瀘寧雞和米易雞MSTN基因多態性及其與生長性狀的關聯性
2015-02-27 12:31:04任冰冰蔣小松熊應龍王康環劉光偉徐亞歐
貴州農業科學 2015年7期
左 斌, 任冰冰, 蔣小松, 熊應龍, 王康環,蔣 利, 劉光偉 , 王 海, 徐亞歐*
(1.西南民族大學 生命科學與技術學院, 四川 成都 610041; 2.四川省畜牧科學研究院, 四川 成都 610066; 3.涼山州原生農業綜合開發有限責任公司, 四川 冕寧 615600)
?
瀘寧雞和米易雞MSTN基因多態性及其與生長性狀的關聯性
左 斌1, 任冰冰1, 蔣小松2, 熊應龍3, 王康環1,蔣 利1, 劉光偉1, 王 海1, 徐亞歐1*
(1.西南民族大學 生命科學與技術學院, 四川 成都 610041; 2.四川省畜牧科學研究院, 四川 成都 610066; 3.涼山州原生農業綜合開發有限責任公司, 四川 冕寧 615600)
為探明瀘寧雞、米易雞肌肉生長抑制素(myostatin,MSTN)基因多態性與生長性狀的關聯性,采用PCR-SSCP法對MSTN基因的多態性進行檢測,同時對瀘寧雞、米易雞的主要生長性狀進行測定,并對MSTN基因多態性與生長性狀的關聯性進行分析。結果表明:MSTN基因外顯子1中有2 100 bp(C/T)和2 109 bp(C/T)2個SNPs位點,未引起氨基酸變異,為同義突變,表現出CT、TT(2 100 bp)和CC、CT、TT(2 109 bp)5種基因型,與心臟重、脛圍和屠宰率有相關性;瀘寧雞存在CC、CT和TT 3種單倍型,米易雞存在CC、TC和TT 3種單倍型。內含子1中有4 295 bp(T/C)、4 359 bp(T/C)和4 404 bp(A/C)3個SNPs位點,表現出CT(4 295 bp)、CT(4 359 bp)、TT、CT和CC(4 404 bp)5種基因型,與腺胃重有顯著相關;瀘寧雞存在CCC和TTT 2種單倍型,米易雞存在CCC、TTC和TTT 3種單倍型。3’非編碼區有7 637 bp(A/G)和7 725 bp(A/G)2個SNPs位點,表現出AG(7 637 bp)、AA、AG和GG(7 725 bp)4種基因型,與腺胃重、腹脂重和半凈膛率具有相關性;瀘寧雞存在AG、GA和GG 3種單倍型。MSTN基因是影響瀘寧雞、米易雞主要生長性狀的主效基因之一,可以作為地方雞品種選育的候選基因。
瀘寧雞; 米易雞; 肌肉生長抑制素; PCR-SSCP; 生長性狀; 單倍型
肌肉生長抑制素(MSTN)基因是1997年MePherron等從小鼠骨骼肌cDNA文庫中克隆出的一個新基因,該基因轉錄合成的蛋白質是一種生長因子,被命名為生長分化因子-8(Growth and Differential Factor-8,GDF-8)[1-2]。該基因的缺失和突變會使一些動物表現出明顯的雙肌現象[3]。在畜牧業上,篩選出MSTN基因突變個體,可通過育種培育出產率高的優良畜禽品種。胡蘭等[4]研究表明,在大骨雞胸肌、腿肌、心肌、腎臟、腦、舌中均有MSTN基因表達,其中骨骼肌中表達水平較高。顧志良等[5]對明星肉雞和絲羽烏骨雞雜交產生的F2代群體進行研究發現,MSTN基因核苷酸突變(G→A,A→G,C→T)導致其5′調控區呈現出多態性,G→A和A→T的突變造成MSTN 3′調控區的多態性,MSTN基因多態性不僅影響骨骼肌和肌肉生長發育,還可能與脂肪沉積有關。隱性白羽雞在MSTN基因5′調控區和3′調控區存在SNPs位點[6]。到目前為止,未見MSTN基因多態性在四川優良地方雞品種瀘寧雞、米易雞上的研究報道。為探明四川優良地方雞品種MSTN基因多態性及其與生長性狀的關聯性,找出該基因與生長性狀相關的遺傳標記,運用PCR-SSCP技術篩選MSTN基因SNPs變異位點,分析其與生長性狀的關系,以期為培育出生長速度快的優良地方雞品種提供理論依據。
1 材料與方法
1.1 雞品種
瀘寧雞(64只)、米易雞(64只)取自涼山州冕寧縣瀘沽鎮秧草壩養殖場,由專人管理,單籠飼養,管理和營養水平一致,自由采食、飲水。分別在81 d、119 d、154 d、210 d進行屠宰性能測定,同時用真空采血管收集血樣EDTA.K2抗凝帶回實驗室,于-80℃保存備用。
