黃海中南部頭足類的群落結構與生物多樣性
2015-04-11 03:25:56李忠義戴芳群陳瑞盛金顯仕
海洋科學 2015年8期
吳 強, 王 俊, 李忠義, 戴芳群, 陳瑞盛, 孫 珊, 金顯仕
(中國水產科學研究院 黃海水產研究所 農業部海洋漁業資源可持續利用重點開放實驗室, 山東省漁業資源與生態環境重點實驗室, 山東 青島 266071)
頭足類具有生命周期短、生長迅速、經濟價值高等特點, 是海洋漁業資源的重要組成部分。中國近海頭足類資源豐富, 是捕撈頭足類的主要國家之一。黃海為半封閉海, 位于中國大陸和朝鮮半島之間,底地形從岸向外傾斜, 在黃海中南部靠近50 m等深線附近存在一由北向南的海槽, 該區域夏季受冷水團控制, 冬季受黃海暖流影響, 是黃海典型生態區[1],具有太平洋褶柔魚(Todarodes pacificus)等頭足類棲息的環境和豐富的餌料基礎, 如太平洋磷蝦(Euphausia pacifica)[2]。目前國內的頭足類群落研究主要限于東海水域, 研究內容包括頭足類的種類組成、數量分布[3-4]及資源量[5-6]等。針對南海頭足類種類組成及資源密度[7-8]的研究也有報道。對黃海頭足類群落的研究則鮮有報道, 已有報道都是以黃海無脊椎動物群落為研究對象, 僅將頭足類作為部分內容進行了簡要分析[9-11]。為了掌握黃海中南部頭足類的資源狀況,本研究根據 2010~2011年間春、秋、冬 3季的底拖網調查資料, 對黃海中南部水域頭足類的種類組成、數量分布及資源量的季節變化做了研究, 旨在摸清黃海中南部水域頭足類的群落結構及數量分布, 了解該水域頭足類的資源狀況, 以期為頭足類資源的管理和開發提供理論依據。
1 材料與方法
1.1 材料來源
數據來自于春季(2010年5月)、秋季(2011年10月)和冬季(2011年 1月)使用“北斗”號科學調查船進行的底拖網調查。調查范圍為黃海中南部(32°00′~37°00′N, 121°00′~125°00′ E), 見圖1。