中國亞洲玉米螟發育歷期的地理變異
2015-03-10 08:44:33涂小云夏勤雯何海敏薛芳森
生態學報 2015年2期
關鍵詞:差異
涂小云,夏勤雯,陳 超,何海敏,薛芳森,*
1. 江西師范大學生命科學學院, 南昌 330022 2. 江西農業大學昆蟲研究所, 南昌 330045
中國亞洲玉米螟發育歷期的地理變異
涂小云1,2,夏勤雯2,陳 超2,何海敏2,薛芳森2,*
1. 江西師范大學生命科學學院, 南昌 330022 2. 江西農業大學昆蟲研究所, 南昌 330045
廣布種昆蟲可調節其生活史以適應棲息地的條件變化,如可以通過調節自身的生長發育和繁殖期,使其發生與棲息地的物候(如食料、氣溫和降雨等)同步,這對昆蟲在棲息地的繁衍具有十分重要的意義。亞洲玉米螟Ostriniafurnacalis為玉米主要害蟲,在中國從南到北都有分布,棲息地氣候也不同,不同地理種群生活史發生了分化,如化性、臨界光周期、滯育后羽化時間、體重和體型、繁殖力和抗寒力等均不同。然而該蟲發育歷期的地理變異尚未見系統報道。為此,詳細比較了來自5個亞洲玉米螟不同地理種群即廣西陽朔(YS)、江西南昌(NC)、山東泰安(TA)、河北廊坊(LF)和黑龍江哈爾濱(HEB)在20—31℃下卵、幼蟲、蛹發育歷期及其與棲息地緯度的關系。結果表明:不同溫度下的不同地理種群的卵期不存在顯著差異,在20、22和25℃下,卵期從南到北稍有延長,卵期與其棲息地緯度呈正相關;但在28℃下,不同地理種群的卵期基本相等,與緯度相關性不顯著。在31℃下,不同地理種群的幼蟲期不存在顯著差異,而在其他溫度下,不同地理種群的幼蟲期存在顯著差異;在22、25和28℃下,幼蟲期與其棲息地緯度呈正相關,而在20和31℃下,幼蟲期與其棲息地緯度呈負相關,在20、22和25℃下,最北的哈爾濱種群的幼蟲期變幅不大,而其他4個種群的幼蟲期隨著溫度的升高明顯縮短。各地理種群20℃下的雌蛹、28℃下的雄蛹和31℃下的雌雄蛹期差異均不顯著,其他雌或雄蛹期差異顯著;各溫度下的雌雄蛹期與其棲息地緯度存在正相關關系。這些研究結果揭示了廣布種昆蟲可以調節其自身發育歷期以適應棲息地環境條件。
亞洲玉米螟;地理變異;發育歷期;卵;幼蟲;蛹
生物體在應對環境變化時形成了其內在的發育策略,廣布種昆蟲可調節其生活史以適應棲息地的條件變化,如可以通過調節自身的生長發育和繁殖期,使其發生與棲息地的物候(如食料、氣溫和降雨等)同步[1- 2],這對昆蟲在棲息地的繁衍具有十分重要的意義。全發育時間(從產卵至成蟲羽化)反映了卵的發育和孵化、幼蟲的取食和生長及幼蟲或蛹變態至成蟲的速率,是環境條件如溫度、可利用的食物、光周期與生理等相互作用的體現。資料表明,在相同溫度下,同種昆蟲高緯度和低緯度種群發育歷期長短因種類不同而異。