太湖大銀魚( Protosalanx chinensis) 細胞色素b 基因序列多態性分析
2015-04-02 07:57:28李大命張彤晴唐晟凱鐘立強
江蘇農業學報 2015年4期
李大命, 張彤晴, 唐晟凱, 鐘立強
( 江蘇省淡水水產研究所,江蘇省內陸水域漁業資源重點實驗室,江蘇 南京210017)
銀魚科( Salangidae) 魚類又稱銀魚( Icefish or Salangid) ,為一年生小型名貴魚類,具有很高的經濟價值和營養價值[1]。大銀魚( Protosalanx chinensis)隸屬于銀魚科,新銀魚亞科( Neosalanginae) ,大銀魚屬( Protosalanx) ,是銀魚科中個體最大的種,主要分布在朝鮮半島西海岸至甌江以北各水系及近海岸,系近海河口洄游或淡水定居魚類。大銀魚在中國水體中分布廣泛,且具有重要的漁業價值,曾為中國重要的出口創匯水產品。近30 多年來,由于水體生態環境污染、水利工程建設和過渡捕撈等因素,中國野生大銀魚的資源量嚴重下降[2],開展大銀魚資源保護已經迫在眉睫。
遺傳多樣性是評估資源現狀、制定野生種群保護和管理策略的重要前提[3-4]。有關大銀魚的研究主要集中在形態學、個體生態學和繁殖生物學等方面[2]。近年來,國內外學者對銀魚的遺傳學開展了一些研究,主要集中在銀魚的分子系統發育、遺傳多樣性和種群結構等方面。比如,羅宏偉等分析了三峽庫區大銀魚、太湖新銀魚和短吻間銀魚線粒體DNA 細胞色素b 基因序列多態性[5]; 郭立等采用4個線粒體基因片段研究了銀魚科魚類的系統發育[6];Zhang 等在長江流域利用cyt b 基因全序列對全世界17 種銀魚中的15 種銀魚屬間的分子系統發育關系進行了比較[7]; Zhao 等利用線粒體cyt b 基因全序列對淮河、長江以及珠江流域的太湖新銀魚13 個種群共計354 個個體的遺傳多樣性和種群結構進行了分析[8]。
