苯并[α]芘脅迫對三角褐指藻抗壞血酸-谷胱甘肽循環活動的影響?
2015-03-31 03:46:55潘魯青
中國海洋大學學報(自然科學版) 2015年12期
牛 曉, 李 赟, 潘魯青
(中國海洋大學海水養殖教育部重點實驗室, 山東 青島 266003)
?
苯并[α]芘脅迫對三角褐指藻抗壞血酸-谷胱甘肽循環活動的影響?
牛 曉, 李 赟??, 潘魯青
(中國海洋大學海水養殖教育部重點實驗室, 山東 青島 266003)
以三角褐指藻 (Phaeodactylumtricornutum)為材料,在連續4d內,以100、400、700μg/L 3個劑量的苯并[α]芘(B[α]P)對三角褐指藻進行脅迫試驗,分析苯并[α]芘對三角褐指藻的毒性效應,測定三角褐指藻的抗壞血酸-谷胱甘肽(AsA-GSH)循環非酶成分、含量及酶活性。研究表明:隨著B[α]P濃度的提高和脅迫時間的延長,B[α]P對三角褐指藻的生長抑制作用逐漸增強;藻細胞內丙二醛(MDA)含量及總抗氧化能力(TAOC)顯著升高,兩者與脅迫強度的相關性系數分別為0.960和0.894;同時,AsA-GSH循環中的非酶成分抗壞血酸(AsA)、谷胱甘肽(GSH)含量增加,相關酶谷胱甘肽還原酶(GR)、抗壞血酸過氧化物酶(APX)、脫氫抗壞血酸還原酶(DHAR)和單氫抗壞血酸還原酶(MHAR)活性普遍增加,其中,GSH含量、GR、APX和DHAR活性與藻細胞內MDA含量相關性系數分別為0.744、0.852、0.652和0.600。結果表明:AsA-GSH循環是三角褐指藻細胞對抗苯并芘脅迫、猝滅活性氧、降低膜脂過氧化的重要方式;細胞內AsA含量、GR活性及APX活性對苯并芘脅迫響應迅速、變化顯著,是潛在的生物指示物。
三角褐指藻; 苯并[α]芘脅迫; 抗壞血酸-谷胱甘肽循環組分
由于工業的發展及人類活動,多環芳烴類的有機污染物廣泛分布在包括海洋在內的多種區域,這些污染物不僅對環境中的生物多樣性造成直接威脅,它們還通過食物鏈富集并傳遞進而對人類的健康造成傷害。在這些受關注的PAHs污染物中苯并[α]芘(B[α]P)毒性最強,被國際癌癥調查組織確定為一級致癌物[1-2],是環境監測的目標污染物之一。
有關B[α]P對海洋生物毒性的研究已涉及到魚類[3]、蝦蟹類[4]、貝類[5]及浮游生物[6]等,有關B[α]P脅迫生物生理生化方面的研究也已有不少報道,這些研究表明, B[α]P誘導的氧化損傷是導致毒性傷害的重要因素。
光合細胞內存在一套完整的抗氧化體系。光合細胞中產生的活性氧自由基(ROS)一般首先經超氧化物歧化酶(SOD)轉化為H2O2,產生的H2O2再分別經過氧化氫酶(CAT)、谷胱甘肽過氧化物酶(GPX)、芬頓反應(Fenton reaction)及AsA-GSH循環等幾種方式清除。其中, AsA-GSH循環廣泛分布于細胞質基質、線粒體、葉綠體和過氧化物酶體之中,由3個氧化還原對和4種酶組成,在猝滅H2O2方面具有極為關鍵的作用[7]。目前,已有研究分別在重金屬脅迫的綠豆[8]和淡水藻類[9]、干旱的小麥[10]、漬水的康乃馨[11]和蘋果[12]、紫外線照射的藻類[13]、鹽度脅迫的小麥[14]和黃瓜[15]以及熒蒽暴露的浮萍[16]上進行了AsA-GSH循環相關組分的變化情況。上述研究表明,各類脅迫均可以造成細胞氧化損傷,從而顯著誘導AsA-GSH循環活動增強。但在微藻上AsA-GSH循環的活動還未見報道,雖然B[α]P對海洋自養生物的毒性已有不少報道,但這些生物在B[α]P脅迫期間AsA-GSH相關成分的變化情況也仍屬未知。
本實驗以三角褐指藻(Phaeodactylumtricornutum)為材料,在比較苯并[α]芘對三角褐指藻生長、脂質過氧化及抗氧化水平影響的基礎上,同時分析B[α]P脅迫下藻細胞內AsA-GSH循環非酶成分含量及相關酶活性的變化,以期了解三角褐指藻AsA-GSH循環在清除體內H2O2的可能作用,為探討三角褐指藻對B[α]P等有機毒物的解毒機制提供依據。
1 材料與方法
1.1 藻種和培養條件