有的昆蟲北方種群發育快,而南方種群發育慢,如甘藍夜蛾Mamestrabrassica在光周期為L15:D9條件下,南方種群的幼蟲期明顯長于北方種群[3];中歐山松大小蠹Dendroctonusponderosae(Hopkins)[4]、黃臉油葫蘆Teleogryllusemma[5]和蜜蜂Exoneurarobusta的幼蜂[6]等均是北方種群發育更快;蟋蟀Dianemobiusnigrofascia若蟲的發育歷期由南到北逐漸縮短[5]。而有的昆蟲北方種群發育卻慢,發育歷期長于南方種群。如一化性蟋蟀modicogryllussiamensis若蟲期明顯長于二化性的[7- 8],舞毒蛾Lymantriadispar北方種群的蛹期亦長于南方種群[9]。草地血黑蝗Melanoplussanguinipes/devastator胚胎發育時間與采集地年平均溫度呈正相關[10],而年平均溫度顯然與緯度有關。尚有昆蟲的發育歷期與棲息地經度有關,如蚱蜢Romaleamicroptera從孵化至4齡的發育歷期由西到東逐漸延長[11]。更多昆蟲發育歷期雖存在地理變異,但與棲息地緯度無相關性或相關性不明確,如不同種類的果蠅Drosophila[12]、大眼長蝽Geocorispunctipes[13]、普通草蛉Chrysoperlacarnea[14]、莎草粘蟲Spodopteraexempta[15]、蟋蟀Dianemobiusnigrofasciatus[5]和小紅蟻Myrmicarubr[16]等。還有些昆蟲發育歷期的地理種群間差異會隨實驗條件下飼養的時間延長而減小,如李錦華將棉鈴蟲Heliothisarmigera(Hübner)安陽種群和四川種群放在相同條件下飼養,兩個種群的幼蟲期和蛹期第一代差異顯著,第二、三代差異不顯著[17]。這些研究結果揭示了昆蟲的發育歷期地理變異的不同模式,也反映了昆蟲生活史對策的多樣性。
亞洲玉米螟Ostriniafurnacalis(Guenée)是中國玉米的主要害蟲,分布廣,南自海南三亞,北至黑龍江的黑河均有分布,分布區跨越的緯度很大,地理環境差異也很大。對亞洲玉米螟不同地理種群生活史特性的研究表明,該蟲的臨界光周期[18- 23]、越冬幼蟲復蘇后的發育歷期[24- 28]、化性(年發生的世代數)和幼蟲的抗寒力[29]、體重和體型[30]、繁殖力[31]、暗期干擾的滯育反應[32]等特性均發生了相應的地理變異。不同化性亞洲玉米螟的有效積溫、發育起點溫度和各蟲態發育歷期也有些報道[22, 25, 33- 35],但主要局限于北方種群。本實驗較系統比較了5個不同地理種群亞洲玉米螟在不同溫度下的各蟲態的發育歷期,旨在探明亞洲玉米螟發育歷期地理變異的特點,以期為不同地區準確預測預報亞洲玉米螟的發生提供參考,也為深入了解昆蟲生活史的進化提供基礎資料。
1 材料與方法
1.1 供試蟲源
亞洲玉米螟分別于2010年6—7月采自廣西陽朔、江西南昌、山東泰安、河北廊坊和黑龍江哈爾濱玉米地,前四個采集地采集的蟲源主要為大齡幼蟲或蛹,哈爾濱采集的蟲源為越冬代成蟲。采集地經緯度見表1。采集蟲源在人工氣候箱(LRH- 250-GS, 廣東醫療器械廠)中(28±1)℃、RH (75±2)%、光周期L16∶D8條件下用人工飼料[36]飼養,供實驗用。
