹鼩BAT中唯一的腎上腺能受體,NE可能通過多種腎上腺能途徑來增加產熱,腎上腺能受體通路在中緬樹鼩適應性產熱中起到了重要作用。
中緬樹鼩;腎上腺能激素;產熱
1 引言
低溫脅迫是刺激小型哺乳動物產熱能力增加的主要環境因子之一[1]。低溫條件下小型哺乳動物往往會增加其靜止代謝率(Resting metabolic rate, RMR)和非顫抖性產熱(Nonshivering thermogenesis, NST)[2]來適應這樣的環境。低溫脅迫刺激靜止代謝率和非顫抖性產熱增加的產熱機理是不同的,刺激靜止代謝率的增加主要來自內臟器官產熱能力的增加,如肝臟[3],非顫抖性產熱的增加則主要是通過褐色脂肪組織(Brown adipose tissue, BAT)中解偶聯蛋白(Uncouping protein, UCP)的數量和活性的增加,從而使得非顫抖性產熱增加[4]。
靜止代謝率是動物維持正常生存的最低代謝水平[5],能反映不同種群或物種的能量消耗水平,對動物適應環境的理解具有重要的意義[6]。動物主要是通過交感神經釋放去甲腎上腺素(norepinephrine, NE)來參與非顫抖性產熱的調節,之前的研究認為非顫抖性產熱是腎上腺能受體刺激的產熱[7],非顫抖性產熱對于小型哺乳動物適應低溫環境具有重要的意義[8]。β3-腎上腺能受體(β3-adrenergic receptor, β3-AR)特異性激動劑BRL37344能有效激活BAT產熱系統,導致非顫抖性產熱產熱顯著增加[9]。中緬樹鼩(Tupaiabelangeri)屬攀鼩目(Scandentia)樹鼩科(Tupaiidae),為東洋界特有的小型哺乳動物,我國云貴高原及其附近的橫斷山區可能是中緬樹鼩的分布的北限[10]。對中緬樹鼩冷馴化條件下的RMR和NST的研究有助于了解中緬樹鼩在低溫脅迫條件下的適應對策,從而進一步闡述該動物對環境變化的適應性。
2 材料和方法
2.1 動物來源
實驗動物捕自冬季大理青光山,動物滅蚤后,帶回云南師范大學動物飼養房飼養,單籠飼養(300 mm×200 mm×120 mm),無巢材,光照條件12L:12D,每日喂以標準飼料和加少許水果和面包蟲[11],塑料瓶供水,自由取食和進水。實驗用中緬樹鼩均為成年非繁殖期個體。實驗前適應30 d后開始實驗,實驗分為3組:未注射組、注射β3-腎上腺素激動劑組和注射NE組。實驗0 d測定3組的RMR、β3-NST和NST,然后將3組動物放入馴化房,進行冷馴化實驗,溫度為5±1 ℃,光照條件12L:12D,馴化28 d。28 d后,再次測定RMR、β3-NST和NST。實驗前3組實驗動物的體重差異不顯著(P>0.05)。
2.2 RMR測定
用開放式呼吸儀(ADML870型,澳大利亞生產)測定動物的RMR,具體的測定方法參照Zhu[12],代謝率的計算方法參照Hill[13]。
2.3 NST測定
皮下注射去甲腎上腺素(norepinephrine, NE),測定NST。誘導NST的NE劑量為0.8 mg/kg,NST測定時間為30 min,期間出現的最大持續耗氧量視為動物的NST[11]。
2.4 β3-NST的測定
肩腳部位皮下注射β3腎上腺素激動劑BRL37344,測定其呼吸率的變化情況。注射劑量經預備實驗確定為0.4 mg/kg。以注射BRL37344后的最大持續耗氧量為動物的β3-NST。
2.5 數據處理
采用SPSS16.0軟件包進行實驗數據的統計。數據經過正態分布和方差齊性檢驗,符合參數檢驗條件。馴化前、馴化后動物的BMR、β3-NST和NST的差異采用單因素方差分析(one-way ANOVA)檢驗,并采用Duncan多重比較。組間同一指標的差異采用獨立樣本t檢驗。結果均以平均值±標準誤(Mean±SE)表示,P<0.05為差異顯著,P<0.01為差異極顯著。
3 結果分析
實驗后,對照組中緬樹鼩的RMR較0 d時增加,但是兩者之間差異不顯著(t=-0.752,P>0.05, 表1)。與對照組相比較,無論是馴化0 d還是馴化后28 d,β3-NST和NST較對照組(0 d)均極顯著的增加(馴化0 d:β3-NST,t=-1.635,P<0.01;NST,t=-1.986,P<0.01;馴化28 d:β3-NST,t=-2.356,P<0.01;NST,t=-2.871,P<0.01,圖1)。馴化0 d時NST組的產熱能力高于β3-NST組,但是兩者之間差異不顯著(t=-0.236,P>0.05)。冷馴化28 d后中緬樹鼩NST組的產熱能力高于β3-NST組,且兩者之間差異極顯著(t=1.843,P<0.01)。