實驗以中國海洋大學微藻種質庫三角褐指藻為實驗材料。三角褐指藻培養基為f/2培養基,250mL錐形瓶培養,培養基體積200mL,三角褐指藻起始細胞濃度5×105個/mL,培養溫度為20℃,光照強度為冷熒光燈30μmol·m-2·s-1,光∶暗比為13∶11[17-18]。
1.2 實驗梯度設置
實驗所用B[α]P為Dr. Ehrenstorfer GmbH公司產品,純度為99.6%。使用二甲基亞砜(DMSO)作為助溶劑,制成B[α]P儲備液,儲備液在海水中的最大溶解度為10mg/L。經預實驗,0.01%的DMSO對三角褐指藻的生長沒有顯著抑制效應,實驗中三角褐指藻培養液中DMSO濃度小于0.01%。
實驗B[α]P濃度設置根據實驗中三角褐指藻的IC50,并結合在另一種海洋硅藻假微型海鏈藻[19]上的研究,設置B[a]P的3個濃度梯度為100、400、700μg/L,每個濃度處理設置3個平行,連續處理96h。本實驗以B[α]P濃度與處理時間的乘積作為B[α]P的脅迫強度。
1.3 藻密度變化及抑制率的測定
在暴露處理過程中,每24h取1mL培養液,在顯微鏡(Nikon HFXⅡ)下用血球計數板統計三角褐指藻細胞密度,基于細胞密度的變化,與對照組相比按如下公式計算百分抑制率(ASTM):
I=(C-X)/C×100%。
式中:I為百分抑制率;C為對照組藻細胞數目;X為處理組藻細胞數目。
1.4 主要生理生化指標的測定
1.4.1 脂質過氧化水平的測定 脂質過氧化水平以測定次級氧化產物MDA含量的方法來反映。MDA含量測定根據Wills[20]的方法稍加改進。單位為nmol·mg-1prot-1。
1.4.2 總抗氧化能力的測定 使用TAOC分析試劑盒(南京建成生物工程研究所,南京)測定總抗氧化能力,單位為U/mg prot。
1.4.3 抗壞血酸和還原型谷胱甘肽含量測定 抗壞血酸含量測定方法參照Kampfenkel等[21]的方法并稍作改進。AsA含量單位為mmol/g鮮重。GSH含量的測定參照Anderson[22]的方法。GSH含量單位為mmol/g鮮重。
1.4.4 抗壞血酸-谷胱甘肽循環酶活測定 參照Ma等[23]和Knorzer等[24]所描述的方法提取抗氧化酶液。隨后分別測定相關酶活性。其中,抗壞血酸過氧化物酶(APX,EC1.11.1.11)活性根據Nakano和AsAda[25]所描述的方法測定。以μmolAsA ·min-1·mg-1prot-1表示APX活性。單脫氫抗壞血酸還原酶(MDAR,EC1.6.5.4)活性根據Miyake和AsAda[26]所述的方法進行測定。以μmolNADH·min-1·mg-1prot-1表示MDAR活性。脫氫抗壞血酸還原酶(DHAR,EC1.8.5.1)活性根據Dalton等[27]所述的方法進行測定。以nmolAsA·min-1·mg-1prot-1表示DHAR活性。谷胱甘肽還原酶(GR,EC1.6.4.2)活性根據Grace和Logan[28]所述的方法進行測定。以nmolNADPH·min-1·mg-1prot-1表示GR活性。總的可溶性蛋白濃度測定依據Bradford[29]的方法并稍作改進,使用牛血清蛋白作為標準蛋白,根據標準曲線求得蛋白含量。
1.5 數據統計分析
所有數據均以3個平行組數據的平均值±標準差(Mean±S.D.)表示, 采用SPSS 17.0 軟件進行單因素方差分析(ANOVA)、Duncan 檢驗法及Pearson相關性分析,置信度0.05,差異顯著用不同字母表示。
2 結果與分析
2.1 B[α]P對三角褐指藻生長的影響
基于B[α]P對三角褐指藻的毒性,本試驗設置的B[α]P脅迫濃度均屬于亞致死濃度。結果如圖1所示,試驗期間3個處理組微藻細胞密度均呈連續增加狀態,但與對照相比,B[α]P處理僅24h,高濃度400和700μg/L組三角褐指藻生長較對照組受到明顯抑制,48h后,低濃度100μg/L組三角褐指藻生長也受到明顯抑制。脅迫至96h,3個處理組三角褐指藻生長均有所恢復,3個處理組96h抑制率從低濃度組到高濃度組依次為16.7%、41.4%和43.1%。