1.2 生長性狀測定
屠宰前測定體斜長、胸寬、胸深、胸骨長、盆骨寬、脛圍和脛長等指標。屠宰性能測定參照楊寧[7]的方法進行,指標包括活重、屠體重、全凈膛重、半凈膛重、胸肌重、腿肌重、腹脂重、肝重、肌胃和腺胃重,半凈膛率、全凈膛率、腹脂率、胸肌率、腿肌率等。
1.3 引物設計及DNA池構建
根據GenBank登錄的MSTN基因序列(登錄號:AF346599)用Primer5.0軟件設計特異性引物(表1),并由上海Invitrogen公司合成,擴增包含該基因3個外顯子序列及含有SNPs位點的特異性序列。用DNA試劑盒提取血液基因組DNA,經TE溶解后,-20℃保存備用。每個品種隨機抽取30個DNA樣品,用紫外分光光度計測量每個DNA樣品濃度各3次,取平均值,將樣品稀釋到終濃度為100 ng/μL,從30個樣品中各取5 μL混合構建成DNA池[8-10]。

表1 試驗所用引物信息
1.4 PCR擴增
以DNA池為模板擴增MSTN基因3個外顯子片段。PCR反應體系(25 μL):超純水9.5 μL,上下游引物(10 μmol/L)各1 μL,DNA模板(100 ng/μL)1 μL,Master Mix 12.5 μL。PCR反應程序:94℃預變性3 min;94℃變性30 s,復性30 s,72℃延伸30 s,30個循環;72℃延伸10 min,4℃保存。1.5%瓊脂糖凝膠電泳檢測PCR產物的亮度及純度。
1.5 SSCP分析及測序
取PCR產物1.5 μL與變性緩沖液(10%蔗糖,0.01%溴酚,0.01%二甲精)20 μL混合,97℃變性3 min后立即冰浴5 min,產物經12%非變性聚丙烯酰胺凝膠[m(Arc)∶m(Bis)=29∶1]在200 V、400 mA條件下電泳3~4 h后銀染顯色。經PCR-SSCP[11-13]分析后,挑選每個不同基因型個體的PCR產物送上海Invitrogen公司測序。用DNAMAN軟件將測序結果與GenBank登陸的序列進行比對分析,找出突變位點。
1.6 統計分析
根據電泳結果不同帶型的差異,判斷個體的基因型,用EXCEL軟件進行數據統計和SPSS17.0軟件進行差異顯著性比較,進行基因型與生長性狀間的關聯分析。利用POPGENE軟件計算各等位基因在各品種中的基因頻率和基因型頻率,用卡方值判斷雞群是否處于哈代-溫伯格平衡,用Nei氏計算法檢測MSTN基因雜合度、純合度和多態信息含量,用SHEsis對SNPs位點進行單倍型構建并計算其頻率。
2 結果與分析
2.1 PCR擴增、SSCP檢測及測序
針對MSTN基因外顯子1,內含子1及3’非編碼區設計3對引物的擴增片段與預期大小一致,沒有非特異性條帶(圖1),可以用于進一步分析。經SSCP分析(圖2)發現,MSTN基因外顯子1變異位點表現出5種帶型,分別命名為TT、CT(2 100 bp)和CC、CT、TT(2 109 bp);內含子1變異位點表現出CT(4 295 bp),CT(4 359 bp),TT、CT、CC(4 404 bp) 5種基因型;3′非編碼區變異位點表現出AG(7 637 bp)和AA、AG、GG(7 725 bp) 4種基因型。經測序后序列比對發現,外顯子1有2個SNP位點,分別為2 100 bp(C/T)和2 109 bp(C/T);內含子1有3個SNP位點,分別為4 295 bp(T/C)、4 359 bp(T/C)和4 404 bp(A/C);3′非編碼區有2個SNP位點,為7 637 bp(A/G)和7 725 bp(A/G)。

注:M為Marker D2000,1~3為外顯子1突變位點,4~6為內含子1突變位點,7~9為3′非編碼區突變位點。
Note: M, Marker D2000; 1~3, Mutation loci of exon 1; 4~6, Mutaion loci of intron 1; 7~9, Mutation loci of 3′ noncoding region(A7637G and A7725G).