太湖是中國第三大淡水湖泊,湖泊漁業是其最重要的功能之一[9-10]。在太湖漁業中,銀魚占有非常重要的地位,大銀魚占太湖銀魚漁業年總產量的50%以上,產品大都出口,創造了巨大的經濟效益[11]。國內學者對太湖大銀魚的胚胎發育、生長和食性等進行了研究[11-13],但對太湖大銀魚的遺傳學研究較少,僅見夏德全等采用RAPD 技術分析了太湖銀魚種群遺傳結構特征[14]。近10 多年來,則鮮有太湖大銀魚遺傳多樣性的研究報道。本試驗采用cyt b 基因作為分子標記,對太湖大銀魚的遺傳多樣性進行初步研究,為合理開發和利用大銀魚野生資源,以及保護太湖銀魚種質資源提供科學依據。
1 材料和方法
1.1 試驗材料
大銀魚于2013 年5 月采自太湖漫山水域,隨機選擇35 尾樣本,其體長在3.1 cm 至6.5 cm 之間。每個大銀魚樣本剪少量尾鰭條放入無水乙醇中固定,帶回實驗室保存備用。
1.2 DNA 提取
大銀魚樣本基因組DNA 的提取參照Sambrook等的方法[15]。采用分光光度計( Unico UV-4802H)和1%瓊脂糖凝膠電泳分別測定DNA 濃度和檢測DNA 質量。置于-20 ℃儲存備用。
1.3 PCR 擴增及測序
擴增cyt b 基因的引物為L14321 和H15634[7]。L14321 引物序列: 5'-CCAGTGACTTGAAAAACCACCG-3'; H15634 引物序列: 5'-CTTAGCTTTGGGAGTTAAGGGT-3',引物由上海生物工程有限公司合成。
PCR 反應體系總體積為50 μl,其中2×PCR Mix 25 μl ( 包括Taq 酶1.25 U,dNTPs 10 μmol,MgCl20.1 mmol ) ,上、下游引物(10 μmol/L) 各2 μl,DNA 模板2 μl,其余體積用超純水補足。PCR 反應程序為:94℃預變性4 min;94 ℃變性40 s,55 ℃退火40 s,72 ℃延伸90 s,30 個循環;72 ℃延伸10 min。
PCR 產物用1%瓊脂糖凝膠電泳檢測,對擴增效果好的PCR 產物進行割膠純化后,直接送上海生物工程有限公司進行測序,為保證所測序列的可靠性,所有樣品均采用雙向測序。