種群代碼為采集地漢語拼音的縮寫
1.2 卵發育歷期的測定
將5個地理種群的室內第1代成蟲置于保鮮袋中用10%蔗糖水飼喂、產卵,收集當天卵塊置于培養皿中,用濕濾紙保濕。觀察并記錄孵化時間。溫度設置20、22、25和28℃,光周期為L16∶D8。每種群測定的卵塊數不少于10個。
1.3 幼蟲和蛹發育歷期的測定
將5個地理種群當日孵化的幼蟲(室內繁殖的第1代)接入透明的圓形塑料盒(直徑12 cm,高15 cm) 中用人工飼料飼養,15d后(約4齡),轉入24孔板中單頭飼養,觀察其化蛹、羽化,逐個記錄化蛹和羽化時間。溫度設置20、22、25、28和31℃,光周期設置L16∶D8。每處理接蟲兩盒,每盒不少于50頭。
1.4 數據統計與分析
實驗數據采用SPSS13.0數據處理系統進行分析。用one-way ANOVA進行方差分析,結果用平均值±標準差(mean ±SD)表示,線性回歸用Linear-regression。
2 結果
2.1 不同地理種群亞洲玉米螟卵發育歷期的地理變異
不同地理種群卵的發育歷期見表2。由表2可知,同一溫度下,不同地理種群卵的發育歷期差異不顯著(在20℃下:F=2.224, df=4,49,P﹥0. 05;在22℃下:F=12.178,df=4, 55,P﹥0. 05;在25℃下:F=1.435,df=4, 64,P﹥0. 05;在28℃下:F=0.781,df=4, 64,P﹥0. 05 )。
在20、22和25℃下,卵期從南到北稍有延長,與棲息地緯度間存在著顯著的正相關關系(P<0. 01);但在28℃下,不同地理種群的卵期基本相等,與棲息地緯度間存在弱正相關關系(P﹥0. 05 ) (圖1)。
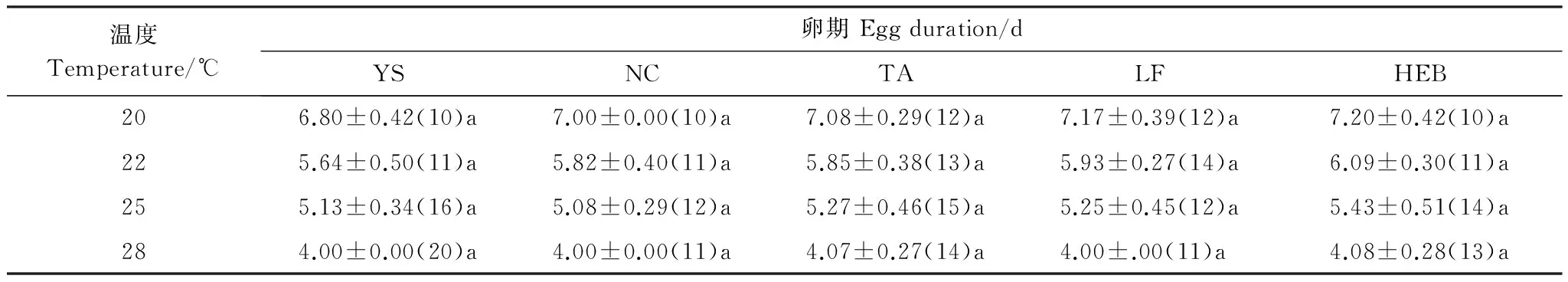
表2 不同地理種群亞洲玉米螟卵期Table 2 Egg duration in different geographic populations of Ostrinia furnacalis
括號中為測量樣本數; 表中數據為平均值±標準差, 同一行數據后不同字母表示差異顯著(Duncan多重比較,P< 0.05)

圖1 亞洲玉米螟卵發育歷期的地理變異Fig.1 Geographic variation in duration of egg of Ostrinia furnacalis
2.2 不同地理種群亞洲玉米螟幼蟲期發育歷期的地理變異