圖1MSTN基因的PCR產物電泳圖譜
Fig.1 Electrophoretoesis pattern of PCR product ofMSTNgene
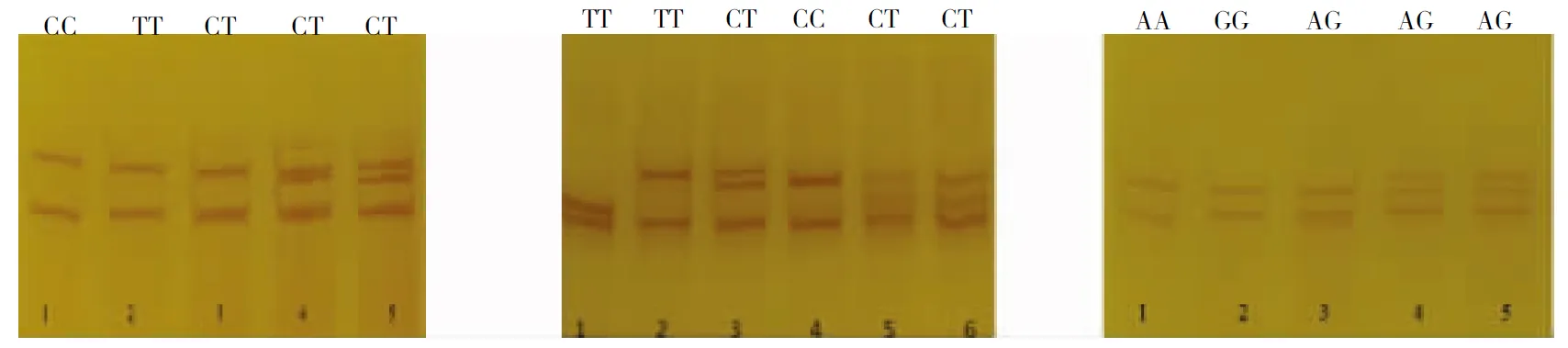
圖2MSTN基因的SNPs分析及基因型(從左到右是外顯子1,內含子1,3′非編碼區)
Fig.2 SNPs analysis and genotype ofMSTNgene (From left to right are exon 1, intron 1 and 3′ noncoding region)
2.2 基因型頻率和基因頻率
從表2可知,MSTN基因外顯子1中瀘寧雞的2個位點均是CC基因型頻率、C等位基因頻率最高;米易雞2 109 bp位點CT基因型、C等位基因頻率最高,2 100 bp位點T為優勢等位基因。內含子1中瀘寧雞的3個位點均是TT基因型頻率最高,T為優勢等位基因;米易雞4 295 bp、4 359 bp位點是CT基因型頻率最高,4 404 bp位點CC基因型頻率最高,4 295 bp、4 359 bp位點T為優勢等位基因,4 404 bp位點C為優勢等位基因。3′非編碼區瀘寧雞7 637 bp位點GG基因型頻率最高,G為優勢等位基因,米易雞在該位點未發生突變;7 725 bp位點GG基因型頻率均最高,G為瀘寧雞和米易雞的優勢等位基因。經卡方適合性檢驗,瀘寧雞在各個變異位點均處于Hardy-Weinberg平衡,米易雞在2 109 bp、4 295 bp和4 359 bp位點均極顯著偏離Hardy-Weinberg平衡。
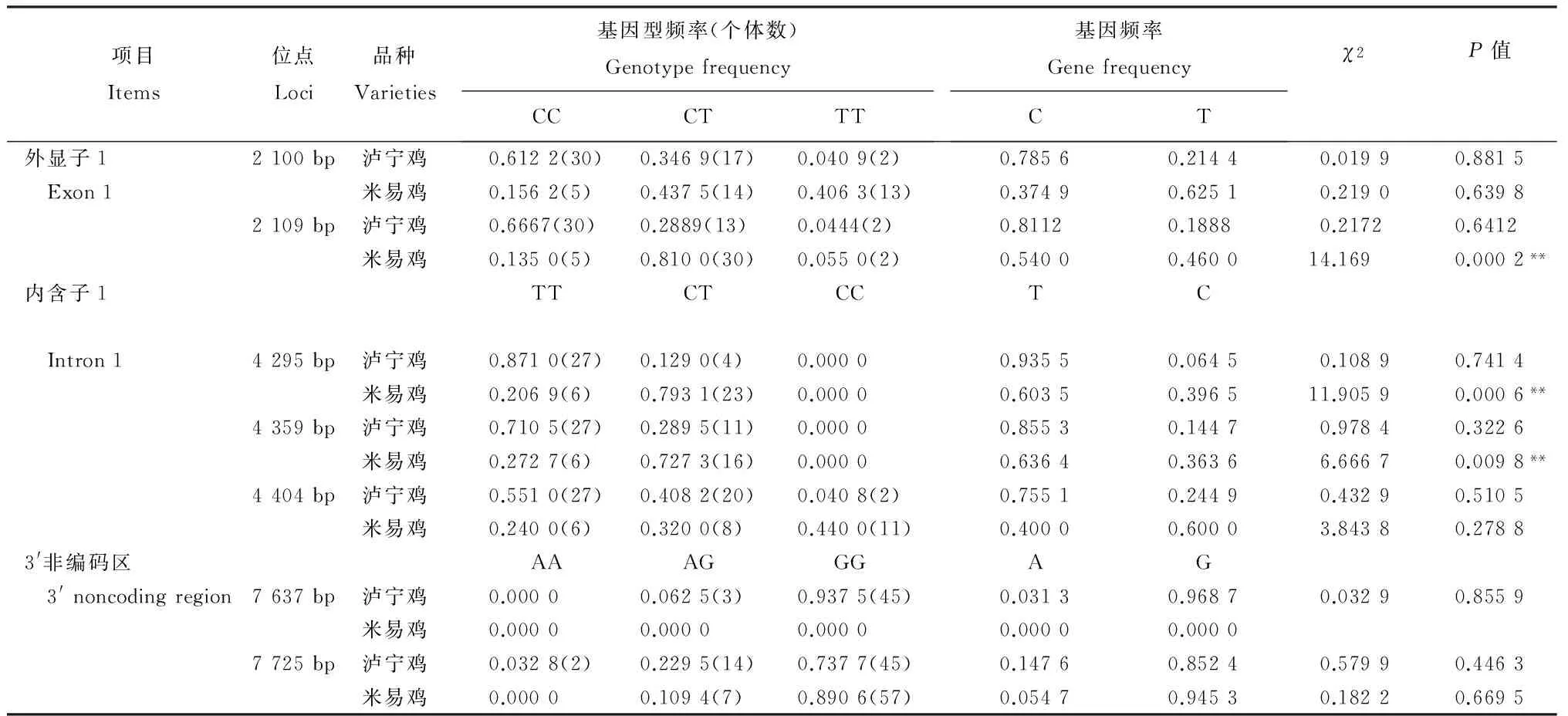
表2 MSTN基因的基因型頻率和基因頻率及χ2 檢驗
2.3 基因雜合度和多態信息含量