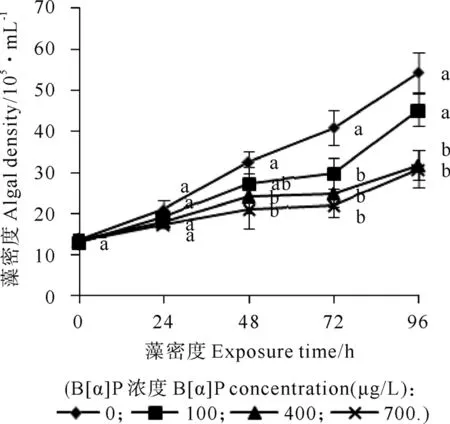
圖1 三角褐指藻在不同濃度B[α]P 脅迫下的生長曲線Fig.1 Growth curves of Phaeodactylum tricornutum in different concentration of B[α]P
2.2 B[α]P對三角褐指藻脂質過氧化水平及抗氧化能力的影響
與各處理組微藻生長相對應,三角褐指藻體內的MDA含量隨著處理濃度的增大及處理時間的延長而逐漸增高(見圖2A),如B[α]P脅迫48h,100μg/L處理組微藻MDA含量一直未顯著增高,400和700μg/L處理組藻細胞的MDA的含量均顯著高于對照,700μg/L處理組MDA含量則顯著高于400μg/L處理組。脅迫72h,3個處理組微藻MDA含量均顯著高于對照,其中700μg/L處理組MDA 含量仍顯著高于100μg/L和400μg/L處理組。脅迫延至96h,100μg/L組微藻MDA含量雖較72h有所降低,仍顯著高于對照組水平,但顯著低于400μg/L組和700μg/L組。結果表明,B[α]P處理與三角褐指藻細胞MDA含量存在明顯的時間-劑量效應。相關分析表明,B[α]P脅迫強度與藻細胞體內MDA含量相關性系數為0.96,相關性顯著(P<0.05),這說明B[α]P脅迫是藻細胞膜脂過氧化水平增加的直接原因。
3個處理組三角褐指藻TAOC的分析顯示(見圖2B),與藻細胞MDA含量的變化相似,脅迫48h內,隨著處理時間的延長,400和700μg/L處理組微藻抗氧化能力逐漸增強。脅迫72h,100μg/L組微藻的抗氧化能力也顯著高于對照,但低于400和700μg/L處理組。脅迫至96h,3個處理組微藻的TAOC均顯著高于對照組,各處理組間的TAOC也隨著B[α]P處理濃度的升高而呈顯著增高,此時,700μg/L處理組微藻總抗氧化水平達最高水平,為對照組的167%。相關分析表明,三角褐指藻體內的總抗氧化能力與苯并芘脅迫強度及細胞內的MDA含量的相關性系數均為0.89,相關性顯著(P<0.05)

圖2 三個濃度苯B[α]P處理三角褐指藻MDA含量、TAOC水平變化
2.3 B[α]P暴露期間三角褐指藻抗壞血酸和谷胱甘肽含量的變化情況
作為微藻抗氧化能力的一部分,B[α]P處理過程中,三角褐指藻內AsA含量變化明顯(見圖3b),僅脅迫24h,400和700μg/L處理組AsA含量已分別是對照組水平的133%和154%,均顯著高于對照。脅迫48h, 100μg/L組微藻AsA含量也達對照組的147%,而此時,700μg/L組抗壞血酸含量已達到峰值,為對照的179%,顯著高于100和400μg/L處理組。脅迫至72h,400μg/L處理組被進一步誘導而顯著高于其它處理組,而100μg/L組抗壞血酸含量已與對照組水平相當,700μg/L組三角褐指藻的抗壞血酸含量也有所降低,但仍高于對照組水平。結果顯示,AsA含量對B[α]P脅迫相當敏感,但相關分析表明,AsA含量與脅迫強度、細胞內MDA含量和總抗氧化能力相關系數分別為0.325、0.200、0.233,均無顯著的相關。
與AsA含量的變化不同,脅迫24h,3個處理組微藻的GSH含量與對照組均無顯著變化(見圖3a)。脅迫至48和72h,2個高濃度處理組微藻GSH含量均顯著高于對照,100μg/L組仍與對照組無顯著不同。脅迫延長至96h,100和700μg/L處理組微藻的GSH含量與72h時相應組的含量相似,400μg/L組GSH含量則被誘導而顯著高于所有處理組,此時,該處理組微藻GSH含量為對照組的155%。相關分析表明,微藻體內的GSH含量與B[α]P脅迫強度、體內MDA含量和抗氧化能力的相關系數分別為0.68、0.74和0.70(P<0.05),均為顯著相關。
2.4 B[α]P暴露對三角褐指藻抗壞血酸-谷胱甘肽循環相關酶活力的影響