不同地理種群不同溫度下幼蟲期見表3。在20、22、25和28℃下,雌雄幼蟲期均差異顯著 (在20℃下雌幼蟲:F=7.834, df=4, 220,P<0.05;雄幼蟲:F=3.490, df=4, 137,P<0.01;在22℃下雌幼蟲:F=52.275, df=4, 227,P<0.05;雄幼蟲:F=34.415, df=4, 178,P<0.05; 在25℃下雌幼蟲:F=89.539, df=4, 352,P<0.05;雄幼蟲:F=74.876, df=4, 281,P<0.05;在28℃下雌幼蟲:F=10.070, df=4, 382,P<0.05;雄幼蟲:F=7.372, df=4, 220,P<0.05)。在31℃下,不同地理種群雌雄幼蟲期差異均不顯著 (雌幼蟲:F=0.283, df=4, 318,P> 0.05;雄幼蟲:F=0.468, df=4, 259,P> 0.05)。有趣的是,在20℃下,幼蟲期與其棲息地緯度存在弱負相關關系,31℃下雄蟲幼蟲期也顯示相似的結果(圖2)。在22、25和28℃下,幼蟲期與其棲息地緯度呈正相關(圖2)。還值得提及的是,在20、22和25℃下,最北的哈爾濱種群的幼蟲期變幅不大,如雌性幼蟲期分別是43.5、40.9和39.5 d;而其他4個種群的幼蟲期隨著溫度的升高明顯縮短,如陽朔雌性幼蟲期分別是47.9、28.4和22.5 d,南昌雌性幼蟲期分別是42.7、35.1和24.3 d,泰安雌性幼蟲期分別是42.0、38.3和25.0 d。

表3 不同地理種群亞洲玉米螟幼蟲期Table 3 Duration of larvae in different geographic populations of Ostrinia furnacalis
2.3 不同地理種群亞洲玉米螟蛹發育歷期的地理變異
不同地理種群不同溫度下蛹期見表4。由表4可知,在20℃下,不同地理種群雌蛹期差異不顯著,雄蛹期差異顯著且隨緯度上升逐漸延長 (雌蛹:F=0.405, df=4, 214,P> 0.05;雄蛹:F=4.630, df=4, 135,P<0.05)。在22和25℃下,雌雄蛹期差異顯著 (在22℃下:雌蛹F=64.423, df=4, 221,P<0.05;雄蛹F=37.131, df=4, 178,P<0.05;在25℃下:雌蛹F=2.556, df=4, 341,P<0.05;雄蛹:F=2.977, df=4, 276,P<0.05)。在28和31℃下,除28℃雌蟲蛹期差異顯著外,其他雌雄蛹期差異均不顯著 (在28℃下雌蛹:F=3.567, df=4, 321,P<0.05;雄蛹:F=2.283, df=4, 194,P>0.05;在31℃下雌蛹:F=2.152, df=4, 275,P>0.05;雄蛹:F=0.983, df=4, 242,P>0.05)。圖3表明,蛹期與其棲息地緯度存在正相關關系。
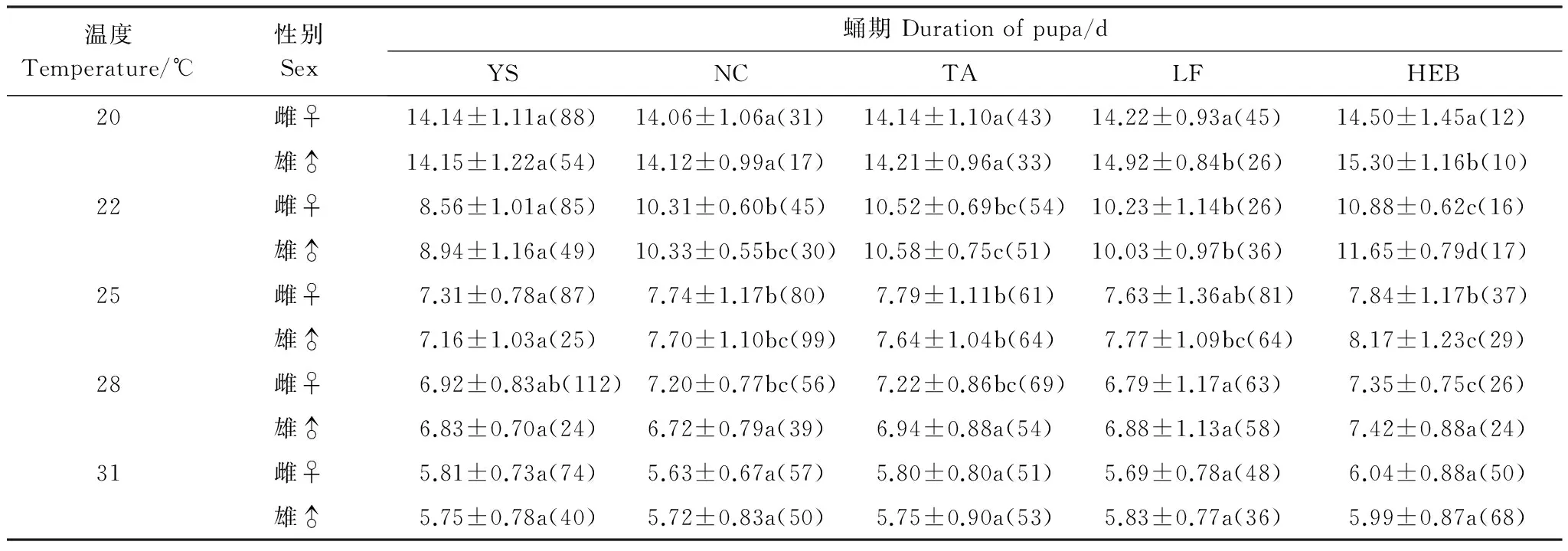
表4 不同地理種群亞洲玉米螟蛹期Table 4 Duration of pupa in different geographic populations of Ostrinia furnacalis