一般認為,基因雜合度(Ho)和多態信息含量(PIC)是度量各群體在位點上遺傳變異的最適參數,其數值大小反映出遺傳結構變異程度的高低,數值越低,表明群體的遺傳多樣性越低。從表3看出:各位點的Ho在0~0.496 7。在2 109 bp、4 359 bp和7 725 bp位點,瀘寧雞的PIC在0.25~0.5,表現出中度多態;在4 295 bp和7 637 bp位點瀘寧雞和7 725 bp位點米易雞的PIC<0.25,表現出低度多態;其他各位點均表現出高度多態。
2.4 多態性與生長性狀間的關聯性
經多態性與生長性狀間的關聯性分析,外顯子1中2 100 bp位點CC基因型米易雞的心臟重顯著高于TT型,TT基因型的屠宰率顯著高于CC型,TT基因型的脛圍極顯著大于CC型;內含子1中4 359 bp位點TT基因型瀘寧雞的腺胃重極顯著高于CT型; 3′非編碼區7 725 bp位點AG基因型瀘寧雞的腺胃重、米易雞的腹脂重顯著高于GG型,GG基因型的半凈膛率顯著高于AG型。表明,突變位點對瀘寧雞生長性狀的影響較米易雞更為明顯。各位點其他性狀間差異不顯著。
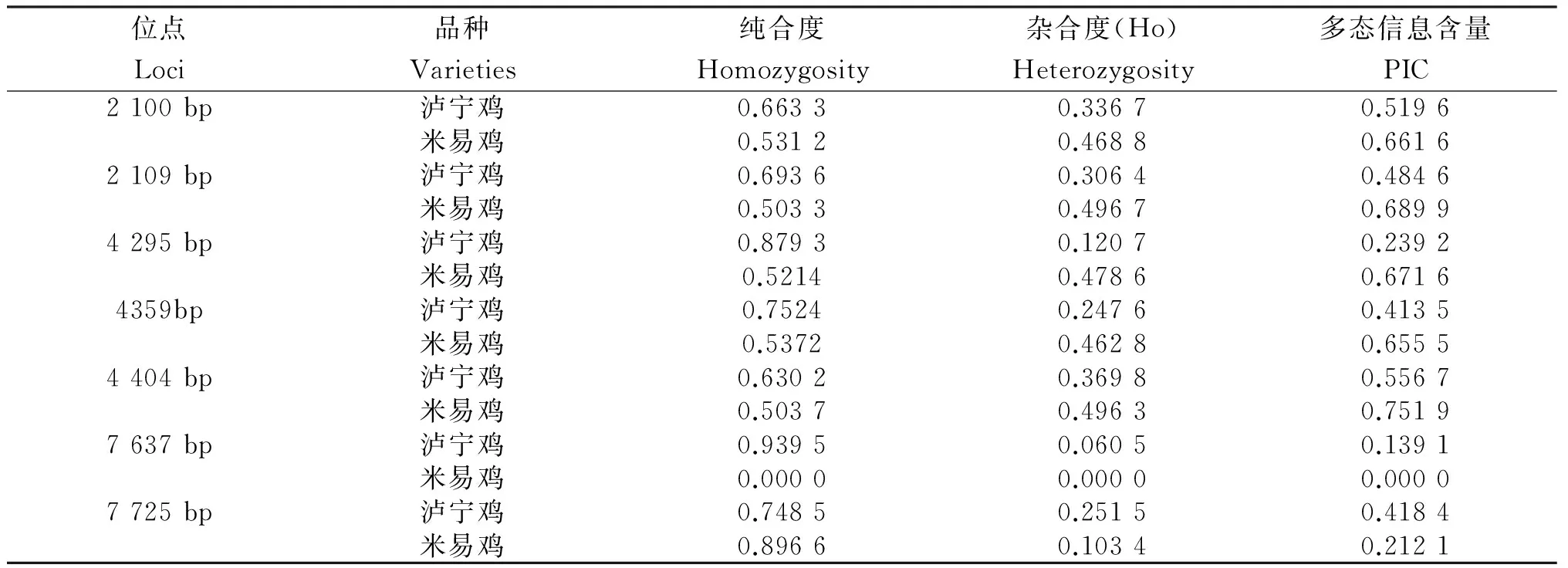
表3 MSTN各位點基因的純合度、雜合度和多態信息含量
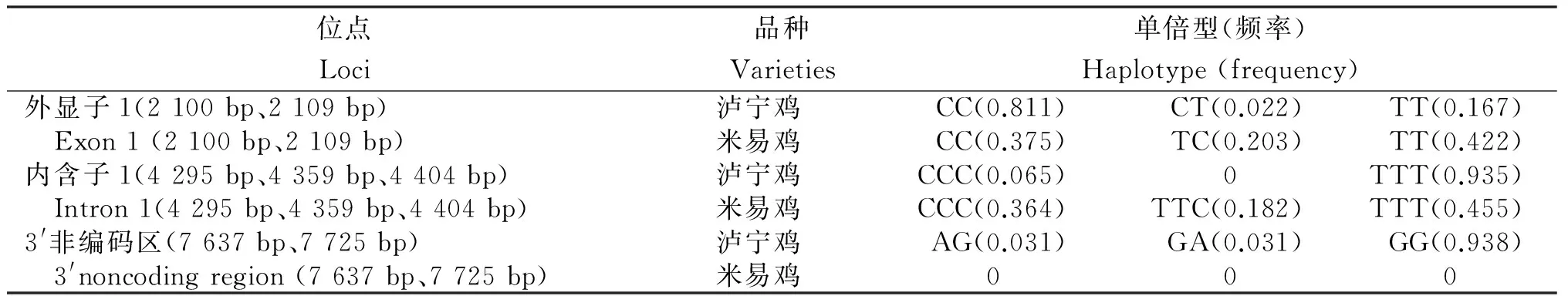
表4 瀘寧雞和米易雞MSTN基因的單倍型及頻率
2.5MSTN基因的單倍型
從表4看出:在外顯子1上瀘寧雞和米易雞分別檢測到3種單倍型,瀘寧雞CC單倍型頻率最大,CT頻率最小;米易雞3種單倍型分布差距較小。內含子1上瀘寧雞有2種單倍型,TTT單倍型明顯高于CCC單倍型;米易雞有CCC、TTC和TTT 3種單倍型,TTT單倍型的頻率最高。 3′非編碼區上只在瀘寧雞中發現AG、GA和GG 3種單倍型,GG單倍型的頻率明顯高于AG和GG型。若位點間無連鎖,n個突變位點應產生2n種單倍型,而在本研究中利用軟件均未檢測到全部單倍型,說明位點間可能處于緊密連鎖不平衡狀態[14]。
