運用穩定同位素技術分析大寧河主要魚類營養層級
2015-03-03 08:02:12鄧華堂巴家文段辛斌朱峰躍劉紹平宋明江陳大慶
水生生物學報 2015年5期
關鍵詞:營養
鄧華堂巴家文段辛斌朱峰躍劉紹平宋明江陳大慶
(1. 中國水產科學研究院長江水產研究所農業部長江中上游漁業資源環境科學觀測實驗站, 武漢 430223;2. 西南大學生命科學學院/動物科技學院, 重慶 400715)
運用穩定同位素技術分析大寧河主要魚類營養層級
鄧華堂1,2巴家文1,2段辛斌1朱峰躍1劉紹平1宋明江1,2陳大慶1
(1. 中國水產科學研究院長江水產研究所農業部長江中上游漁業資源環境科學觀測實驗站, 武漢 430223;2. 西南大學生命科學學院/動物科技學院, 重慶 400715)
運用氮穩定同位素技術分析了大寧河靜水水域和流水河段主要魚類的氮穩定同位素比值和營養層級,并對靜水水域不同水文時期相同魚類的營養層級進行了比較研究。結果表明, 顆粒有機物(POM)氮穩定性同位素變化幅度較大, 并存在季節差異。大寧河下游靜水水域魚類 δ15N值范圍為 4.54‰—17.51‰, 營養級處于1.51—3.88, 平均營養層級為2.49; 上游流水水域魚類的δ15N值范圍為2.25‰—10.81‰, 營養層級范圍為1.49—4.01, 平均營養層級為 2.87。大寧河上游魚類的平均營養層級大于下游靜水水域, 可能是由于上游底棲生物豐富, 魚類傾向攝食適口性更高的動物性食物而導致。大寧河下游靜水水域汛期的魚類營養層級較非汛期的值顯著降低, 可能是因為汛期的水文擾動影響魚類攝食中間捕食者, 以及水位的降低導致魚類食物競爭增加迫使其攝食低營養水平的食物。重復基準生物采樣建立精確充足的基線值以及確定合適的富集度,是提高營養層級評估準確性的重要手段。
穩定同位素; 大寧河; 魚類; 營養層級
在自然界中, 穩定同位素的分餾效應(Isotope fractionation)使得生態系統中各組分的穩定同位素自然豐度會產生一定程度的差異[1]。利用穩定同位素技術對這種差異進行分析, 進而解釋其產生的理化及生態學原因, 成為研究生態系統結構與功能的主要手段之一[2—4]。用于同位素分析的元素有多種,根據各種元素的在生物體內、組織或化合物中體現出的不同的同位素特征, 分別具有其各自的應用范圍[5]。消費者的氮穩定同位素相對于其食物將富集2‰—5‰[6], 因此氮同位素常用于確定食物網中各物種的相對營養位置[7]以及食物鏈的長度[8—10]。為評估食物網的營養結構, 通常是選取一種通用基線值(General baseline)來確定食物網中每個物種的相對營養位置, 從而對食物網中每個物種的營養關系以及食物鏈長度進行研究[6,9,11], 同時也能進行不同時間序列[11]或不同食物網[12]的對比研究。這類方法在湖泊[7, 13]、河流[11, 14]以及海洋[12, 15, 16]生態系統食物網研究中已經廣泛運用。然而, 由于基準生物也存在一定的變異性[17], 河流中食物網營養結構的對比性研究報道并不多見[11]。
在三峽水庫蓄水后, 庫區的水生生態系統結構與功能發生了改變。為充分開發利用三峽庫區水域資源, 推進庫區生態漁業發展, 水生生物增殖放流已作為庫區魚類種群自然恢復和增殖的重要手段之一。作為增殖放流試點區域, 大寧河每年開展多種經濟魚類增殖放流活動。目前關于庫區干流的食物來源[18]、食物網動態變化[19]的研究已有相關報道,然而對大寧河漁業生態系統的營養結構和生態位狀況并不了解。本文通過氮穩定同位素技術方法, 分析了大寧河主要魚類的營養層級組成和食物鏈, 進而探討不同水文時期的魚類營養層級變化趨勢, 為了解大寧河魚類的營養關系及營養結構的變動情況,構建大寧河漁業生態系統食物網結構奠定基礎, 以期進一步為增殖放流實施及效果評估體系建立提供理論基礎。
1 材料與方法
1.1 調查區域與時間
大寧河為三峽庫區一級支流, 全長202 km, 流域面積4415.84 km2[20], 河口距三峽大壩約120 km。三峽大壩蓄水后, 受回水影響河口巫山-水口段形成靜水水域, 該區域長約40 km, 平均寬度約60 m, 河床平均水深達38.3 m[21]。龍溪(距河口約50 km)以上江段仍為流水生境, 該段河道彎曲, 灘多流急, 河床主要由砂礫卵石組成。受長江流域汛期及三峽庫區水位調節的影響, 大寧河下游的水文變化一般分為汛期和非汛期, 大寧河靜水區域每年的 5月下旬開始水位顯著下降, 進入汛期, 由于三峽水庫 5—8月進行汛期調度, 大寧河下游水位波動較大; 在 9月中旬水位回升, 10月達到最高水位175 m后并維持高水位, 在來年1月水位緩慢下降, 直至5月, 在此期間水位總體較穩定。
為全面了解大寧河魚類食物網營養層級, 本研究在2011年5月、10月, 2012年5月、10月, 2013年6月、9月、12月及2014年3月進行調查, 同時在靜水生境和流水生境分別采樣。靜水水域魚類采樣點設置為巫山和大昌, 兩點相距 30 km, 樣本主要是通過三層刺網、地籠和板罾獲得。流水水域魚類在巫溪及以上江段采集, 樣本主要是通過刺網和地籠捕獲。作為食物網基準生物的螺類于巫山、大昌采集, 蜉蝣類于寧廠、巫溪河段采集后混合。顆粒有機物(POM)采樣點設置為巫山、雙龍、大昌、巫溪(圖1)。
1.2 樣品采集和測定