采樣工具為“北斗”號科學調查船專用底拖網, 網口高度為6.1~8.3 m, 網口寬度約25 m, 囊網網目24 mm, 拖速為3.0 kn, 拖網時間為1 h。計劃站位共59個, 受天氣及海況等影響, 實際拖網站位為春季(48個)、秋季(47個)和冬季(36個)。 調查采樣及生物學測定根據《海洋調查規范》[12]進行, 種類鑒定參照董正之[13], 漁獲率統一為單位時間生物量(kg/h)或豐度(個/h)。
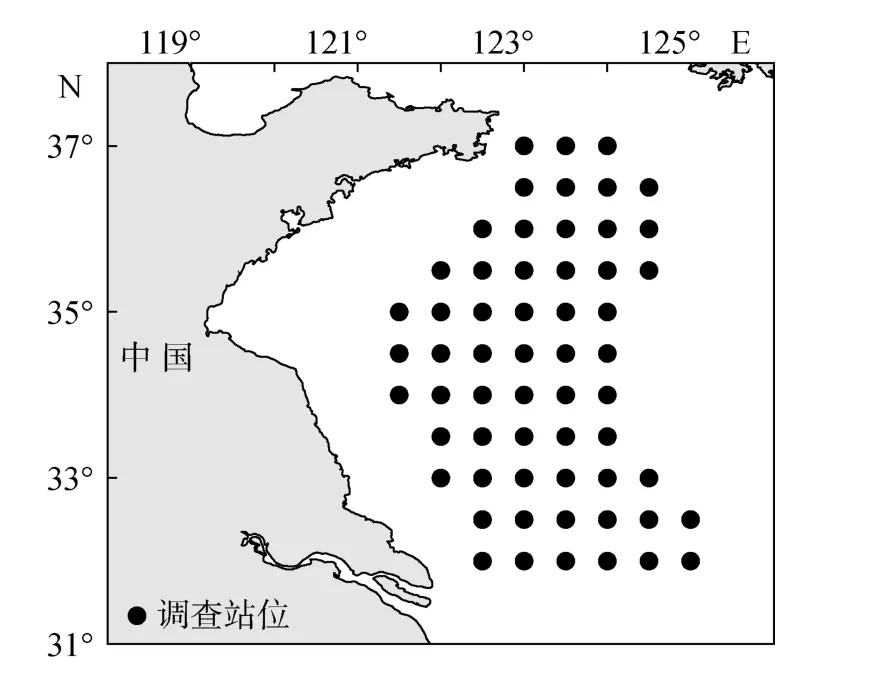
圖1 黃海中南部調查站位Fig.1 Survey stations in central and southern Yellow Sea
1.2 生態優勢度
群落物種生態優勢度的研究采用Pinkas等[14]相對重要性指數(IRI):
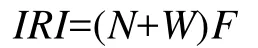
式中,N為某一種類的個體數密度占總個體數的百分率;W為某一種類的生物量占總生物量的百分率;F為某一種類出現的站數占調查總站數的百分率。相對重要性指數(IRI)包含了生物的個體數、生物量和出現頻率 3個重要信息, 常被用來研究漁業生物群落各種類的生態優勢度。物種IRI值大于1000時定為優勢種,IRI值大于等于100且小于1000的種類定義為重要種。
1.3 資源密度
資源密度以拖網掃海面積法來估算, 計算式為:
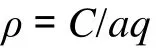
式中:ρ為資源密度(質量: kg/km2);C為平均每小時拖網漁獲量(質量: kg/h);a為網具每小時掃海面積(km2/h), 黃海中南部調查拖網時速為 3 n mile(1n mile=1.852 km), 網口寬度平均為0.0217 km, 因此每網掃海面積為0.12 km2; 為網具捕獲率(q=1–E,E為逃逸率), 本文E取值0.5[3,15-16]。
1.4 生物多樣性
Margalef豐富度指數[17]:D=(S–1)lnN
Shannon-Wiener多樣性指數[18]:

Pielou均勻性指數[19]:J'=H'/lnS