3 結論與討論
顏文錦等[15]對MSTN基因單核苷酸多態性與京海黃雞初生重和1、4、8、12周齡體重等性狀進行方差分析表明,各多態位點對體重有顯著影響(P<0.05)。朱智等[16]對溫嶺雞外顯子1研究表明,不同基因型的活重、屠宰重、半凈膛重、全凈膛重、腿肌重、胸肌重、腹脂重和胸肌率、腹脂率、屠宰率有顯著或極顯著差異(P<0.05或P<0.01)。研究結果表明,瀘寧雞和米易雞MSTN基因共發現7個SNPs位點,外顯子12 100 bp(C/T)位點對雞群心臟重、屠宰率和脛圍有顯著影響(P<0.05);內含子1中4 359 bp(T/C)位點雞群的腺胃重差異極顯著(P<0.01);3′非編碼區7637(A/G)位點雞群的腹脂重差異顯著,7 725 bp(A/G)位點雞群的腺胃重和半凈膛率差異顯著。外顯子1的2個突變位點均未引起氨基酸變異,為同義突變,可能是由于MSTN基因DNA序列和結構的高度保守性,密碼子的簡并性使DNA分子上堿基組成有較大余地的變動,堿基發生改變氨基酸并未產生變異;但對雞的生長性狀產生了顯著影響,推測可能是核苷酸的變化導致一些順式作用元件的調控功能改變,影響MSTN基因的表達,進而對生長性狀產生影響[17]。內含子1上堿基發生突變可能會影響基因轉錄、剪接、mRNA的穩定性,影響最終的翻譯;或是內含子上SNPs與基因的表達調控有關[18]。3′非編碼區雖然不直接參與轉錄翻譯過程,但是堿基的突變可能導致某個轉錄因子的結合部位發生改變,從而導致基因的表達水平發生變化。3′末端發生變異可能影響DNA二級結構,進而影響轉錄穩定性及翻譯效率等。