魚類樣品采集后, 測量記錄體長(mm)和體重(g), 取其背部肌肉, 用去離子水沖洗后, 在 60℃下連續烘干至恒重, 使用研缽研碎成均勻粉末, 干燥保存。POM樣品在各采樣點采上中下層水混合, 先通過浮游動物網過濾(網目 112 μm), 然后抽濾到預先灼燒的 Whatman GF /C纖維濾膜上獲得, 用去離子水沖洗后, 于 60℃烘干 24h至恒重, 用錫箔紙包裹后保存。底棲動物螺于地籠中收集, 并將其放置于充分曝氣的蒸餾水中過夜, 使其腸含物排空; 去殼取肌肉在 60℃下連續烘干至恒重, 用研缽研成粉末, 干燥保存; 蜉蝣用小型手抄網于上游河底石縫收集, 烘干至恒重, 干燥保存。
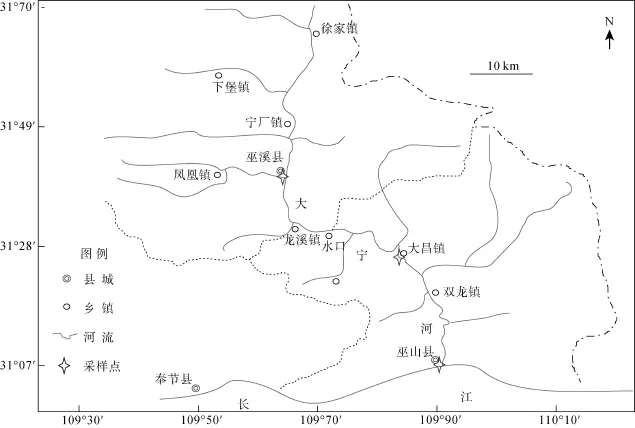
圖1 大寧河采樣點分布圖Fig. 1 The location of sampling sites in the Daning River
所有樣品的穩定性同位素比值測定均使用Flash EA1112 HT元素分析儀和DELTA V Advantage同位素比率質譜儀(中國林業科學院)進行。碳氮穩定性同位數分析分別采用國際標準物質 PDB(Pee Dee Belnite)和標準大氣氮對比。測定精度 δ15N<0.2‰。
1.3 營養層級計算營養層級(Trophic level)的計算公式如下:
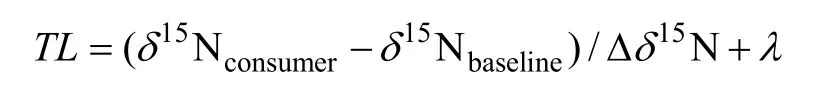
式中, TL為消費者的營養層級,15consumerN為消費者氮穩定同位素比值,15baselineN
δ為系統中作為基線生物的氮穩定同位素比值,15NδΔ 為相鄰營養級之間氮穩定同位素富集度, λ為該基線生物的營養級,初級生產者時λ=1, 初級消費者時λ=2。
Δδ
營養層級的計算首先需要確定該系統中的基線生物和富集度。基線生物一般為生態系統中常年存在、食性簡單初級消費者[6,7]。本文中靜水水域選取銅銹環棱螺(Bellamya aeruginosa)為基線生物; 上游流水水域選取在上游廣泛分布的浮游目似動蜉屬(Cinygmina spp.)為基線生物。富集度以文獻中淡水生態系統中普遍運用的數值3.4‰[6]。
1.4 統計分析
對 POM 氮穩定性同位素值, 采用單因素和雙因素方差分析其時空變化特征。根據大寧河靜水水域水位變動的規律以及魚類肌肉同位素具有的低周轉率和半衰周期[2], 將 9—10月采集的魚類作為受汛期影響的樣品, 12月至來年6月采集的魚類作為水位穩定時期的樣品(非汛期), 研究不同水文時期魚類食物網營養層級變動規律; 采用配對雙樣本 t檢驗, 對不同時期的同種魚類穩定性同位素值及營養層級進行差異分析。以 P<0.05作為差異顯著水平。對上述統計分析均通過SPSS 16.0軟件完成, 圖像處理采用CorelDRAW 12.0軟件。實驗數據用平均數±標準差(Mean±SD)表示。
2 結果
2.1 顆粒有機物(POM)氮穩定性同位素時空變化特征
大寧河靜水水域的巫山、雙龍、大昌三個采樣點的δ15N值范圍為0.73‰—11.99‰, 均值為4.83‰,上游巫溪的 δ15N值范圍為 3.59‰—7.14‰, 均值為4.97‰, 各采樣點不同季節POM的δ15N值見表1。季節與位點的雙因素方差分析表明, 不同季節(F= 2.6508, P=0.1060), 不同位點(F=2.4845, P=0.1205)的POM氮穩定性同位素值均無顯著差異。單因素方差分析表明, 不同位點POM的δ15N值無顯著差異(F=2.0570, P=0.1366); 不同季節POM的δ15N值有顯著性差異(F=3.1702, P<0.05)。多重比較顯示, 夏季和秋季之間存在顯著差異(P<0.05), 其他季節間的差異均不顯著。靜水區域POM的δ15N均值(三個采樣點)與上游流水區域(巫溪)無顯著差異(t=–0.1208, P=0.9049)。
2.2 主要魚類氮穩定性同位素值及其營養層級
大寧河下游靜水水域共測定 32種魚類氮穩定同位素樣本, 共計 183個(表 2)。靜水水域魚類的δ15N值范圍為4.54‰—17.51‰, 相差達12.96‰, 平均值為(9.64±2.34)‰。δ15N均值最高的魚類為光澤黃顙魚(Pelteobagrus nitidus), 其值為(14.01±0.41)‰;均值最低的為團頭魴(Megalobrama amblycephala),為(5.38±0.12)‰(表2)。作為基線生物的銅銹環棱螺(Bellamya aeruginosa), 非汛期采集到的δ15N平均值為(7.05±0.96)‰(n=3), 汛期采集螺的平均值為(9.31±0.01)‰(n=2)。
在靜水水域中, 魚類平均營養層級為 2.49(表2)。營養級最高的魚類為光澤黃顙魚與銅魚(Coreius heterodon), 營養層級均為 3.88; 營養級最低的魚類為團頭魴, 營養層級為1.51。全部種類中, 營養層級小于2的魚類有6種, 在2—3間的魚類共21種, 大于3的種類共5種。每種魚類的不同個體間的營養層級范圍變化較大, 共有17種魚類的營養層級范圍幅度變化超過 1個營養級, 其中蛇(Saurogobio dabryi)變化范圍最大, 幅度達到2.87個營養層級。

表1 各采樣點不同季節POM氮穩定性同位素值Tab.1 The seasonal values of the stable nitrogen isotope ratio of POM at each site