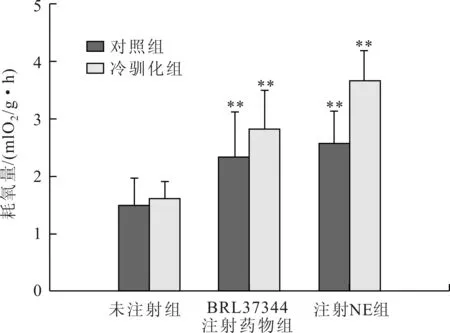
圖1 不同腎上腺能受體激動劑對中緬樹鼩產熱能力的影響(與對照組比較)

表1 不同腎上腺能受體激動劑對中緬樹鼩產熱能力的影響
4 討論
本研究組先前的研究表明:隨著中緬樹鼩棲息地緯度和海拔高度的增加,中緬樹鼩RMR和NST出現明顯的季節性變化[14],低溫[15]和短照[16]脅迫可以顯著刺激產熱能力的增加,而且在低溫馴化條件下中緬樹鼩的RMR增加的比例大于NST增加[17],本研究在此基礎上,對冷馴化條件下不同腎上腺能受體激動劑對中緬樹鼩產熱能力的影響進行了研究。
產熱能力的增加是小型哺乳動物應對低溫脅迫的一種重要生理適應對策[1]。NST產熱能力的大小是決定動物抵抗低溫能力的主要影響因子之一[4],具有重要的生物學意義[5]。低溫暴露可使小型哺乳動物的NST顯著增加[7]。BRL37344是β3-AR受體特異性激動劑,而NE是一種非特異性的腎上腺能受體激動劑,兩者都可誘導NST產熱[18]。本文的實驗結果表明,冷馴化后中緬樹鼩的RMR,β3-NST和NST均比對照組(未冷馴化)增加,表明低溫可使中緬樹鼩的產熱能力顯著增強,這與本研究組之前的實驗結果一致[15]。這與其他一些小型哺乳動物在低溫條件下的適應策略相似[19]。
BAT的產熱主要是由β3-AR介導,研究表明低濃度的NE也能刺激BAT的產熱反應[20]。本研究結果表明,與對照組相比較,無論是馴化前還是馴化28 d后,注射NE后其NST增加極顯著,可能說明β3-AR在中緬樹鼩NST產熱過程中的非常重要的作用。對布氏田鼠的研究也表明,BAT的β-AR在低溫條件下也起到重要的作用[21]。另外,注射BRL37344組不論無論是馴化前還是馴化28 d后其NST誘導的產熱顯著增加,說明這兩種藥物的作用是相互獨立的。這一特征與其他實驗嚙齒動物相似[21],如金黃倉鼠[22]。此外,本研究還發現,無論是馴化0 d還是冷馴化28 d,在注射BRL37344后,與未注射的對照組相比較,其NST產熱增加極顯著,這可能說明β3-AR確實能使中緬樹鼩產熱能力顯著增加,中緬樹鼩BAT中存在有β3-AR。已有研究表明BAT中存在有兩種腎上腺能受體:α-AR和β-AR,其中β-AR的三種亞型,即β1-AR、β2-AR和β3-AR[22]。因為NE是一種非特異性的腎上腺能受體激動劑,它可以刺激多種腎上腺能受體,并誘導相應的生理反應,而本研究中β3-AR誘導中緬樹鼩的NST顯著低于NE誘導產熱,這可能說明β3-AR并不是中緬樹鼩BAT中唯一的腎上腺能受體。
冷馴化可使中緬樹鼩的產熱能力顯著增強,其BAT中可能存在有β3-AR,并且β3-AR可能并不是BAT中唯一的腎上腺能受體,腎上腺能受體通路在中緬樹韻適應性產熱中起到了重要作用。
[1]Zhao Z J, Cao J, Liu Z C,et al. Seasonal regulations of resting metabolic rate and thermogenesis in striped hamster (Cricetulus barabensis) [J]. Therm Biol,2010,35(1):401~405.
[2]Zhu W L, Wang Z K. Seasonal changes in body mass, serum leptin levels and hypothalamic neuropeptide gene expression in male Eothenomys olitor[J]. Comparative Biochemistry and Physiology,2015,184(1):83~89
[3]Zhu W L, Cai J H, Lian X, et al. Adaptive character of metabolism in Eothenomys miletus in Hengduan Mountains region during cold acclimation[J]. Journal of Thermal Biology,2010,35(8):417~421.