式中:S為種類數;N為生物量或總密度,Pi=ni/N, 為第i種頭足類占總生物量或個體數的比例,ni為第i種頭足類占的生物量或個體數。
1.5 群落結構相似性
利用Bray-Curtis相似性指數[20]計算黃海中南部各季節頭足類類群落結構相似性, 計算公式為:

式中xij,xim分別為第i個種類在j季節和m季節單位時間的資源密度(經過4次方根轉換)。當B=100時,表示兩個群落完全相同; 當B=0時, 表示兩個群落完全不同。
群落結構的差異性利用 ANOSIM (one-way analysis of similarities)程序進行檢驗分析。
1.6 數據處理
數據處理使用EXCEL2010、SPSS 15和PRIMER 5;圖件繪制使用SURFER 8.0和PRIMER 5。
2 結果與分析
2.1 種類組成及優勢種
3個季節于黃海中南部水域共捕獲頭足類13種,隸屬于3目6科6屬, 其中以秋季捕獲種類數最多,為10種, 春季和冬季均捕獲8種。表1統計了各季節各種類的生物量百分比(W)、個體數百分比(N)和出現頻率(F), 進而計算出相對重要性指數(IRI)。可以看出, 春季優勢種為雙喙耳烏賊(Sepiola birostrata)和槍烏賊(Loligospp.), 且雙喙耳烏賊為絕對優勢種;秋季優勢種為槍烏賊和針烏賊; 冬季優勢種為槍烏賊、雙喙耳烏賊和太平洋褶柔魚(Todarodes acificus),其中以槍烏賊相對重要性指數值最高。
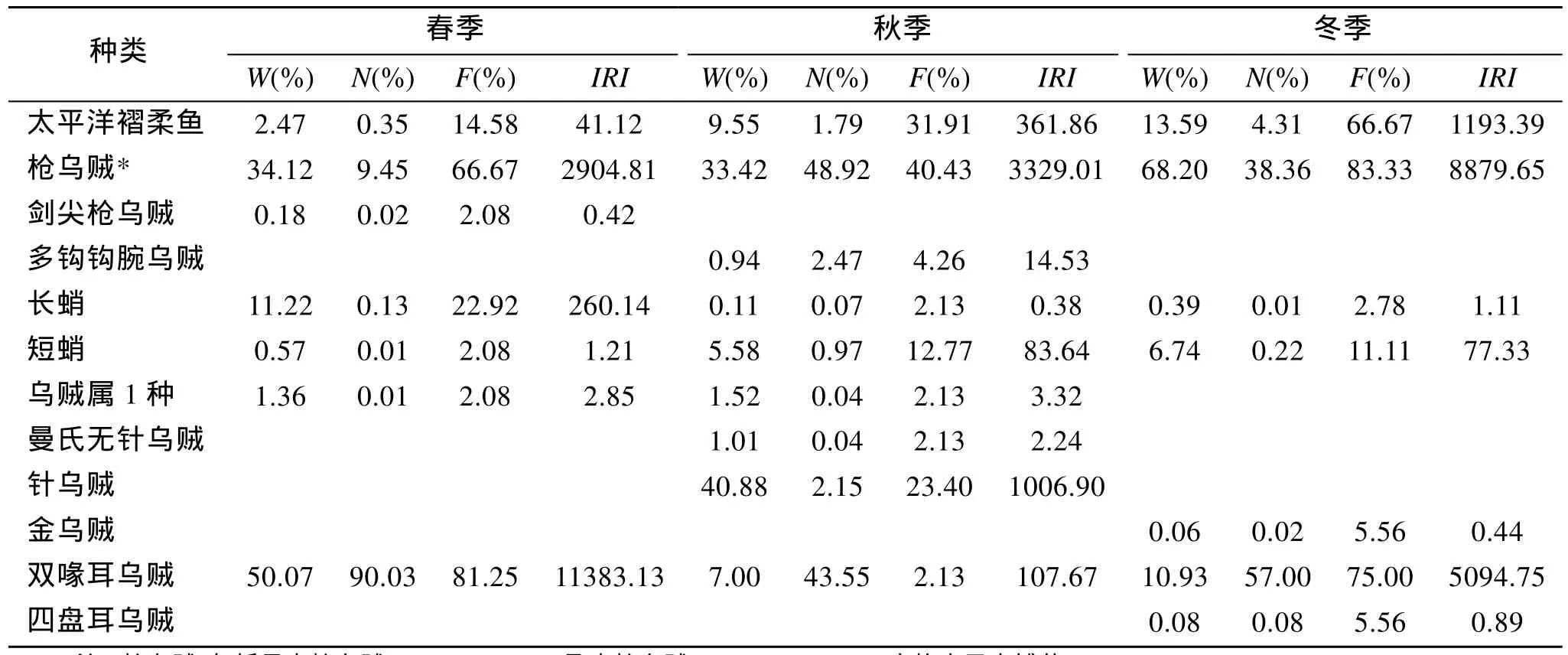
表1 黃海中南部頭足類種類組成及優勢度特征值Tab.1 The species composition and characteristic index of Cephalopoda in Central and Southern Yellow Sea
2.2 資源密度