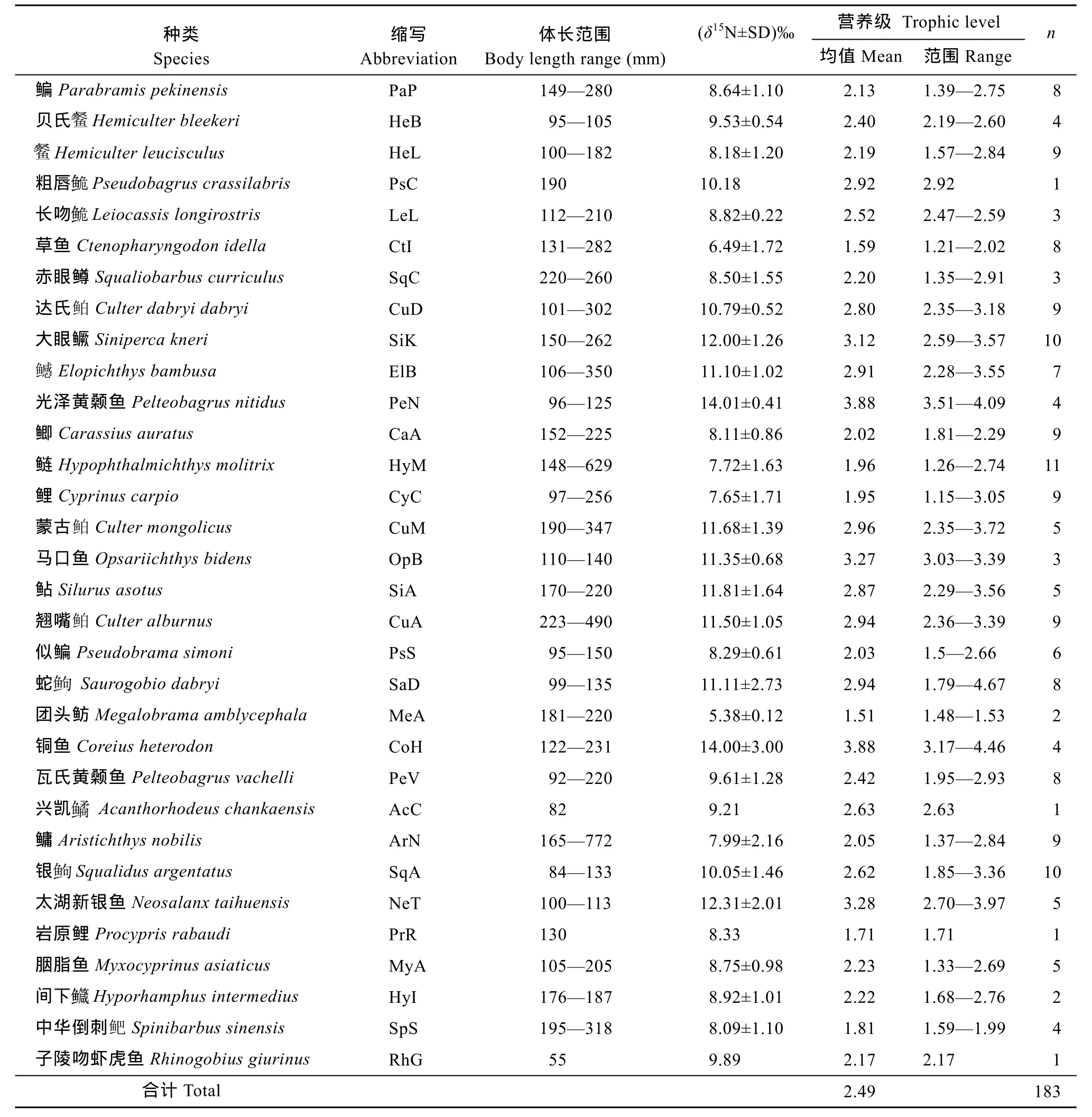
表2 大寧河下游靜水水域主要魚類的δ15N值和營養層級Tab. 2 δ15N values and trophic levels of major fish species in the lentic waters of the downstream of the Daning River
大寧河上游流水水域共測量魚類氮穩定同位素樣本46個, 共15種魚類(表3)。流水水域魚類的δ15N值范圍為2.25‰(中 華倒刺 鲃Spinibarbus sinensis)–10.81‰(大眼鱖Siniperca kneri), 相差8.56‰, 平均值為(6.94±1.44)‰。上游流水水域的魚類氮穩定同位素總體低于下游靜水水域。基線生物為似動蜉屬(Cinygmina spp.)物種, 其δ15N平均值為(3.96±3.53)‰(n=4)。上游水域魚類平均營養層級為 2.87, 顯著高于下游水域(t= –4.7842, P<0.05)。營養級小于2的魚類僅中華倒刺, 為 1.49, 大于 4的種類僅大眼鱖,為4.01, 其余種類均在2—4之間。每種魚類的營養級變化幅度較小, 均不超過1個營養級, 多數在0.5個營養級變化范圍內。
2.3 靜水水域不同時期營養層級比較
大寧河下游靜水水域, 共25種魚類在兩個時期均采集到。同種類不同時期的魚類營養層級配對檢驗, 汛期的魚類營養層級較非汛期的值顯著降低(t=5.2263, P<0.05), 平均下降0.42個營養級。圖2所示, 在汛期, 僅草魚(Ctenopharyngodon idella)、銅魚(Coreius heterodon)、鳙(Aristichthys nobilis)營養層級增加, 其余種類均有不同幅度下降。降幅達到1個營養級的魚類共有 3種, 分別為赤眼鱒(Squaliobarbus curriculus)、間下(Hyporhamphus intermedius)、胭脂魚(Myxocyprinus asiaticus)。降幅在1個營養級內的魚類共19種, 占總有降幅的魚類種數的86.36%;其中在0.5個營養級降幅內的魚類共12種。
3 討論
3.1 水庫生態系統中的食物來源及基線生物的選擇
研究表明, 水庫中浮游植物的食物源貢獻最為重要[22]。通常, 河流中的顆粒有機物(POM)除了包括內源性的浮游植物和微型碎屑之外, 還接受了陸源性動植物碎屑, 同時也包含土壤沖刷帶入的泥沙顆粒[23]。河流建壩形成靜水水域后, 回水滯留時間長, 透明度增加, POM中浮游植物的比例較大[22]。大寧河回水區域已多次暴發過水華, 水體處于中營養狀態[24], 因此, POM樣品基本代表了以藻類為主的初級生產力。然而由于大型河流POM組成復雜以及汛期徑流量的影響, 其穩定同位素值季節變異較大[23,25]。在本研究中, 大寧河流域的POM氮值變化幅度達到了11.26‰, 并出現了一定的季節變化。因此選擇POM作為基線生物, 會出現較大的誤差。而且, 三峽庫區的食物源多樣, Wang 等[18]研究表明,沿岸陸源植物的碳源貢獻對三峽庫區食物網的貢獻更為重要。因此選擇能覆蓋沿岸食物網的基線生物才能正確反映系統中各種魚類的營養層級。
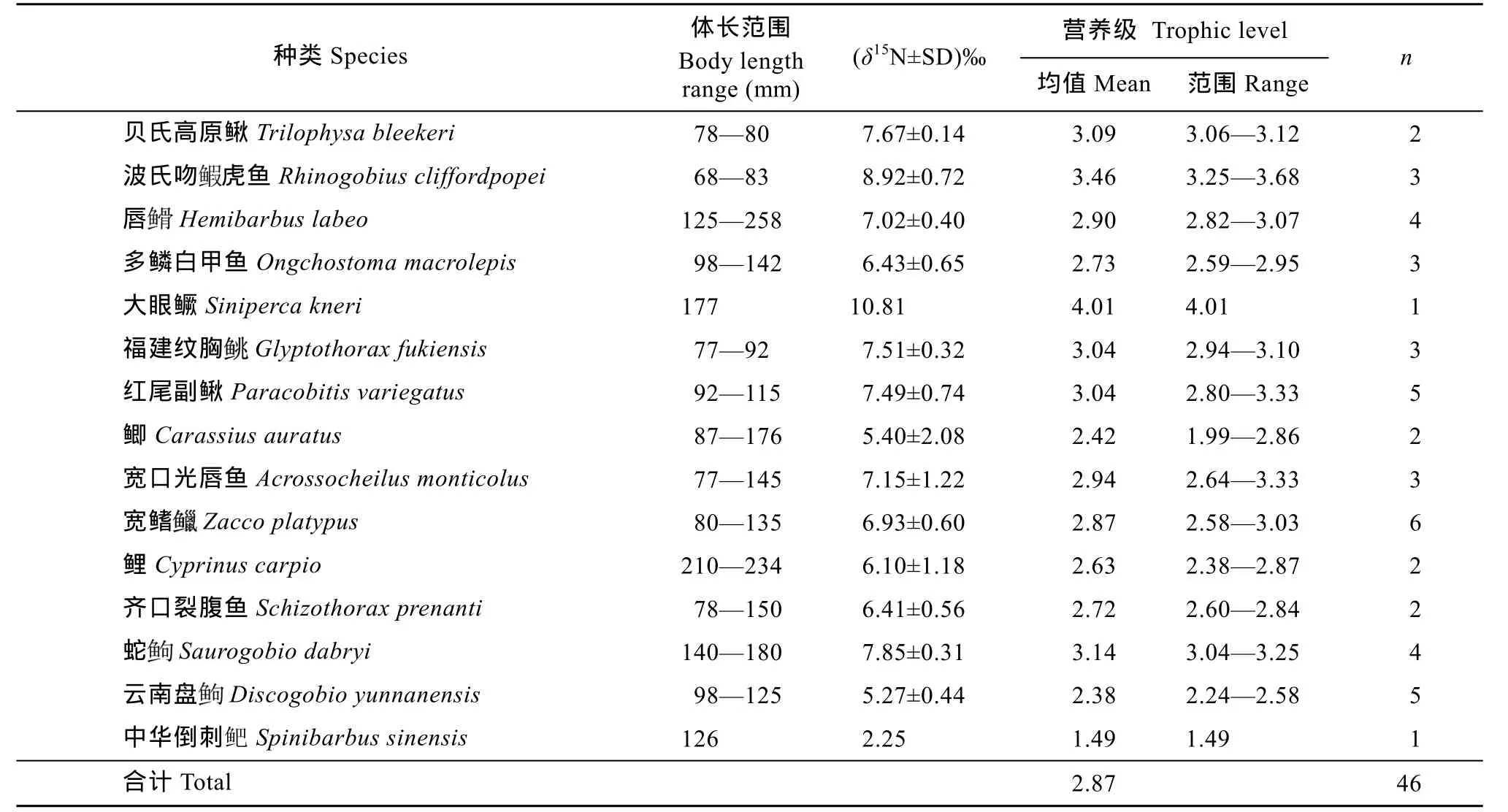
表3 大寧河上游流水水域主要魚類的δ15N值和營養層級Tab. 3 δ15N values and trophic levels of major fish species in the lotic waters of the upstream of the Daning River

圖2 不同水文時期大寧河下游魚類的營養層級比較(魚類縮寫見表2)Fig. 2 Comparison of trophic levels of fish species in the downstream of the Daning River during different hydrological periods (for abbreviation see Tab. 2)
螺類主要棲息在庫區的沿岸帶, 刮食附著藻類和攝食部分有機碎屑。螺類具有相對較低的δ15N周轉率[26], 其同位素特征值能較好的整合著生藻類和碎屑時空變化特征, 一般是沿岸帶食物網的基線生物代表[6]。雖然在部分湖泊中, 螺類氮值季節變異不大[27], 其季節變化的程度小于濾食POM的蚌類[17]。但也有報道表明, 螺類在一些水域中仍存在一定的季節差異[11,26], 這可能與環境因素相關。在時間尺度上, 螺蚌類無脊椎動物的穩定同位素特征在生產力高的富營養環境中更加易變, 而且人為外來營養的輸入也會增加這種變異趨勢[17]。因此研究不同水域或不同時期的營養層級, 其基準值的選擇也需要慎重[28]。而在大寧河流域, 在汛期和非汛期采集到的同種類螺的氮穩定同位素值具有不同的值, 因此在計算不同時期魚類的營養層級時, 選用的是各自時期的基線生物的氮值。
3.2 大寧河主要魚類營養層級變化規律