1.4 數據統計與分析
采用DNAstar[16]軟件對DNA 序列進行比對拼接,并輔以人工校對。用DnaSP[17]軟件計算多態位點數目( Np) 、單倍型數目( Nh) 、單倍型多樣性指數( h) 、核苷酸多態性( π) 和平均核苷酸差異數( k)等。用MEGA 4.1[18]軟件計算序列的堿基組成、變異位點和簡約信息位點。以太湖新銀魚( Neosalanx taihuensis) 為外群,采用鄰接法( Neighbor-Joining,NJ) 構建單倍型分子系統樹,并對各分支進行1 000次重復檢驗。利用Tajima’s D 中性檢驗分析大銀魚種群歷史上是否發生擴張。
2 結果
2.1 太湖大銀魚cyt b 基因堿基組成和變異分析
經校對獲得35 條cyt b 基因同源序列,其長度為1 141 bp,序列中堿基A、T、G 和C 的平均含量分別為21.7%、29.3%、16.7%和32.4%,A +T 的含量( 50.92%) 略高于G + C 的含量( 49.07%) 。在這1 141 bp 序列中,共檢測到13 個變異位點,占分析位點總數的1.14%。其中9 個為簡約位點,4 個單一多態位點( 表1) 。這些突變位點中有11 個轉換位點和2 個顛換位點,沒有插入和丟失位點。
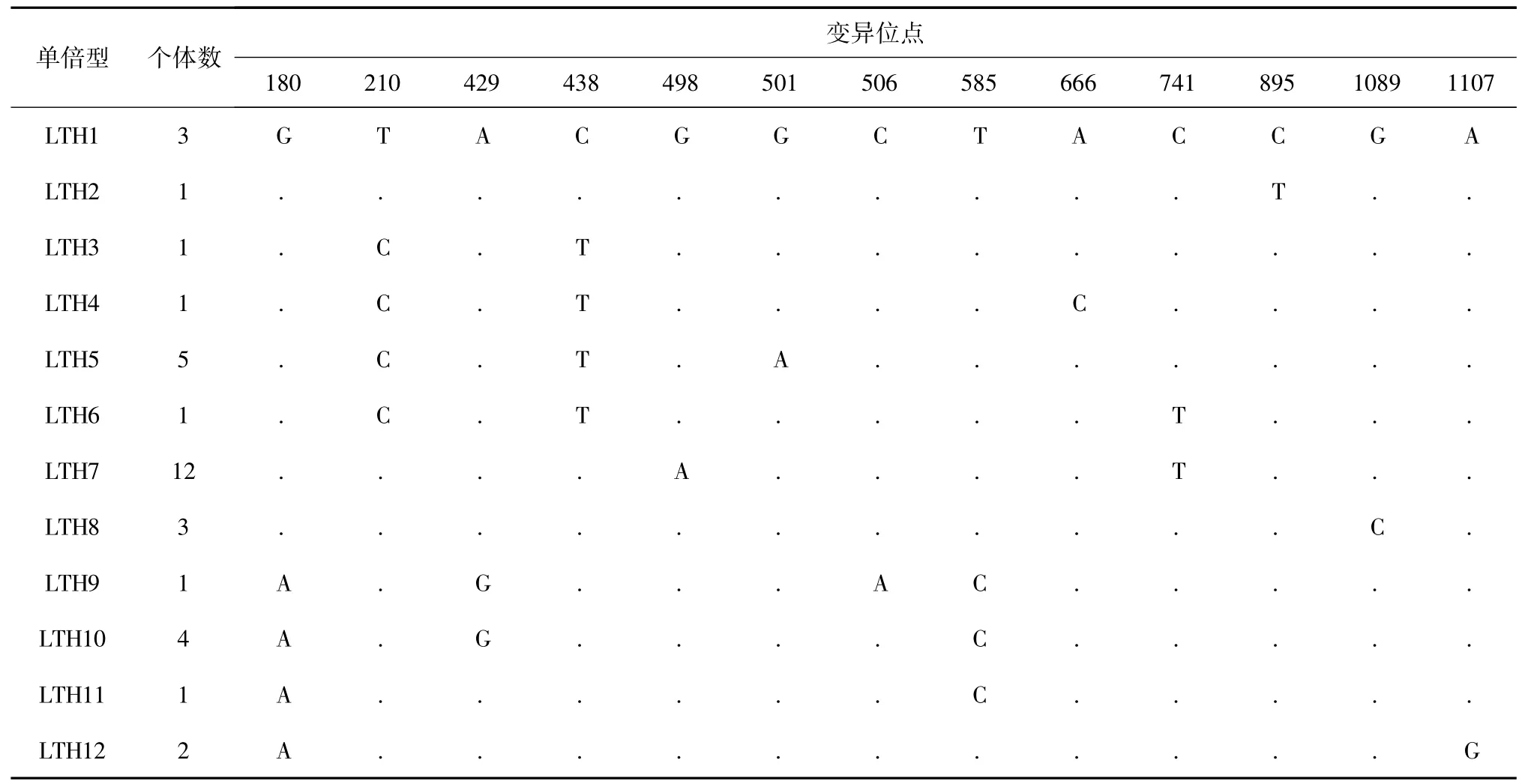
表1 太湖大銀魚mtDNA cyt b 基因單倍型多態性位點分布Table 1 Distribution of polymorphic sites in 12 haplotypes of Protosalanx chinensis with mtDNA cyt b in Lake Taihu
2.2 太湖大銀魚種群的遺傳多樣性