[4]Zhang Z Q, Wang D H. Seasonal changes in thermogenesis and body mass in wild Mongolian gerbils (Meriones unguiculatus)[J]. Comp Biochem Physiol, 2007(148):346~353.
[5]MaNab B K. On the utility of uniformity in the definition of basal rate of metabolism[J]. Physiol Zool,1997(70):718~720.
[6]Bozinovic F, Gallardo P. The water economy of South American desert rodents: from integrative to molecular physiological ecology[J]. Comp Biochem Physiol,2006(142):163~172.
[7]Speakman J R. The energy cost of reproduction in small rodents[J]. Acta Ther Sin,2007(27):1~13.
[8]Tang G B, Cui J G, Wang D H. Role of hypoleptinemia during cold adaptation in Brandt's voles (Lasiopodomys brandtii)[J]. Am J Physiol Regul Integr Comp Physiol, 2009,297(5):1293~1301.
[9]Strosberg A D. Strueture and function of the adrenergic receptor[J]. Annu Rev Pharmacol Toxicol,1997(37):421~450.
[10]Sloan Wilson D, Clark A B, Coleman K. Shyness and boldness in humans and other animals[J]. Trends Ecol Evol,1994,9(11):442~446.
[11]Zhu W L, Jia T, Huang C M, et al. Changes of energy metabolism, thermogenesis and body mass in the tree shrew (Tupaia belangeri chinensis) during cold exposure[J]. Italian Journal of Zoology,2012,79(2):175~181.
[12]Zhu W L, Mu Y, Liu J H, et al. Energy requirements during lactation in female Apodemus chevrieri (Mammalia: Rodentia: Muridae) in Hengduan mountain region[J]. Italian Journal of Zoology,2015,82(2):165~171.
[13]Hill R W. Determination of oxygen consumption by use of the paramagnetic oxygen analyser[J]. Appl Physiol,1972(33):261~263.
[14]Zhu W L, Zhang H, Wang Z K. Seasonal changes in body mass and thermogenesis in tree shrews (Tupaia belangeri) the roles of photoperiod and cold[J]. Journal of Thermal Biology,2012(37):479~484.