大寧河靜水水域的食物鏈長度為 3.88, 上游流水區域食物鏈長度為 4.01, 大寧河上游的魚類食物鏈更長。而且大寧河食物網縱向對比發現, 大寧河上游魚類的平均營養層級大于下游靜水庫區(P<0.05)。Mercado-Silva等[29]在墨西哥中部的Laja River研究中發現, 庫區的雜食性的鯉鯽魚類較其他流水區域有較低的營養位置, 在本研究中也有相同的現象。庫區水文條件的改變, 使得藻類和碎屑的利用性增加,這些雜食性魚類更易于攝食浮游植物[29]。而大寧河上游的喜流水性魚類多以固著藻類和底棲生物為食,魚類可能更加傾向攝食適口性更高的動物性食物[11],因此導致了上游魚類營養層級的提高。在同步的底棲動物調查中也發現, 大寧河上游底棲動物的物種數和生物量均很豐富。
在不同的水文時期, 大寧河魚類的平均營養層級有顯著的變化。在汛期, 因為上游來水(流量脈沖),庫區的水位經常會出現波動。McHugh等[30]研究表明, 河流中魚類的平均營養位置會隨著河流擾動的增加而下降。環境的擾動可能通過限制魚類食物中次級消費者的豐度而縮短食物鏈長度, 影響到魚類的平均營養層級[30], 而且魚類在水文變化大的環境中攝食到中間捕食者的機會小于穩定環境[30]。另一方面, 為防洪需求, 庫區汛期水位一直控制在較低的水平。水位的降低, 使魚類聚集的區域集中, 導致一些食物資源的限制, 增加的物種競爭迫使一些物種攝食低營養水平來獲取足夠的食物[11]。這些因素都會導致魚類平均營養層級的下降。
不同時期大寧河和魚類的平均營養層級雖有下降, 但平均降幅僅0.42個營養層級(圖2), 多數魚類并未有很大的幅度變化(未超過 0.50個營養級), 說明汛期的水文波動, 可能改變各種魚類的攝食組成,并未使魚類的食物類型出現非常明顯的轉變。并且草魚、銅魚、鳙的平均營養層級升高, 可能是由于大寧河食物網中其他的食物或能量途徑也占據重要作用的原因。而評估大型流域的營養層級, 由于食物來源的多樣性, 需要多個食物源的基準參考[6,11]。雖然,三峽庫區食物網中沿岸陸源的碳源貢獻很大[18], 但在支流庫灣潛在其他餌料資源可能比較豐富。因此,只有對三峽庫區支流食物網的食物來源進一步研究,才能更加準確全面地評估庫區支流食物網的物種營養結構組成及能量流動。
一般穩定的生態系統食物網結構復雜, 食物來源多樣, 食物層級不易波動。汛期水文的影響并未導致大寧河魚類營養層級較大的波動, 從大寧河主要魚類的營養層級可以看出, 大寧河大多數魚類的營養層級未超過3, 尤其是兇猛性 鲌類、鲇等營養級水平較低, 說明通過增殖放流建立的大寧河漁業群落結構較為穩定, 而且高層級魚類多利用低層級的食物, 潛在的替代食物可能較多。在今后的增殖放流中, 可以根據魚類的營養層級, 合理調節放流種類數量, 以提高經濟價值較高魚類比例, 同時亦可評估營養結構時間尺度的變化與漁業捕撈壓力關系, 從而為合理的捕撈規劃和漁政管理提供理論指導。
3.3 營養層級評估的準確性
根據基線生物來確定食物網中消費者營養層級的方法, 基準生物選擇和富集度 Δδ15N值是決定其準確性的兩個關鍵因素[3]。雖然選擇長壽命的無脊椎動物較周轉率低的浮游動植物作為基準生物, 對營養層級的評估更加適合[26]。但不同種類或不同季節的無脊椎動物為基準生物, 對部分魚類其營養位置的計算也會產生不同程度的偏差[17,31]。Xu等[31]利用不同基準生物計算鳙的營養位置, 營養位置為 2.4—3.9, 平均營養位置為 3.4, 與實際相差了一個營養層級。徐軍等[26]根據基準值的季節變化, 模擬評估了不同采樣頻次對營養級的影響, 發現基準值的季節變化對營養級的大小和寬幅均有較大影響。因此, 對一個食物來源復雜的生態系統, 建立精確充足的基線值是必要的[3]。
由于個體組織的生理及環境因素均會影響穩定性同位素值的富集[32], 因此理論上每個營養層次的轉化, 其富集度并不相同, 而且不同的食物鏈富集度也會有區別。Post[6]驗證了營養位置對氮富集度取值最為敏感, 而且氮富集度取值低時相較于高的富集度時更加敏感。只有在平均到多個營養路徑時, Δδ15N值為3.4‰才能是一個有效的估計值[7]。因此,當 Δδ15N值運用到個體或單個物種中時, 需要分析導致營養富集產生差異的各種原因, 并合理取值[3,32]。但對于整個食物網的物種層級評估研究, 由于3.4‰已經是許多不同營養層級之間以及消費者的食物來源之間平均值[33], 因此計算出的整個食物網層級有一定的參考性。Vander Zanden and Rasmussen[33]發現富集度在食肉動物之間的變異程度小于草食性動物, 因此利用初級消費者作為基線生物, 較初級生產者也會減少層級評估的誤差[32]。雖然營養層級的精確評估, 還需要考慮到多種食物來源, 物種個體大小以及各種組織間同位素富集的異質性等因素[1]。受實驗條件及樣本采集限制, 目前利用普遍的基準生物相對位置評估食物網的物種營養層級的方法仍然是應用最為廣泛的[2,3,28]。重復基準生物采樣增加其代表性[17], 結合不同富集度產生營養層級估算值的變化范圍, 利用各種模型如引入 Bayesian methods分析各種來源或水平上的誤差[3,34], 也是今后提高營養層級評估準確性的重要方式。
致謝:
感謝中國林業科學院穩定同位素實驗室柯淵老
[1] Middelburg J J. Stable isotopes dissect aquatic food webs from the top to the bottom [J]. Biogeosciences, 2014, 11(8): 2357—2371