35 個大銀魚個體共定義12 個單倍型( LTH1 ~LTH12) ,其中單倍型LTH7 個體數最多,為12 個( 表1) 。大銀魚種群單倍型多樣性指數和核苷酸多樣 性 指 數 分 別 為0.850 ± 0.045 和0.002 96 ±0.000 17,平均核苷酸差異數為3.378,具有高單倍型多樣性和低核苷酸多樣性的特點( 表2) 。中性檢驗Tajima’s D 值為正值(0.224 52) ,且差異不顯著( P >0.10) ,表明太湖大銀魚種群比較穩定,沒有經歷過種群擴張。

表2 太湖大銀魚種群的遺傳多樣性參數Table 2 Parameter of genetic diversity of P. chinensis population in Lake Taihu
2.3 cyt b 單倍型聚類分析
以太湖新銀魚為外群,采用臨接法( NJ) 構建太湖大銀魚12 個單倍型的分子系統樹( 圖1) 。由圖1 可以看出,12 個大銀魚cyt b 單倍型聚為一支,太湖新銀魚單獨一支。單倍型間的遺傳距離較小,最大值為0. 006,平均遺傳距離為0. 003( 表3) ,表明單倍型之間沒有出現遺傳分化。
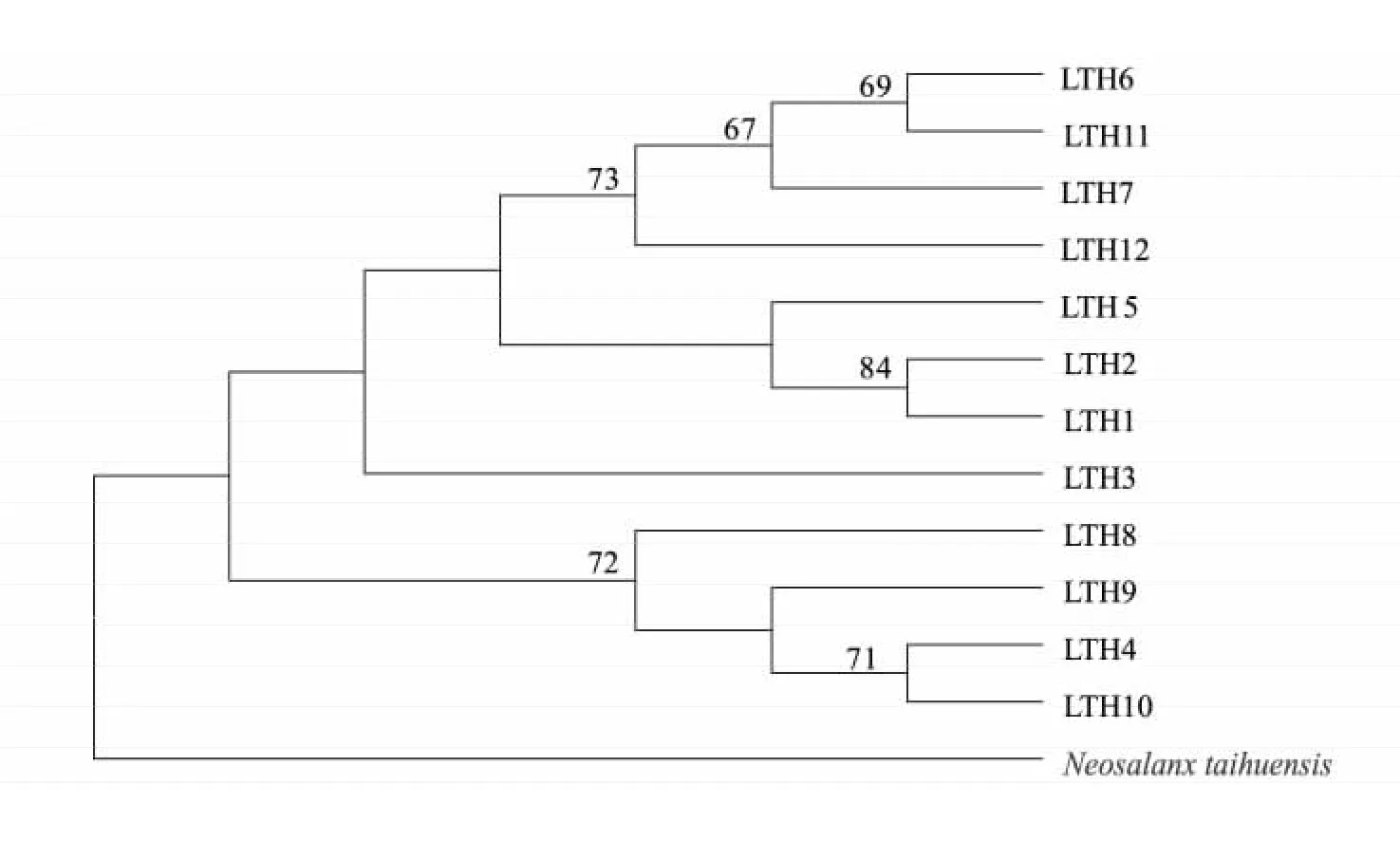
圖1 大銀魚cyt b 單倍型的NJ 系統樹Fig.1 The phylogenetic tree of mtDNA cyt b haplotypes of Protosalanx chinensis
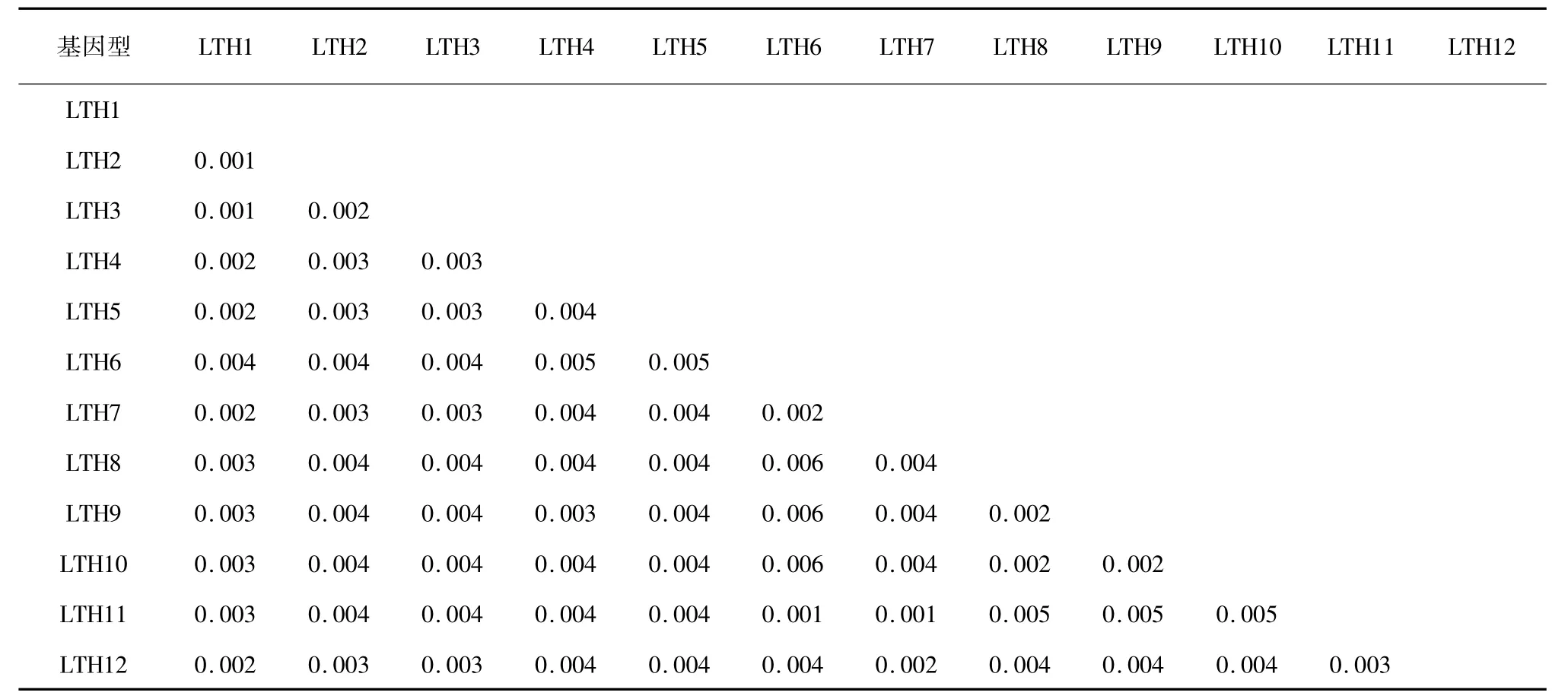
表3 太湖大銀魚cyt b 基因序列12 個單倍型遺傳距離(Kimura-2-parameter 法)Table 3 Genetic distance between 12 haplotypes of P. chinensis with cyt b sequences (Kimura-2-parameter method)
3 討論