3 討論
前蘇聯的昆蟲學家Danilevskii首次報道了來自3個不同地理種群(蘇肯米43°N,貝爾格拉德50°N和列寧格勒60°N)梨劍紋夜蛾Acronyctarumicis蛹的發育歷期是隨緯度的上升而稍有延長[37]。捷克的Honěk分析了來自熱帶、亞熱帶和溫帶地區335種昆蟲發育起點溫度和有效積溫[38]。結果表明,卵、幼蟲和蛹的發育起點溫度是熱帶昆蟲種類最高,亞熱帶昆蟲次之,溫帶昆蟲最低;而各蟲態需求的有效積溫則相反,溫帶最高,其次是亞熱帶和熱帶昆蟲。本實驗首次揭示了來自廣泛地理緯度亞洲玉米螟在不同溫度下卵、幼蟲和蛹發育歷期的變異。在實驗溫度下,卵期隨緯度的增大而稍有延長,與其棲息地緯度呈正相關,但不同地理種群卵期差異不顯著,特別在28℃條件下,不同地理種群卵期十分接近(表2)。這一結果表明卵的發育歷期基本沒有受到地理變異的影響。在22、25和28℃下,不同地理種群幼蟲期與采集地緯度間呈正相關關系,處理間幼蟲期差異顯著;而20℃下雌雄幼蟲期及31℃下雄蟲幼蟲期與其棲息地緯度呈負相關關系,其中20℃下幼蟲期差異顯著,但31℃下幼蟲期差異不顯著(表3,圖2)。這表明幼蟲發育歷期出現了明顯的地理變異。特別值得提及的是,在20、22和25℃下,最北的哈爾濱種群的幼蟲期變幅不大,而其他四個種群的幼蟲期隨著溫度的升高明顯縮短,這可能反映了亞洲玉米螟生活史對策的進化。在哈爾濱,其有效積溫和作物生長季節只允許亞洲玉米螟繁殖1代。不同地理種群蛹發育歷期顯示了與幼蟲發育歷期相似的地理變異,除31℃外,不同地理種群蛹期差異顯著,蛹期與其棲息地緯度呈正相關(表4,圖3)。

圖2 亞洲玉米螟幼蟲發育歷期的地理變異Fig.2 Geographic variation in duration of larvae of Ostrinia furnacalis

圖3 亞洲玉米螟蛹發育歷期的地理變異Fig.3 Geographic variation in duration of pupa of Ostrinia furnacalis
COⅠ基因和COⅡ基因分析認為亞洲玉米螟遺傳分化與地理位置具有一定的相關性[39- 40]或不具有顯著的相關性[41]。而亞洲玉米螟不同地理種群的遺傳分化常以一些表現型的地理變異反映出來。本研究結果證實,不同地理種群亞洲玉米螟幼蟲和蛹的發育歷期等表現型存在顯著的地理差異,但這些差異會隨實驗溫度變化而變化,特別在高溫下,發育歷期的差異明顯縮小。這些結果進一步揭示了即使在同一種類昆蟲中,不同實驗條件(如溫度)下,其各個蟲態發育歷期的地理變異也可能不同。
[1] Tauber C A, Tauber M J. Insect seasonal cycles: genetics and evolution. Annual Review of Ecology and Systematics, 1981, 12: 281- 308.