2.4.1 APX活性變化 三角褐指藻APX活性對B[α]P處理相當敏感(見圖4a),處理24h,3個處理組APX活性即被顯著誘導,誘導效應與B[α]P濃度直接有關,400和700μg/L組的APX活性顯著高于100μg/L處理組。處理72h,700μg/L組APX活性已比對照組升高了33%,各處理組間APX的活性隨B[α]P處理濃度的升高而增高,并均達顯著水平。進一步處理至96h,100μg/L組APX活性降低至對照組水平,700μg/L組APX活性也有所降低,與400μg/L組APX活性水平相似,仍均顯著高于對照組。相關分析表明,微藻APX活性與B[α]P脅迫強度、MDA含量和總抗氧化能力相關性系數分別為0.70、0.65和0.54(P<0.05),達顯著相關水平。
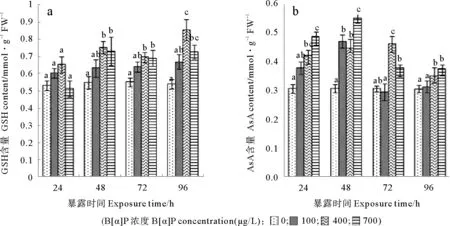
圖3 三個濃度B[α]P處理三角褐指藻AsA含量、GSH含量變化
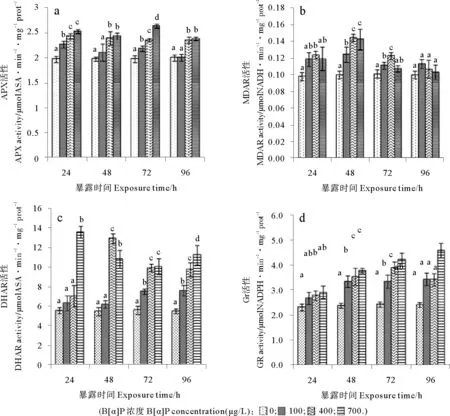
圖4 三種濃度苯并[α]芘處理三角褐指藻APX、MDAR、DHAR及GR活性變化Fig.4 The effect of BaP in three content on APX,MDAR,DHAR,GR activity of Phaeodactylum tricornutum
2.4.2 MDAR活性變化 MDAR活性變化(見圖4b),處理24h,雖然3個處理組MDAR活性均被誘導增高,但只有400μg/L組的MDAR活性與對照有顯著差異。處理至48h,3個處理組MDAR活性均顯著高于對照,此時,400和700μg/L組MDAR活性分別較對照組提高了44%和42%,顯著高于100μg/L處理組。隨后3個處理組的MDAR活性均呈現下降趨勢,72h時400μg/L組的MDAR活性仍顯著高于對照,至96h,3個處理組均與對照無顯著差異。相關分析表明,MDAR活性與B[α]P脅迫強度、MDA含量和總抗氧化能力分別為0.099、0.015、0.104,均無顯著相關。
2.4.3 DHAR活性的變化 如圖4c所示,B[α]P處理24h,高濃度700μg/L處理組DHAR活性被誘導,其活性是對照組的142%,其余2個處理組DHAR活性雖有增高,但與對照均無顯著差異。48h時,400μg/L組DHAR活力也迅速升高,達對照組的135%,700μg/L處理組DHAR活性有所下降,顯著低于400μg/L組。處理72h后,100μg/L組DHAR活性也顯著高于對照,但仍顯著低于其它2個處理組。持續處理96h, 3個處理組DHAR活性均顯著高于對照,處理組間DHAR活性隨著B[α]P處理濃度的增高而顯著升高。相關分析表明,DHAR活性與B[α]P脅迫強度、藻細胞MDA含量和總抗氧化能力相關性系數為0.68、0.60和0.63(P<0.05),達顯著相關水平。
2.4.4 GR活性的變化 如圖4d所示,處理24h,3個處理組的GR活性即顯著高于對照,B[α]P處理濃度越高,微藻的GR活性被誘導得越高。隨著處理時間延長,各處理組GR活性被進一步誘導,組間GR活性也隨B[α]P處理濃度的升高而顯著增高。至72h,400μg/L組的GR活性達峰值,而700μg/L組的GR活性仍繼續升高至處理96h時達最高。相關分析表明,GR活性與B[α]P脅迫強度、細胞內MDA含量和總抗氧化能力的相關性系數為0.89、0.85和0.93(P<0.05),均具有極高的相關性。
3 討論
AsA-GSH循環也稱作Halliwell-AsAda循環,該循環的非酶成分為3個氧化還原對,即還原型和氧化型的抗壞血酸(AsA/DHA)、還原型和氧化型的谷胱甘肽(GSH / GSSG)以及還原型和氧化型的煙酰胺腺嘌呤二核苷酸磷酸(NADPH / NADP+),4種相關酶為,消耗AsA用以還原H2O2的抗壞血酸過氧化物酶(APX)、合成AsA的單脫氫抗壞血酸還原酶(MDAR)和脫氫抗壞血酸還原酶(DHAR)以及生成GSH的GR。從抗氧化的角度來看,APX是直接的效應酶,其它3個酶與形成AsA或GSH有關。AsA是許多酶促反應或氧化還原反應的電子提供者,能直接猝滅超氧負離子和羥基自由基,也能夠再生α-生育酚來保護膜系統,降低膜脂過氧化的危害[30]。GSH存在于幾乎所有的細胞原件,因為其參與AsA-GSH循環、維持巰基(-SH)穩定以及作為GSTs底物[31],被視為細胞內對ROS引起的氧化損傷具有防御功能的關鍵物質。