[2] Boecklen W J, Yarnes C T, Cook B A, et al. On the use of stable isotopes in trophic ecology [J]. Annual Review of Ecology, Evolution, and Systematics, 2011, 42: 411—440
[3] Layman C A, Araujo M S, Boucek R, et al. Applying stable isotopes to examine food-web structure: an overview of analytical tools [J]. Biological Reviews, 2012, 87(3): 545—562
[4] Zhang H, Wu G G, Zhang H, et al. Seasonal variations of stable isotopes in fish fauna from East Lake Dongting[J]. Acta Hydrobiologica Sinica, 2013, 37(4): 796—798 [張歡,吳功果, 張歡, 等. 東洞庭湖魚類穩定同位素的季節變化.水生生物學報, 2013, 37(4): 796—798]
[5] West J B, Bowen G J, Cerling T E, et al. Stable isotopes as one of nature's ecological recorders [J]. Trends in Ecology & Evolution, 2006, 21(7): 408—414
[6] DeNiro M J, Epstein S. Influence of diet on the distribution of nitrogen isotopes in animals [J]. Geochimica et Cosmochimica Acta, 1981, 45(3): 341—351
[7] Post D M. Using stable isotopes to estimate trophic position: models, methods, and assumptions [J]. Ecology, 2002a, 83(3): 703—718
[8] Post D M. The long and short of food-chain length [J]. Trends in Ecology & Evolution, 2002b, 17(6): 269—277
[9] Sabo J L, Finlay J C, Post D M. Food chains in freshwaters [J]. Annals of the New York Academy of Sciences, 2009, 1162(1): 187—220
[10] Zhang H, He L, Zhang P Y, et al. Food chain length theory: a review [J]. Acta Ecologica Sinica, 2013, 33(24): 7630—7643 [張歡, 何亮, 張培育, 等. 食物鏈長度理論研究進展. 生態學報, 2013, 33(24): 7630—7643]
[11] Roach K A, Thorp J H, Delong M D. Influence of lateral gradients of hydrologic connectivity on trophic positions of fishes in the Upper Mississippi River [J]. Freshwater Biology, 2009, 54(3): 607—620
[12] Valls M, Sweeting C J, Olivar M P, et al. Structure and dynamics of food webs in the water column on shelf and slope grounds of the western Mediterranean [J]. Journal of Marine Systems, 2014, 138: 171–181
[13] Mao Z, Gu X, Zeng Q, et al. Food web structure of a shallow eutrophic lake (Lake Taihu, China) assessed by stable isotope analysis [J]. Hydrobiologia, 2012, 683(1): 173—183
[14] Winemiller K O, Zeug S C, Robertson C R, et al. Food-web structure of coastal streams in Costa Rica revealed by dietary and stable isotope analyses [J]. Journal of Tropical Ecology, 2011, 27(5): 463—476師對同位素測定提供的幫助。