線粒體DNA( mtDNA) 具有嚴格的母系遺傳,進化速率快,幾乎無重組等特點,被廣泛用于魚類系統發育和群體遺傳研究,其中細胞色素b 基因( cyt b)為重要的蛋白質編碼基因,其進化速率適中,能夠有效反映亞種、種群、屬等不同水平的遺傳信息[19]。本研究獲得了35 條太湖大銀魚cyt b 基因同源序列(1 141 bp) ,堿基G( 16.71%) 的含量最低,具有明顯的反G 偏倚,顯示出cyt b 基因的共同特征[20]。共檢測到13 個變異位點,沒有插入或丟失位點,其中11 個轉換位點,2 個顛換位點,結果符合同種魚類中轉換在數量上遠超過顛換的特征[21]。
遺傳多樣性是指生物種內和種間的遺傳變異度,是生物進化與適應環境的基礎[22]。單倍型多樣性指數和核苷酸多樣性指數是衡量一個物種或群體遺傳多樣性的2 個重要指標。本研究中,太湖大銀魚的單倍型多樣性指數和核苷酸多樣性指數分別為0.850 ±0.045 和0.002 96 ±0.000 17,呈現出高單倍型多樣性和低核苷酸多樣性的特征。有研究者通過對線粒體DNA 序列的遺傳變異分析,將不同單倍型多樣性和核苷酸多樣性間的組合分成4 種類型:第1 種類型是較低的單倍型多樣性( h <0.5) 與較低的核苷酸多樣性( π <0.005) ; 第2 種類型是高單倍型多樣性與低核苷酸多樣性; 第3 種類型是低單倍型多樣性與高核苷酸多樣性; 第4 種類型是高單倍型多樣性與高核苷酸多樣性[23]。本研究中大銀魚種群的結果屬于第2 種類型,該類型是由于經過瓶頸效應后伴隨著迅速的種群增長與突變的積累造成,快速的種群增長有利于提高對新突變的保持力而導致核苷酸多樣度降低。一方面由于過度捕撈、環境污染等因素,在上世紀80 年代太湖大銀魚種群數量急劇下降,遺傳多樣性降低,即受到遺傳瓶頸效應的影響;另一方面,大銀魚生長快、繁殖周期短,能迅速擴大種群數量,有利于積累更多突變。因此太湖大銀魚種群遺傳多樣性具有較高的單倍性多樣性和較低的核苷酸多樣性特點。另外,通過比較本研究太湖大銀魚遺傳多樣性參數與文獻報道[7,11,24]的研究結果,可以得出太湖大銀魚種群的遺傳多樣性最高。因此,保護太湖大銀魚種質資源對于保護中國銀魚種質資源的基因遺傳多樣性具有重要意義。