已有研究顯示,隨著細胞內氧化脅迫加劇,膜脂過氧化程度增高,與AsA-GSH循環相關的非酶成分含量及相關酶活性多被誘導增高,如Nath等用砷脅迫水稻,發現根組織H2O2、MDA、 AsA和GSH含量普遍升高[32]。Zezulka等發現熒蒽可導致浮萍MDA含量升高,與此同時,浮萍的抗氧化水平呈誘導升高狀態,APX活性也受誘導而增強[16]。Anjum等發現Cd脅迫30d的綠豆內AsA-GSH循環的相關酶如GR、MDAR、DHAR活性均顯著增高[8]。Saher等研究發現康乃馨內MDA和過氧化物水平明顯增高時,APX、DHAR、MDAR、GR活性均被誘導增強[11]。另外在Cd脅迫的金魚藻[9],H2O2預處理的栽培稻種子[33],以及鹽度脅迫的魚腥藻內[34]均發現這些脅迫導致APX活性增強,可對持續性的脅迫起到一定的拮抗作用。也有些研究發現,在發生細胞氧化脅迫時,AsA-GSH循環相關成分的變化有時候并不完全一致,如Chakrabarty等在蘋果上的分析發現,氧化脅迫導致APX、MDAR和GR酶活性增強時,而DHAR活性并未出現誘導增強[12]。Abhay用鉛脅迫Talinumtriangulare,結果發現其AsA和GSH含量較對照組明顯降低[35]。Talaat發現高鹽脅迫導致菜豆生長抑制,此時菜豆體內GSH和AsA含量以及抗氧化酶活性也均低于對照組[36]。Shu等研究發現,鹽度脅迫導致黃瓜體內AsA及GSH含量降低,APX、DHAR和GR活性降低,而MDAR活性卻升高了[15]。
作者曾同時比較了B[α]P對7種海洋微藻的毒性,結果顯示三角褐指藻對B[α]P有相當高的耐受性[37]。本研究采用3個較低濃度的B[α]P處理三角褐指藻,旨在分析B[α]P導致三角褐指藻發生氧化性脅迫時,細胞內AsA-GSH循環相關成分的變化情況。結果顯示,本研究設計的3個處理濃度比較合適,試驗期間,各處理組隨著處理濃度的升高及處理時間的延長,藻細胞生長隨之表現不同程度的抑制,細胞內MDA含量及總抗氧化能力顯著增加,二者與B[α]P脅迫強度的相關性系數分別為0.96和0.89,均呈高度相關關系。
在藻細胞膜脂過氧化水平增高及總抗氧化能力被誘導時,三角褐指藻細胞內AsA-GSH循環的非酶成分含量增加,相關酶活均被顯著誘導,其中GR活性和GSH含量與體內MDA含量及總抗氧化能力、APX活性與體內MDA含量、DHAR活性與體內總抗氧化能力的相關性分別達到極顯著相關水平,APX活性與體內總抗氧化能力及DHAR活性與體內MDA含量分別達到顯著相關水平。雖然AsA含量及MDAR活性與膜脂過氧化程度及總抗氧化能力的相關性均未達顯著水平,但AsA對B[α]P脅迫相當敏感,含量的變化幅度最大,MDAR活性也隨著B[α]P脅迫強度的增大而呈現顯著的漲落。結果說明,AsA-GSH循環是三角褐指藻抵抗B[α]P脅迫、降低膜脂過氧化程度的重要方式。
相關性分析還顯示,三角褐指藻細胞AsA含量與MDAR活性的相關性最高,與DHAR活性及APX活性的相關性也達極顯著水平。APX活性增高,說明AsA的消耗在增加。MDAR和DHAR均參與AsA的合成,這2種酶活性的增高,表明AsA形成能力增強。MDAR和DHAR活性與AsA含量的相關性存在一定差異,似乎說明B[α]P脅迫期間,三角褐指藻內MDAR形成AsA的作用有所上升。GSH含量只與循環內GR活性顯著相關而非高度相關,似乎表明,盡管GR參與GSH的生成,但由于GSH在細胞內的功能比較豐富,其合成及消耗的方式也可能相當復雜,GSH含量并不只與GR活性有關。