[2] Tauber M J, Tauber C A, Masaki S. Seasonal Adaptation of Insects. New York: Oxford University Press, 1986.
[3] Masaki S. Geographic adaptation in the seasonal life cycle ofMamestrabrassice(Linné) (Lepidoptera: Noctuidae). Bulletin of Faculty of Agricultrual Hirosaki University, 1968, 14: 16- 26.
[4] Bentz B J, Logan J A, Vandygriff J C. Latitudinal variation inDendroctonusponderosae(Coleoptera: Scolytidae) development time and adult size. The Canadian Entomologist, 2001, 133(3): 375- 387.
[5] Masaki S. Geographical variation of life cycle in crickets (Ensifera: Grylloidea). European Journal of Entomology, 1996, 93: 281- 302.
[6] Cronin A L, Schwarz M P. Latitudinal variation in the life cycle of allodapine bees (Hymenoptera: Apidae). Canadian Journal of Zoology, 1999, 77(6): 857- 864.
[7] Taniguchi N, Tomioka K. Duration of development and number of nymphal instars are differentially regulated by photoperiod in the cricketModicogryllussiamensis(Orthoptera: Gryllidae). European Journal of Entomology, 2003, 100(2): 275- 281.
[8] Tanaka S, Arai T, Tanaka K. Nymphal development, diapause and cold-hardiness in a nymph-overwintering cricket. Entomological Science, 1999, 2: 173- 183.
[10] Dingle H, Mousseau T A. Geographic variation in embryonic development time and stage of diapause in grasshopper. Oecologia, 1994, 97(2): 179- 185.
[11] Jannot J E, Brinton J, Kocot K, Akman O, Juliano S A. Ontogenetic mechanisms underlying a geographic size cline in a grasshopper,Romaleamicroptera. Annals of the Entomological Society of America, 2009, 102(3): 467- 475.
[12] van der Linde K, Sevenster J G. Local adaptation of developmental time and starvation resistance in eightDrosophilaspecies of the Philippines. Biological Journal of the Linnean Society, 2006, 87(1): 115- 125.
[13] Ruberson J R, Yeargan K V, Newton B L. Variation in diapause responses between geographic populations of the predatorGeocorispunctipes(Heteroptera: Geocoridae). Annals of the Entomological Society of America, 2001, 94(1): 116- 122.
[14] Chang Y F, Tauber M J, Tauber C A, Nyrop J P. Interpopulation variation inChrysoperlacarneareproduction: implications for mass-rearing and storage. Entomologia Experimentalis et Applicata, 2000, 95(3): 293- 302.
[15] Wilson K, Gatehouse A G. Seasonal and geographical variation in the migratory potential of outbreak populations of the African armyworm moth,Spodopteraexempta. Journal of Animal Ecology, 1993, 62(1): 169- 181.
[16] Kipyatkov V, Lopatina E, Imamgaliva A. Duration and thermal reaction norms of development are significantly different in winter and summer brood pupae of the antsMyrmicarubraLinnaeus, 1758 andM.ruginodisNylander, 1846(Hymenoptera: Formicidae). Myrmecologische Nachrichten, 2005, 7: 69- 76.
[17] 李錦華. 不同地理種群棉鈴蟲飼養比較. 昆蟲知識, 1982, 21(4): 7- 8.
[18] 杜正文, 蔡蔚琦. 玉米螟在江蘇光周期的反應初報. 昆蟲學報, 1964, 13(1): 129- 132.
[19] 王承綸, 張榮, 桂承明, 王蘊生. 亞洲玉米螟生物學特性研究——Ⅰ. 光周期反應對玉米螟發育的關系. 吉林農業科學, 1980, (3): 53- 58.
[20] 熊繼文, 蒙黔英. 亞洲玉米螟貴陽種群的滯育及其解除的初步研究. 貴州農學院學報, 1984, (2): 50- 59.
[21] 沈榮武, 薛芳森, 朱杏芬. 玉米螟化性及田間滯育發生時間的研究. 江西植保, 1988, (1): 18- 19.
[22] 魯新, 李建平, 王蘊生. 亞洲玉米螟化性類型的初步研究. 玉米科學, 1995, 3(1): 75- 78.