根據拖網調查結果, 黃海中南部水域春、秋、冬3個季節頭足類平均資源密度為 6.70 kg/km2, 其中,以冬季密度最高, 為 12.72 kg/km2, 其次為春季4.85 kg/km2, 秋季密度最低, 僅為2.52 kg/km2。綜合春、秋、冬3個季節, 槍烏賊和雙喙耳烏賊在黃海中南部水域資源密度最高(表2)。
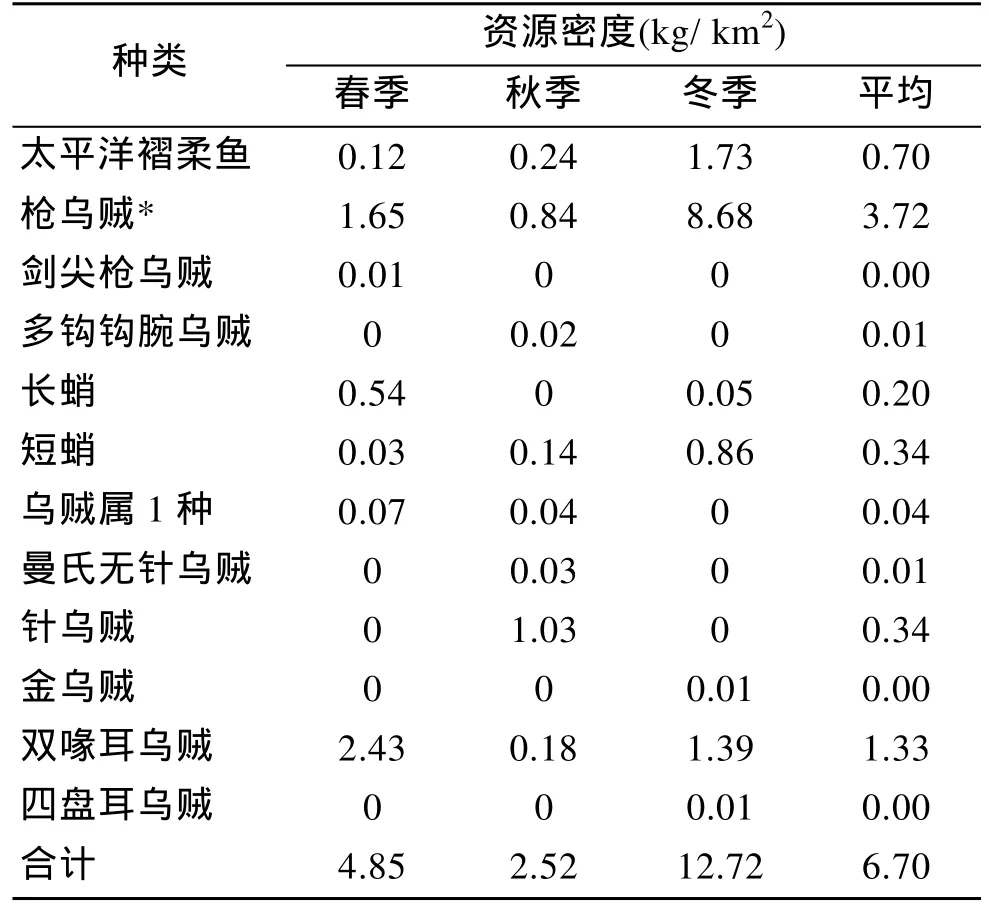
表2 黃海中南部頭足類資源密度Tab.2 The resources density of Cephalopoda in central and southern Yellow Sea
捕獲頭足類可分為 3大類, 分別為槍形目(Teuthoidea)(即魷魚類, 包括太平洋褶柔魚、火槍烏賊、日本槍烏賊、劍尖槍烏賊(Loligo edulis)和多鉤鉤腕烏賊(Abralia ltihamata))、八腕目(Octopoda)(即章魚類, 包括長蛸(Octopus variabilis)和短蛸(Octopus ocellatus))和烏賊目(Sepioidea)(即烏賊類, 包括烏賊屬1種、曼氏無針烏賊(Sepiella maindroni)、針烏賊(Sepia esculenta)、金烏賊(Sepia esculenta)、雙喙耳烏賊(Sepiola birostrata)和四盤耳烏賊(Euprymna morsei))。黃海中南部水域頭足類生物量組成中, 春季以烏賊目和槍形目為主, 分別占總生物量的51.43%和 36.78%; 秋季以烏賊目和槍形目為主, 分別占 51.35%和 42.95%; 冬季以槍形目占絕對優勢,占 81.79%; 綜合 3個季節, 槍形目占頭足類生物量的66.17%, 八腕目占8.07%, 烏賊目占25.75%(圖2)。
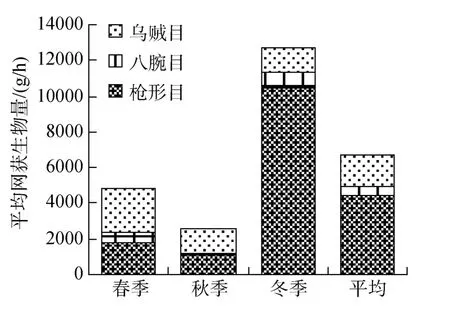
圖2 黃海中南部頭足類生物量組成Fig.2 The biomass composition of Cephalopoda in central and southern Yellow Sea