從單倍型間的遺傳距離和NJ 系統樹可以看出,太湖大銀魚單倍型間的遺傳距離非常低,所有單倍型聚為一支,表明大銀魚種群內沒有明顯的遺傳分化,這與夏德全等[14]對太湖大銀魚種群的遺傳結構分析結果是一致的,太湖大銀魚種群內、種群間的遺傳相似性指數及遺傳距離都十分相近,這可能與歷史上大銀魚可能受到較為嚴重的遺傳瓶頸效應的影響有關。但需要注意的是,不同的cyt b 單倍型出現的頻率有較大差異,單倍型LTH7 個體數有12 個,但6 個單倍型( LTH2、LTH3、LTH4、LTH6、LTH9 和LTH11) 僅有1 個個體出現,這一方面可能是大銀魚種群進化過程中自然選擇的結果,也可能受到近些年來太湖較大規模大銀魚增殖放流所帶來的選擇效應的影響。假如一旦這些數量較少的單倍型失去,會降低大銀魚的遺傳多樣性。因此,在增值放流之前,有必要對增殖放流群體與野生群體進行遺傳多樣性和種群結構進行對比研究,從而避免增殖放流可能帶來的不利影響。
物種的遺傳多樣性高低與其適應能力、生存能力和進化潛力密切相關。豐富的遺傳多樣性意味著較高的適應生存潛力,蘊藏著較大的進化潛能、豐富的育種和遺傳改良潛力。物種遺傳變異的減少可能導致嚴重的后果,如成活率、生長與繁殖效率變低,降低種群個體對環境變化的適應能力。本研究結果表明目前太湖大銀魚有較高的單倍型多樣性,但核苷酸多樣性較低。同時大銀魚的種群波動劇烈,對棲息水環境質量要求很高。因此,為保護和合理利用太湖大銀魚資源,我們建議加大太湖環境保護力度,改善水環境質量、建立大銀魚種質資源保護區,保護區內禁止一切人類活動、控制銀魚捕撈強度和縮短捕撈時間等措施,以實現太湖大銀魚漁業的可持續發展。
[1] 張 潔. 銀魚科魚類及其物種多樣性保護[J]. 生物學通報,2008,43(6) :4-6.
[2] 王忠鎖,傅萃長,雷光春. 中國銀魚的多樣性及其保護對策[J]. 生物多樣性,2002,10(4) :416-424.