[23] Xia Q W, Chen C, Tu X Y, Yang H Z, Xue F S. Inheritance of photoperiodic induction of larval diapause in the Asian corn borerOstriniafurnacalis. Physiological Entomology, 2012, 37(2): 185- 191.
[24] 魯新, 周大榮, 李建平. 亞洲玉米螟越冬幼蟲化性與復蘇后發育歷期的關系. 玉米科學, 1998, (增刊): 100- 102.
[25] 魯新, 周大榮. 吉林省亞洲玉米螟化性類型與其發育歷期的關系. 植物保護學報, 1999, 26(1): 1- 6.
[26] 馬瑞, 錢海濤, 董輝, 夏新, 叢斌. 不同地理種群亞洲玉米螟越冬幼蟲復蘇后的發育歷期研究. 湖北農業科學, 2008, 47(5): 541- 543.
[27] 陳曦, 吳尊曌, 董輝, 叢斌. 3地亞洲玉米螟越冬幼蟲化性與復蘇后發育歷期的關系. 江蘇農業科學, 2009, (2): 114- 115.
[28] 魯新, 劉宏偉, 汪洋洲, 丁巖, 張國紅, 李麗娟. 亞洲玉米螟化型的主要特性及其遺傳穩定性. 植物保護學報, 2009, 36(2): 183- 184.
[29] 劉寧, 文麗萍, 何康來, 王振營, 趙廷昌. 不同地理種群亞洲玉米螟抗寒力研究. 植物保護學報, 2005, 32(2): 163- 168.
[30] 涂小云, 夏勤雯, 陳超, 陳元生, 匡先鉅, 薛芳森. 亞洲玉米螟體重和體型的地理變異. 昆蟲學報, 2011, 54(2): 143- 148.
[31] 涂小云, 陳元生, 夏勤雯, 陳超, 匡先鉅, 薛芳森. 亞洲玉米螟成蟲壽命與繁殖力的地理差異. 生態學報, 2012, 32(13): 4160- 4165.
[32] 涂小云, 陳元生, 夏勤雯, 陳超, 匡先鉅, 薛芳森. 亞洲玉米螟兩個不同地理種群對暗期干擾的滯育應. 昆蟲學報, 2011, 54(8): 943- 948.
[33] 魯新, 田志來, 張國紅, 李麗娟. 不同化性亞洲玉米螟有效積溫和成蟲飛翔能力的比較. 植物保護學報, 2005, 32(3): 333- 334.
[34] 夏新, 叢斌, 宋立秋, 劉洪敏. 不同地理種群和不同化性亞洲玉米螟的有效積溫和存活率的研究. 安徽農業科學, 2007, 35(19): 5795, 5797.
[35] 王桂清, 忻亦芬. 沈陽地區不同化性亞洲玉米螟歷期和有效積溫的研究. 沈陽農業大學學報, 2000, 31(5): 444- 447.
[36] 喬利, 鄭堅武, 成衛寧, 李怡萍. 不同飼料配方對亞洲玉米螟生長發育和繁殖的影響. 西北農林科技大學學報: 自然科學版, 2008, 36(5): 109- 112.
[37] Danilevskii A S. Photoperiodism and Seasonal Development of Insect. Edinburgh and London: Oliver and Boyd, 1965.
[38] Honěk A. Geographical variation in thermal requirements for insect development. European Journal of Entomology, 1996, 93(3): 303- 312.
[39] 黃麗莉, 闕海勇, 陳超, 匡先鉅, 薛芳森. 亞洲玉米螟不同地理種群基于COⅠ基因序列初步分析. 應用昆蟲學報, 2012, 49(6): 1508- 1512.
[40] 楊瑞生, 王振營, 何康來, 白樹雄. 稈野螟屬部分種的線粒體COⅡ基因序列分析及其分子系統學(鱗翅目: 草螟科). 昆蟲學報, 2008, 51(2): 182- 189.
[41] 李菁, 張穎, 王振營, 何康來, 王強. 基于線粒體DNA COⅡ基因的亞洲玉米螟中國不同地理種群遺傳分化及基因流研究. 昆蟲學報, 2010, 53(10): 1135- 1143.