黃海中南部水域頭足類個體數組成中, 春季以烏賊目占絕對優勢, 占總個體數的 90.04%; 秋季以槍形目和烏賊目為主, 分別占 50.71%和 45.78%; 冬季以烏賊目和槍形目為主, 分別占 57.01%和 42.67%;綜合 3個季節, 槍形目占頭足類個體數的 27.66%,八腕目占0.26%, 烏賊目占72.08%(圖3)。
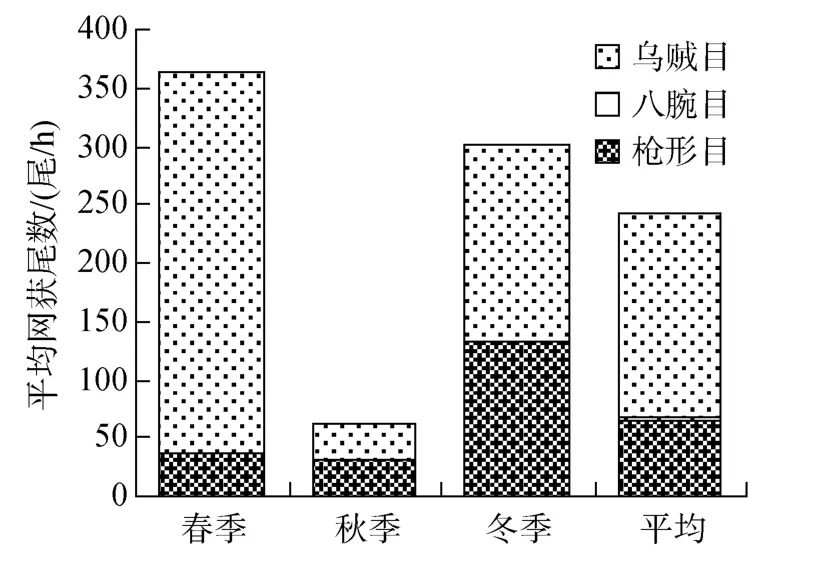
圖3 黃海中南部頭足類個體數組成Fig.3 The individual composition of Cephalopoda in central and southern Yellow Sea
2.3 空間分布
2.3.1 水平分布
由圖4可以看出黃海中南部水域不同季節頭足類的生物量分布狀況。春季, 頭足類主要分布在石東漁場與連東漁場交界水域和海州灣漁場東南部, 主要種類為雙喙耳烏賊、槍烏賊及長蛸, 其中雙喙耳烏賊比例最高, 占總生物量的 50.07%; 秋季, 頭足類以連青石漁場西南部及大沙漁場北部密度最高, 主要以針烏賊及槍烏賊為主, 兩者分別占頭足類總生物量的 40.88%和 33.42%; 冬季, 頭足類數量以石島漁場東南部以及連青石漁場密度最高,主要以槍烏賊為主, 占頭足類總生物量的68.20%。綜合來看, 黃海中南部頭足類生物量分布的季節變化較大。
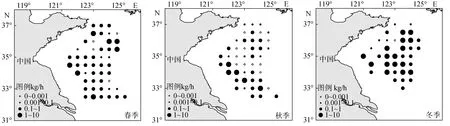
圖4 黃海中南部頭足類生物量分布Fig.4 The biomass distribution of Cephalopoda in central and southern Yellow Sea
2.3.2 垂直分布
鑒于頭足類不同季節棲息水深的變化, 研究了黃海中南部各季節頭足類在各水深的生物量分布,考慮到太平洋褶柔魚在生物量組成中的優勢地位,將其單獨列出進行統計分析。由表3可看出: 春季,頭足類生物量的 47.0%分布在 60 m 以深水域; 其中,槍形目主要分布在60 m以深水域, 八腕目主要分布在20~40 m水深, 烏賊目主要分布在20~40 m水深。