[15]Zhang L, Zhu W L, Wang Z K. Role of photoperiod on hormone concentrations and adaptive capacity in tree shrews, Tupaia belangeri[J]. Comparative Biochemistry and Physiology,2012(163):253~259.
[16]Zhang L, Zhang H, Zhu W L, et al. Energy metabolism, thermogenesis and body mass regulation in tree shrew (Tupaia belangeri) during subsequent cold and warm acclimation[J]. Comparative Biochemistry and Physiology,2012(162):437~442.
[17]Zhang L, Liu P F, Zhu W L, et al. Variations in thermal physiology and energetics of the tree shrew (Tupaia belangeri) in response to cold acclimation[J]. Journal of Comparative Physiology,2012(182):167~176.
[18]Heldmaier D. Source if heat during nonshivering thermogenesis in Djungarian hamster[J]. Comp Physiol,1985(156):237~245.
[19]王政昆,李慶芬,孫儒泳.褐色脂肪組織產熱及其調節機理[J].生理科學進展,27(4):353~355.
[20]Nedergaard J, Valeria G, Anita M, et al. UCP1: the only protein able to mediate adaptive non-shivering thermogenesis and metabolic inefficiency[J]. Biochimica et Biophysica Acta, 2001(1504):82~106.
[21]李慶芬,劉小團,黃晨西.長爪沙鼠冷馴化過程眾褐色脂肪組織產熱活性及解偶聯蛋白基因表達[J].動物學報,2001,47(4):388~393.
[22]Zhao J, Cannon B, Nedergaard J. Thermogenes is β3-but not βl-adrenergically mediated in rat brown fat cells, even after cold acclimation[J]. Am J Physiol,1998,275(6):2002~2011.
Study on the Effects of Different Adrenergic Agonistson the Thermogenic Capacity of Tupaia BelangeriUnder Cold Exposure
Zhu Wanlong, Wang Zhengkun
(KeyLaboratoryofEcologicalAdaptiveEvolutionandConservationonAnimals-PlantsinSouthwestMountainEcosystemofYunnanProvinceHigherInstitutesCollege,SchoolofLifeSciences,YunnanNormalUniversity,Kunming650500,China)
his article mainly studies oneffect of different β-AR agonists on the thermogenic capacity ofTupaia belangeri under the condition of cold exposure and tests Tupaia belangeri's resting metabolic rate (RMR) and nonshivering thermogenesis (NST). The results show that RMR increasesin the group ofcold exposure compared with control group (out of cold exposure).But β3-NST and NST increase significantly compared with control group. After the injection of NE and BRL37344, the oxygen consumption increases obviously in both groups. NST is higher thanβ3-NST in control group, but the differences are not obvious. In the group of cold exposure, the differences between NST and β3-NST are obvious. All of results indicate that the cold exposure makes theTupaia belangeri's thermogenic capacity stronger and there exists β3-AR in BAT. The article speculates thatβ3-AR is not the only AR in BAT. Perhaps, and NE may enhance thermogenesis by many adrenergic approaches, which may play an important role in the adaptive thermogenesis in T. belangeri.
Tupaia belangeri; adrenaline; thermogenesis
2015-09-30
國家國際科技合作項目(編號:2014DFR31040) ;十二五國家支撐計劃(編號:2014BAI01B00);國家自然科學基金項目(編號:31360096)資助
朱萬龍(1983—),男,江蘇南京人,博士,副教授,主要從事動物生理生態學研究。
Q953
A
1674-9944(2015)12-0001-03
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
發明與創新(2022年30期)2022-10-03 08:40:56
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
動漫星空(興趣百科)(2020年12期)2020-12-12 05:31:40
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
人大建設(2018年6期)2018-08-16 07:23:10
文理導航·科普童話(2017年5期)2018-02-10 19:42:14
無人機(2017年10期)2017-07-06 03:04:36
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