多數位點的基因雜合度(Ho)和多態信息含量(PIC)表現出米易雞高于瀘寧雞,說明米易雞群體的遺傳多樣性較瀘寧雞群高。瀘寧雞在各個變異位點均處于Hardy-Weinberg平衡狀態,米易雞在C2109T、C4295T和4359(C/T)位點都極顯著偏離Hardy-Weinberg平衡,這可能與人工選擇某種性狀有關。顧志良等[6]研究發現,MSTN基因SNPs位點對腹脂重、腹脂率、初生重、胸肌重和胸肌率有影響。本研究未檢測全部基因型,可能造成某些生長性狀差異不顯著的結果。
由于SNP屬于二態性遺傳標記,多態信息含量較低,對于復雜的多基因性狀,分析某一基因中單個位點的多態性往往不能準確得出基因或等位基因與性狀真實的相關性,而需要同時對多個位點組成的單倍型進行分析,才能找出與某種表型相關聯的遺傳標記[19]。研究結果表明,瀘寧雞在外顯子1上有優勢單倍型CC,頻率為0.811;在內含子1上和3′非編碼區都檢測到具有明顯優勢的單倍型,分別為TTT(0.935)和GG(0.938)。米易雞在外顯子1和內含子1各位點上單倍型頻率差異不大,因其在7 637 bp處未發生突變,所以沒有構建單倍型。單倍型各位點處于連鎖不平衡狀態。
MSTN基因突變影響心臟重、腺胃重、腹脂重、脛圍、屠宰率和半凈膛率,單倍型分析各位點處于連鎖不平衡狀態。因此,可將MSTN基因作為地方雞生長性狀的候選基因。
[1] Mcpherron A C,Lee S J.Double muscling in cattle due to mutationsin the myostatin gene[J].Proc Natl Acad Sci,1997,94:12457-12461.
[2] Lee S J.RegμLation of muscle mass by myostatin[J].Annual Review of Cell and Developmental Biology,2004,20:61-81.