進一步比較AsA-GSH循環各成分在B[α]P脅迫時的變化情況,結果顯示,對B[α]P脅迫快速做出變化的組分有AsA含量、GR活性和APX活性,而GSH含量、DHAR活性及MDAR活性的變化則是在B[α]P脅迫強度進一步增高時才被誘導增強,如AsA含量,僅脅迫24h, 400和700μg/L處理組AsA含量已顯著高于對照,脅迫48h, 100μg/L處理組AsA含量即達對照組的147%。而GSH含量,脅迫至48h,400和700μg/L處理組GSH含量才顯著高于對照, 100μg/L處理組的GSH含量一直與對照無顯著差異。這說明三角褐指藻AsA-GSH循環不同成分對B[α]P脅迫的反應存在明顯不同。相對而言, AsA含量、GR活性及APX活性對B[α]P脅迫的反應更為快速,可作為生物指示物用于生態環境安全的評價。
[1] IARC. Monographs on the Evaluation of Carcinogenic Risk of Chemicals to Man: Polycyclic Aromatic Hydrocarbons [M]. Lyon: IARC, 1983: 211-224.
[2] Straif K, Baan R, Grosse Y, et al. Carcinogenicity of polycyclic aromatic hydrocarbons [J]. Lancet Oncol, 2005, 6(12): 931-932.
[3] Huang L X, Zuo Z H, Zhang Y Y, et al. Use of toxicogenomics to predict the potential toxic effect of Benzo(a)pyrene on zebrafish embryos: Ocular developmental toxicity [J]. Chemosphere, 2014, 108: 55-61.
[4] Ren X Y, Pan L Q, Wang L. Metabolic enzyme activities, metabolism-related genes expression and bioaccumulation in juvenile white shrimpLitopenaeusvannameiexposed to benzo[a]pyrene [J]. Ecotoxicology and Environmental Safety, 2014, 104: 79-86.
[5] 潘魯青, 劉娜, 王靜. 櫛孔扇貝B[α]P脅迫下生物標志物篩選的研究 [J]. 水生生物學報, 2009, 36(2): 299-306.
[6] Carvalho R N, Burchardt A D, Sena F, et al. Gene biomarkers in diatomThalassiosirapseudonanaexposed to polycyclic aromatic hydrocarbons from contaminated marine surface sediments [J]. Aquatic Toxicology, 2011, 101(1): 244-253.
[7] Noctor G, Foyer, C H. Ascorbate and Glutathione: keeping active oxygen under control [J]. Annual Review of Plant Physiology and Plant Molecular Biology, 1998, 49: 249-279.
[8] Anjum N A, Umar S, Iqbal M, et al. Cadmium causes oxidative stress in mung bean by affecting the antioxidant enzyme system and ascorbate-glutathione cycle metabolism [J]. Russian Journal of Plant Physiology, 2011, 58(1): 92-99.