[15] Stowasser G, Atkinson A, McGill R A R, et al. Food web dynamics in the Scotia Sea in summer: A stable isotope study [J]. Deep Sea Research Part II: Topical Studies in Oceanography, 2012, 59: 208—221
[16] Yang G H, Hou X Q, Sun X L, et al. Constructi on food web model of Liusha Bay-using stable isotope analysis results [J]. Acta Hydrobiologica Sinica, 2013, 37(1): 150—156 [楊國歡,侯秀瓊, 孫省利, 等. 流沙灣食物網結構的初探——基于穩定同位素方法的分析結果. 水生生物學報, 2013, 37(1): 150—156]
[17] Jardine T D, Hadwen W L, Hamilton S K, et al. Understanding and overcoming baseline isotopic variability in running waters [J]. River Research and Applications, 2014, 30(2): 155—165
[18] Wang J, Gu B, Huang J, et al. Terrestrial contributions to the aquatic food web in the middle Yangtze River [J]. PloS One, 2014, 9(7): e102473
[19] Li B, Wang Z J, Yang J P, et al. The dynamic and seasonal variation of the fish food webs in the mainstream of Three Gorges Reservoir [J]. Journal of Fisheries of China, 2013, 37(7):1015—1022 [李斌, 王志堅, 楊潔萍, 等. 三峽庫區干流魚類食物網動態及季節性變化. 水產學報, 2013, 37(7):1015—1022]
[20] Liu R M, Shen Z Y. Integrated assessment and changes of ecological environment in Daning River watershed [J]. Journal of Beijing Normal University (Natural Science), 2006, 42(2): 200—203 [劉瑞民, 沈珍瑤. 大寧河流域生態環境綜合評價及其演變. 北京師范大學學報 (自然科學版), 2006, 42(2): 200—203]
[21] Wang K, Duan X B, Liu S P, et al. Survey on temporal and spatial distribution of fish in the Daing River [J]. Acta Hydrobiologica Sinica, 2009, 33(3): 516—521 [王珂, 段辛斌, 劉紹平, 等. 三峽庫區大寧河魚類的時空分布特征.水生生物學報, 2009, 33(3): 516—521]
[22] Hoeinghaus D J, Winemiller K O, Agostinho A A. Landscape-scale hydrologic characteristics differentiate patterns of carbon flow in large-river food webs [J]. Ecosystems, 2007, 10(6): 1019—1033
[23] Zeug S C, Winemiller K O. Evidence supporting the importance of terrestrial carbon in a large-river food web [J]. Ecology, 2008, 89(6): 1733—1743
[24] Wang L P, Zheng B H, Zhang J L, et al. Effects on euthrophication and hydrodynamics of Daning River after impoundment of Three Gorges Reservoir [J]. Journal of Lake Sciences, 2012, 24(2): 232—237 [王麗平, 鄭丙輝, 張佳磊, 等. 三峽水庫蓄水后對支流大寧河富營養化特征及水動力的影響. 湖泊科學, 2012, 24(2): 232—237]
[25] Pingram M A, Collier K J, Hamilton D P, et al. Spatial and temporal patterns of carbon flow in a temperate, large river food web [J]. Hydrobiologia, 2014, 729(1): 107—131