[3] Lee S J,Mcpherron A C.Myostatin and the control of skeletal muscle mass[J].Curr Opin Genet Dev,1999,9(5):604-607.
[4] 胡 蘭,王 娜,胡 銳,等.大骨雞MSTN基因的表達檢測[J].中國家禽學報,2003,7(1):46-48.
[5] 顧志良,朱大海,李 寧,等.雞Myostatin基因單核苷酸多態性與骨骼肌和脂肪生長的關系[J].中國科學:C輯,2003,33(3):273-280.
[6] 顧志良,張海峰,朱大海,等.雞Myostatin基因單核苷酸多態性的群體遺傳學分析[J].遺傳學報,2002,29(7):565-570.
[7] 楊 寧.現代養雞生產[M].北京:北京農業大學出版社,1993.
[8] 肖 雄,羅慶斌.利用DNA池技術分析鵝HSP70基因的多態性[C].楊凌:第十五次全國動物遺傳育種學術討論會論文集,2009.
[9] 宋桃偉,李敬瑞,劉若余,等.利用DNA池技術研究豬GH基因啟動子序列的多態性[J].生物技術,2011,21(6):43-46.
[10] 崔建勛,杜紅麗,張細權.利用DNA池和測序技術快速篩查SNPs及估算基因頻率[J].遺傳學報,2005,32(4):372-377.
[11] 湯賢春,路 健,李學英.PCR-SSCP技術在基因多態分析中的應用[J].中國西部科技,2010,6(9):55-57.
[12] 趙 爽,潘秋麗,姜宮凌俠.PCR-SSCR的效果分析[J].生物技術通報,2010(4):132-134,155.
[13] 張 利.黎曉敏.PCR-SSCP技術研究現狀[J].飼料博覽,2011(6):16-18.