[9] Arvind P, Prasad MNV. Zinc alleviates cadmium-induced oxidative stress in Ceratophyllum demersum L: a free-floating freshwater macrophyte [J]. Plant Physiology and Biochemistry, 2003, 41(4): 391-397.
[10] Kang G Z, Li G Z, Liu G Q, et al. Exogenous salicylic acid enhances wheat drought tolerance by influence on the expression of genes related to ascorbate-glutathione cycle [J]. Biologia Plantarum, 2013, 57(4): 718-724.
[11] Saher S, Piqueras A, Hellin E, et al. Hyperhydricity in micropropagated carnation shoots: the role of oxidative stress [J]. Physiologia Plantarum, 2004, 120(1): 152-161.
[12] Chakrabarty D, Park S Y, Ali M B, et al. Hyperhydricity in apple ultrastuctural and physiological aspects [J]. Tree Physiology, 2005, 26(3): 377-388.
[13] Singh V P, Srivastava P K, Prasad S M. Differential effect of UV-B radiation on growth, oxidative stress and ascorbate-glutathione cycle in two cyanobacteria under copper toxicity [J]. Plant Physiology and Biochemistry, 2012, 61: 61-70.
[14] Rehman R U, Zia M, Abbasi B H, et al. Ascorbic Acid and Salicylic Acid Mitigate NaCl Stress in Caralluma tuberculata Calli [J]. Applied Biochemis try and Biotechnology, 2014, 173(4): 968-979.
[15] Shu S, Yuan L Y, Guo S R, et al. Effects of exogenous spermine on chlorophyll fluorescence, antioxidant system and ultrastructure of chloroplasts inCucumissativusL. under salt stress [J]. Plant Physiology and Biochemistry, 2012, 63: 209-216.
[16] Zezulka S, Kummerova M, Petr Babula, et al. Lemna minor exposed to fluoranthene: Growth, biochemical,physiological and histochemical chan ges [J]. Aquatic Toxicology, 2013, 140: 37- 47.
[17] Huang A Y, He L W, Wang G C. Identification and characterization of microRNAs fromPhaeodactylumtricornutumby highthroughput sequencing and bioinformatics analysis [J]. BMC Genomics, 2011, 12: 337.
[18] Sapriel G, Quinet M, Heijde M, et al. Genome-Wide Transcriptome Analyses of Silicon Metabolism inPhaeodactylumtricornutumReveal the Multilevel Regulation of Silicic Acid Transporters [J]. Plos One, 2009, 4(10): e7458.
[19] Bopp S K, Lettieri T. Gene regulation in the marine diatomThalassiosirapseudonanaupon exposure to polycyclic aromatic hydrocarbons (PAHs) [J]. GENE, 2007, 396(2): 293-302.
[20] Wills E D, Snell K, Mullock B. Evaluation of iipid perocidation in lipids and biological membranes. biochemical toxicology: a practical approach [M]. Wa shington: IRL Press, 1987: 127-152.
[21] KamPfenkel K, VanMontagu M, Inze D, et al. Extraction and determination of ascorbate and dehydroascorbate from Plant Tissue [J]. Analytical Biochemistry, 1995, 225(1): 165-167.
[22] Anderson M E. Determination of glutathione and glutathione disulfide in biological samples [J]. Methods Enzymol, 1985, 113: 548-550.
[23] Ma F W, Cheng L L. Exposure of the shaded side of apple fruit to full sun leads to up-regulation of both the xanthophylls cycle and the ascorbate-glutathioneeycle [J]. HortScience, 2004, 39(4): 887-887.
[24] Knorzer O C, Burner J, Boger P. Alterations in the antioxidative system of suspension-cultured soybean cells(Glycinemax)induced by oxidative stress [J]. Physiol Planlarum, 1996, 97(2): 388-396.
[25] Nakano Y, AsAda K. Hydrogen peroxide is scavenged by ascorbate-specifie peroxidase in spinach chloroplasts [J]. Plant Cell Physiol, 1981, 22: 867-880.
[26] Miyake C, AsAda K. Thylakoid-bound ascorbate peroxidase in spinach chloroplasts and photoreduction of its primary oxidation product monodehydroascorbate radicals in thylakoids [J]. Planl Cell Physiol, 1992, 33(5): 541-553.