[26] Xu J, Zhang M, Xie P. Variability of stable nitrogen isotopic baselines and its consequence for trophic modeling [J]. Journal of Lake Sciences, 2010, 22(1): 8—20 [徐軍, 張敏,謝平. 氮穩定同位素基準的可變性及對營養級評價的影響. 湖泊科學, 2010, 22(1): 8—20]
[27] Mao Z G, Gu X H, Zeng Q F, et al. Seasonal and spatial variations of the food web structure in a shallow eutrophic lake assessed by stable isotope analysis [J]. Fisheries Science, 2014, 80(5): 1045—1056
[28] Anderson C, Cabana G. Estimating the trophic position of aquatic consumers in river food webs using stable nitrogen isotopes [J]. Journal of the North American Benthological Society, 2007, (2): 273—285
[29] Mercado-Silva N, Helmus M R, Zanden M. The effects of impoundment and non-native species on a river food web in Mexico's central plateau [J]. River Research and Applications, 2009, 25(9): 1090—1108
[30] McHugh P A, McIntosh A R, Jellyman P G. Dual influences of ecosystem size and disturbance on food chain length in streams [J]. Ecology Letters, 2010, 13(7): 881—890
[31] Xu J, Zhang M, Xie P. Sympatric variability of isotopic baselines influences modeling of fish trophic patterns [J]. Limnology, 2011, 12(2): 107—115
[32] Martínez del Rio C, Wolf N, Carleton S A, et al. Isotopic ecology ten years after a call for more laboratory experiments [J]. Biological Reviews, 2009, 84(1): 91—111
[33] Vander Zanden M, Rasmussen J B. Variation in δ15N and δ13C trophic fractionation: implications for aquatic food web studies [J]. Limnology and Oceanography, 2001, 46(8): 2061—2066
[34] Jackson A L, Inger R, Parnell A C, et al. Comparing isotopic niche widths among and within communities: SIBER–Stable Isotope Bayesian Ellipses in R [J]. Journal of Animal Ecology, 2011, 80(3): 595—602
THE ANALYSIS OF THE TROPHIC LEVELS OF THE MAJOR FISH SPECIES IN THE DANING RIVER USING STABLE ISOTOPE TECHNOLOGY
DENG Hua-Tang1,2, BA Jia-Wen1,2, DUAN Xin-Bin1, ZHU Feng-Yue1, LIU Shao-Ping1, SONG Ming-Jiang1,2and CHEN Da-Qing1
(1. Scientific Observing and Experimental Station of Fishery Resources and Environment in the Upper and Middle Reaches of the Yangtze River, Ministry of Agriculture; Yangtze River Fisheries Research Institute, Chinese Academy of Fishery Sciences, Wuhan 430223, China; 2. School of Life Science/College of Animal Science and Technology, Southwest University, Chongqing 400715, China)