[14] 劉錚鑄,李祥龍,鞏元芳,等.綿羊MSTN基因內含子2和外顯子3部分序列的SNP檢測和單倍型分析[J].中國畜牧雜志,2010,46(7):9-11.
[15] 顏文錦,李寧川,于佳慧,等.Myostatin基因單核苷酸多態性與京海黃雞體重的相關性分析[J].上海畜牧獸醫通訊,2007(5):15-17.
[16] 朱 智,吳登俊,徐寧迎.雞Myostatin基因單核苷酸多態性及其對屠體性狀的遺傳效應分析[J].遺傳,2007,29(5):593-598.
[17] 溫彥濤,武子寅,趙振華,等.雞MSTN基因多態性及其與屠體性狀的關聯分析[J].中國家禽,2012,34(16):29-32.
[18] 王悅冰,郎志宏,黃大昉.內含子對真核基因表達規律的影響[J].生物技術通報,2008(4):1-4.
[19] 張根喜,丁馥香,趙秀華,等.肌肉生長抑制素基因外顯子3的多態性及其與邊雞繁殖性狀的關聯分析[J].中國畜牧雜志,2012,48(1):9-11.
(責任編輯: 馮 衛)
Polymorphism of Myostatin (MSTN) Gene and Correlation BetweenMSTNPolymorphism and Growth Traitss in Luning and Miyi Chicken
ZUO Bin1, REN Bingbing1, JIANG Xiaosong2, XIONG Yinglong3, WANG Kanghuan1,JIANG Li1, LIU Guangwei1, WANG Hai1, XU Ya’ou1*
(1.CollegeofLifeScienceandTechnology,SouthwestUniversityforNationalities,Chengdu,Sichuan610041, 2.SichuanAcademyofLivestockSciences,Chengdu,Sichuan610066; 3.YuanshengAgriculturalIntegratedDevelopmentLLC.,Mianning,Sichuan615600,China)
MSTN polymorphism of Luning Chicken and Miyi Chicken was detected by PCR-SSCP and main growth traits of Luning Chicken and Miyi Chicken were determined to analyze the correlation betweenMSTNpolymorphism and growth traits of Luning Chicken and Miyi Chicken. Results:There are two SNPs loci of 2 100 bp(C/T) and 2 109 bp(C/T)in exon 1 ofMSTNgene and there are 5 genotypes of CT, TT(2 100 bp), CC, CT and TT(2 109 bp) related to heart weight, shank girth and slaughter rate. Three haplotypes of CC,CT and TT are identified in Luning Chicken and Miyi Chicken. There are three SNPs loci of 4 295 bp(T/C), 4 359 bp(T/C) and 4 404 bp(A/C) in intron 1 and there are CT(4 295 bp), CT(4 359 bp), TT, CT and CC(4 404 bp)genotypes related to glandular stomach weight significantly. Luning Chicken has two haplotypes of CCC and TTT but Miyi Chicken has three haplotypes of CCC, TTC and TTT. There are two SNPs loci of 7 637 bp(A/G) and 7 725 bp(A/G) in 3’ non-coding region and there are AG(7 637 bp), AA, AG and GG(7 725 bp)genotypes related to glandular stomach weight, abdominal fat weight and half-eviscerated weight. Luning Chicken has three haplotypes of AG, GA and GG.MSTNgene is one of major genes influencing main growth traits of Luning and Miyi Chicken and should be used as a candidate gene for breeding of local chicken varieties.
Luning Chicken; Miyi Chicken; myostatin (MSTN); PCR-SSCP; growth trait; haplotype
2015-03-05; 2015-05-28修回
四川省畜禽育種攻關項目“肉雞配套系選育”(2011NZ0099-6);四川省應用基礎項目“四川民族地區主要地方雞種遺傳資源及肉質性狀的遺傳特性研究”(2013JY0044);西南民族大學研究生創新型科研項目(CX2014SZ105)
左 斌(1991-),男,在讀碩士,研究方向:分子遺傳育種。E-mail:990363506@qq.com
*通訊作者:徐亞歐(1957-),男,教授,從事動物遺傳資源研究。E-mail:xuyaou@163.com
1001-3601(2015)07-0353-0026-05
S831
A