[27] Dalton D A, Russell S A, Hanus F J, et al. Enzymatie recetions of ascothate and glutathione that prevent peroxide damage in soybean root nodules [J]. Proc Natl Acad Sci USA, 1986, 83: 3811-3815.
[28] Grace S C, Logan B A. Acclimation of foliar antioxidant systems to growth irradiance in three broad-leaved evergreen species [J]. Plant Physiol, 1996, 112(4): 1631-1640.
[29] Bradford M M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein dye binding [J]. Biochem, 1976, 72: 248-254.
[30] Gill S S, Tuteja N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants [J]. Plant Physiology and Biochemistry, 2010, 48(12): 909-930.
[31] Madhavarao K V, Raghavendra A S, Reddy K J. Photooxidative stress. in: Physiology and Molecular Biology of Stress Tolerance in Plants [M]. The Netherlands: Springer, 2006: 157-186.
[32] Nath S, Panda P, Mishra S, et al. Arsenic stress in rice: Redox consequences and regulation by iron [J]. Plant Physiology and Biochemistry, 2014, 80: 203-210.
[33] Hsu Y T, Kao C H. Heat shock-mediated H2O2accumulation and protection against Cd toxicity in rice seedlings [J]. Plant And Soil, 2007, 300(1): 137-147.
[34] Srivastava A K, Bhargava P, Rai L C. Salinity and copper-induced oxidative damage and changes in antioxidative defense system ofAnabaenadoliolum[J]. World Journal of Microbiology and Biotechnology, 2005, 21(6): 1291-1298.
[35] Kumar A, Majeti N V P. Proteomic responses to lead-induced oxidative stress inTalinumtriangulareJacq. (Willd. ) roots: identification of key biomarkers related to glutathione metabolisms [J]. Environmental Science and Pollution Research, 2014, 21(14): 8750-8764.
[36] Talaat N B. Effective microorganisms enhance the scavenging capacity of the ascorbate-glutathione cycle in common bean (PhaseolusvulgarisL. ) plants grown in salty soils [J]. Plant Physiology and Biochemistry, 2014, 80: 136-143.
[37] 沈忱, 李赟, 潘魯青. 苯并[α]芘對海洋微藻生長及細胞特征的影響 [J]. 海洋環境科學, 2012, 31(4): 511-514.
責任編輯 朱寶象
Effect of Benzo [α] Pyrene on the Ascorbate-Glutathione Cycle ofPhaeodactylumtricornutum
NIU Xiao, LI Yun, PAN Lu-Qing
(The Key Laboratory of Mariculture, Ministry of Education, Ocean University of China, Qingdao 266003, China)
In order to investigate the toxic effect of benzo [α] pyrene (B[α]P) onPhaeodactylumtricornutumand understand the variation of the ascorbate-glutathione cycle of the microalgal cells during B[α]P stress, the culture ofP.tricornutumwas exposed to three doses of B[α]P for different times (24, 48, 72 and 96 h) with the level of total antioxidant capacity (TAOC) , the activity of associating enzymes and the content of including malondialdehyde (MDA), ascorbate (AsA) and glutathione (GSH) determined. The results showed that the growth ofP.tricornutumwas seriously inhibited by B[α]P, while the content of MDA and the level of total antioxidant capacity(TAOC) increased significantly. The correlation of content of MDA and the level of TAOC with stress strength of B[α]P was 0.960 and 0.894, respectively. The content of AsA and GSH increased, and the activity of enzymes including glutathione reductase (GR), ascorbate peroxidase (APX), dehydroascorbate reductase (DHAR) and monodehydroascorbate reductase (MDAR) was all induced significantly. The correlation of the content of MDA with the activity of GR, APX, DHAR and the content of GSH was 0.744, 0.852, 0.652 and 0.600, respectively, which indicated that the ascorbate-glutathione cycle inP.tricornutumhas important protection function by scavenging reactive oxygen species to reduce lipid peroxidation from B[α]P stress. The activity of GR, APX and the content of AsA could be considered as microalgal sensitive biological indicator for further study because of their quick and significantly response to B[α]P stress.
Phaeodactylumtricornutum; B[α]P stress; AsA-GSH cycle
國家海洋局海洋公益項目(201105013-3)資助
2014-10-08;
2015-01-15
牛 曉(1988-), 男, 碩士生。E-mail: yaorinumber1@126.com
??通訊作者: E-mail: sxsdlwl@ouc.edu.cn
Q945.78
A
1672-5174(2015)12-030-08
10.16441/j.cnki.hdxb.20140315