Geographic variation in developmental duration of the Asian corn borer,Ostriniafurnacalis(Guenée) (Lepidoptera: Crambidae) in China
TU Xiaoyun1, 2, XIA Qinwen2, CHEN Chao2, HE Haimin2, XUE Fangsen2,*
1CollegeofLifeSciences,JiangxiNormalUniversity,Nanchang330022,China2InstituteofEntomology,JiangxiAgricultureUniversity,Nanchang330045,China
Insect species with broad geographic distributions can phenologically adjust their life cycles to adapt to different habitats, for example by altering their growth, development, and reproduction periods to better match their emergence times with local conditions, such as food availability, optimum temperature, rainfall, etc.). These adjustments are very important for such widespread species to be able to reproduce in different habitats. The Asian corn borer,Ostriniafurnacalis(Guenée) (Lepidoptera: Crambidae) is a major pest insect ofZeamaysL., that poses a serious economic threat to corn production in China. Its distribution ranges from north to south in China, through Southeast Asia, and into Australia, and the pest has recently been found in isolated regions of the United States. Because of its wide distribution, the Asian corn borer encounters great variation in climatic conditions across its range and exhibits considerable diversity in its life-history parameters among geographic populations. Populations differ distinctly in voltinism, critical photoperiod, post-diapause emergence time, body size, fecundity, and cold hardiness. However, little is known about whether the developmental duration ofO.furnacalispopulations varies with geographic origin. In the present study, we systematically investigated the developmental duration of the egg, larval, and pupal phases at five different temperatures (20, 22, 25, 28 and 31℃) using insects collected from five different populations of the Asian corn borer. Each population inhabited a different latitude: Yangshuo, Guangxi (YS, 24.8°N, 110.5°E); Nanchang, Jiangxi (NC, 28.8°N, 115.9°E); Taian, Shandong (TA, 36.2°N, 117.1°E); Langfang, Hebei (LF, 39.5°N, 116.7°E) and Haerbin, Heilongjiang (HEB, 44.9°N, 127.2°E). Although at any given temperature the duration of the egg phase did not differ significantly among populations, the length of this phase increased slightly from south to north at 20, 22, and 25℃ and had a highly significant positive correlation with the latitudinal origin of the populations at these three temperatures. However, the egg phase lasted about the same amount of time in all populations at 28℃, with no significant correlation to latitudinal origin. The duration of the larval phase was not significantly different among the five populations at 31℃, but it differed significantly at 20, 22, 25, and 28℃. The larval-phase duration had a significant positive correlation with the latitudinal origin of the populations at 22, 25, and 28℃. However, these two parameters were negatively correlated at 20 and 31℃. The duration of the larval phase varied little with temperature in the HEB population, but it decreased significantly as temperature increased in the YS, NC, TA, and LF populations. The developmental durations of female pupae at 20℃, male pupae at 28℃, and all pupae at 31℃ did not differ significantly among populations, but all other pupal categories had significantly different durations among the five populations. The pupal-phase duration had a positive relationship with the latitudinal origin of the population at any given temperature. These results indicate that a single widespread species of insect may adapt to varying environmental conditions across its geographic range by altering the developmental duration of its life-history stages in different ways in each population.
Ostriniafurnacalis; geographic variation; developmental duration; egg; larva; pupa
國家自然科學基金項目(31260430);國家自然科學基金項目(31360327)
2013- 03- 26;
日期:2014- 03- 25
10.5846/stxb201303260512
*通訊作者Corresponding author.E-mail: xue_fangsen@hotmail.com
涂小云,夏勤雯,陳超,何海敏,薛芳森.中國亞洲玉米螟發育歷期的地理變異.生態學報,2015,35(2):324- 332.
Tu X Y, Xia Q W, Chen C, He H M, Xue F S.Geographic variation in developmental duration of the Asian corn borer,Ostriniafurnacalis(Guenée) (Lepidoptera: Crambidae) in China.Acta Ecologica Sinica,2015,35(2):324- 332.
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