In this study, we measured the stable nitrogen isotope ratios of the major fish species in the lotic reaches and the impounded reaches of the Daning River to identify their trophic levels, and we compared the mean trophic levels of several conspecifics in lentic waters during different hydrological periods. The results showed that the δ15N values of particulate organic matter (POM) fluctuated in a wide range and exhibited significant seasonal variation. The δ15N values of species in the lentic downstream ranged from 4.54‰ to 17.51‰, and the trophic levels of these organisms were 1.51—3.88 with an average of 2.49. In the upstream, the δ15N values of fish species varied between 2.25‰ and 10.81‰, and their trophic levels were in the range of 1.49—4.01 with an average of 2.87. The fact that the mean trophic levels were higher in the upstream than that in the downstream, which indicated that the fish tended to consume preferable invertebrate in the lotic areas probably due to their high diversity and abundant biomass. The mean trophic levels of conspecifics in the impounded waters were significantly lower in the flooding period than that in the non-flooding period. This is most likely because the hydrological disturbance affected the intermediate predators. Another possibility is that during the low-water period, the fish in the downstream need to forage for food at lower trophic levels because of the intense competition. To improve the assessment of the trophic levels, we will need to determine the appropriate trophic discrimination values and to establish an adequate isotopic baseline by repeated sampling of the baseline organisms.
Stable isotope; Daning River; Fish; Trophic level
Q958.12
A
1000-3207(2015)05-0893-09
10.7541/2015.118
2015-01-26;
2015-05-22
公益性行業(農業)科研專項經費(201303056-5); 中國長江三峽集團公司科研項目(No. 0799526; No. CT-12-08-01)資助
鄧華堂(1985—), 男, 湖北松滋人; 博士研究生; 主要從事漁業生態方面研究。E-mail: dht12202@163.com
陳大慶(1964—), 男, 研究員; E-mail: chdq@yfi.ac.cn
猜你喜歡
中老年保健(2022年5期)2022-11-25 14:16:14
今日農業(2021年11期)2021-08-13 08:53:34
今日農業(2021年4期)2021-06-09 06:59:58
今日農業(2020年24期)2020-03-17 08:58:18
中國生殖健康(2019年8期)2019-01-07 01:18:24
幸福(2018年33期)2018-12-05 05:22:46
廣東第二課堂·小學(2017年2期)2017-02-20 15:10:08
廣東第二課堂·小學(2017年1期)2017-02-20 14:47:45
少兒科學周刊·兒童版(2015年12期)2016-05-16 03:38:08
小學閱讀指南·低年級版(2016年8期)2016-05-14 13:24:45