[3] AVISE J C. A role for molecular genetics in the recognition and conservation of endangered species[J]. Trends in Ecology and Evolution,1989,4: 279-281.
[4] WAITS L. Mitochondrial DNA phylogeography of the North American brown bear and implications for conservation[J]. Conservation Biology,1989,13: 408-417.
[5] 羅宏偉,段辛斌,王 珂,等. 三峽庫區3 種銀魚線粒體DNA細胞色素b 基因序列多態性分析[J]. 淡水漁業,2009,39(6) : 16-21.
[6] 郭 立,李 雋,王忠鎖,等. 基于四個線粒體基因片段的銀魚科魚類系統發育[J]. 水生生物學報,2011,35(3) : 449-459.
[7] ZHANG J,LI M,XU M Q,et al. Molecular phylogeny of icefish Salangidae based on complete mtDNA cytochrome b sequences,with comments on estuarine fish evolution[J]. Biological Journal of the Linnean Society,2007,91(2) : 325-340.
[8] ZHAO L,ZHANG J,LIU Z J,et al. Complex population genetic and demographic history of the Salangid,Neosalanx taihuensis,based on cytochrome b sequences[J]. BMC Evolution Biology,2008,8: 201-219.
[9] 谷孝鴻,白秀玲,江南,等. 太湖漁業發展及趨于設置與功能定位[J]. 生態學報,2006,26 (7) : 2247-2254.
[10] 周 毅,石建華,錢 偉,等. 太湖藻類分布及其對魚類的毒性試驗[J]. 江蘇農業科學,2013,41(4) :311-313.
[11] 張開翔. 大銀魚胚胎發育的觀察[J]. 湖泊科學,1992,4(2) :25-37.
[12] 王玉芬,蔣全文. 太湖大銀魚生長特性的研究[J]. 湖泊科學,1992,4(1) : 56-62.
[13] 朱成德. 太湖大銀魚生長與食性的初步研究[J]. 水產學報,1985,9(3) : 275-287.
[14] 夏德全,曹 螢,吳婷婷,等. 太湖中大銀魚、太湖新銀魚和寡齒新銀魚群體的遺傳結構[J]. 水產學報,1999,23(3) : 254-260.
[15] SAMBROOK J,RUSSELL D W. Molecular cloning: A laboratory manual[M]. 3 rd ed. New York: Cold Spring Harbor Laboratory Press,2002: 463-469.
[16] BURLAND T G. DNASTAR's Laser gene sequence analysis software[J]. Methods in Molecular biology,2000,132: 71-91.
[17] ROZAS J,SACHEZ-DELBARRIO J C,Messeguer X,et al.DnaSP,DNA polymorphism analyses by the coalescent and other m ethods [J]. Bioinformatics,2003,19 ( 18) : 2496-2497.
[18] TAMURA K,DUDLEY J,Nei M,et al. MEGA 4: molecular evolutionary genetics analys is ( MEGA ) softw are version 4.0[J].Molecular Biology and Evolution,2007,10: 1092-1093.
[19] GRAY M W,BURGER G,LANG B F. Mitochondrial evolution[J]. Science,1999,283 (5407) : 1476-1481.
[20] ROGERS A R,HARPENDING H. Population growth makes waves in the distribution of pairwise genetic differences[J]. Molecular Biology and Evolution,1992,9(3) :552-569.
[21] KOHER T D,THOMAS W K,MEYER A,et al. Dynamics of mitochondrial DNA evolution in an imals: Amplification and sequencing with conserved primers[J]. Proceedings of the National Academy of Sciences,1989,86 (16) : 6196-6200.
[22] 施立明. 遺傳多樣性及其保存[J]. 生物科學信息,1990,2(1) : 158-164.
[23] GRANT W S,BOWEN B W. Shallow population histories in deep evolutionary lineages of marine fishes: insights from sardines and anchovies and lessons for conservation[J]. Journal of Heredity,1998,89: 415-426.
[24] 趙 亮,張 潔,劉志謹,等. 喬氏新銀魚基于細胞色素b 序列的種群遺傳結構和種群歷史[J]. 生物多樣性,2010,18( 3) :251-261.