秋季,頭足類生物量的 50.4%分布在 40~60 m 水深; 其中,93.1%的太平洋褶柔魚分布在60 m以深水域, 其他槍形目主要分布在 40~60 m水深, 八腕目主要分布在16~40m水深, 烏賊目主要分布在40~60 m水深。冬季,65.1%的頭足類分布在60 m以深; 其中, 太平洋褶柔魚及其他槍形目絕大多數分布在60 m以深, 八腕目主要分布在15~40 m水深, 烏賊目主要分布在60 m以深。總體來看, 黃海中南部, 冬、春 2季頭足類主要分布在60 m以深水域, 秋季頭足類主要分布在40~60 m水深。
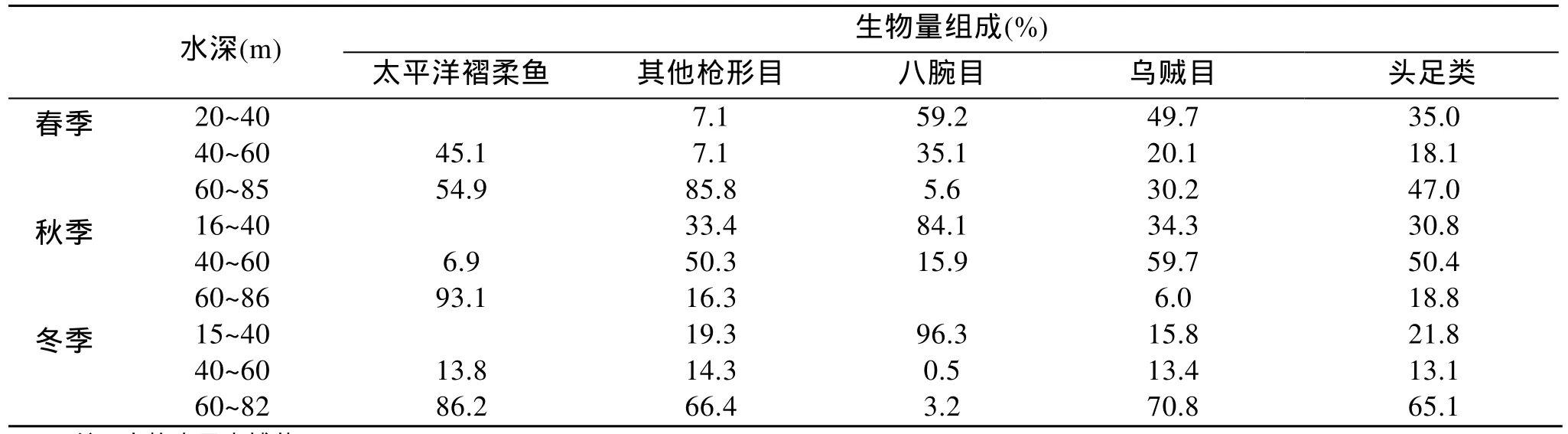
表3 頭足類在不同水深的生物量組成Tab.3 The biomass composition of the Cephalopoda in different depth of water
2.4 生物多樣性指數
黃海中南部頭足類各季節生物多樣性指數及標準差見表4。根據生物量, 黃海中南部頭足類豐富度指數、均勻度指數及多樣性指數均以冬季最高, 其次是春季, 秋季各項指數最低。根據個體數, 黃海中南部頭足類豐富度指數、均勻度指數及多樣性指數的季節變化趨勢與根據生物量得出的結果一致, 冬季最高, 其次是春季, 秋季各項指數最低。利用One-Way ANOVA 程序中的 Tukey 檢驗, 分別根據生物量與個體數數據, 統計分析了黃海中南部頭足類春、秋、冬季之間的變異程度。根據生物量, 春季與秋季、秋季與冬季的Margalef豐富度指數(D),以及春季與冬季的 Shannon-Wiener多樣性指數(H')均呈顯著性差異(P<0.05); 春季與秋季、秋季與冬季的Pielou均勻性指數(J')均呈極顯著差異(P<0.01)。根據個體數, 春季與冬季的 Margalef豐富度指數呈顯著性差異(P<0.05); 秋季與冬季的Pielou均勻性指數、春季與冬季以及秋季和冬季的Shannon-Wiener多樣性指數均呈極顯著差異(P<0.01)。
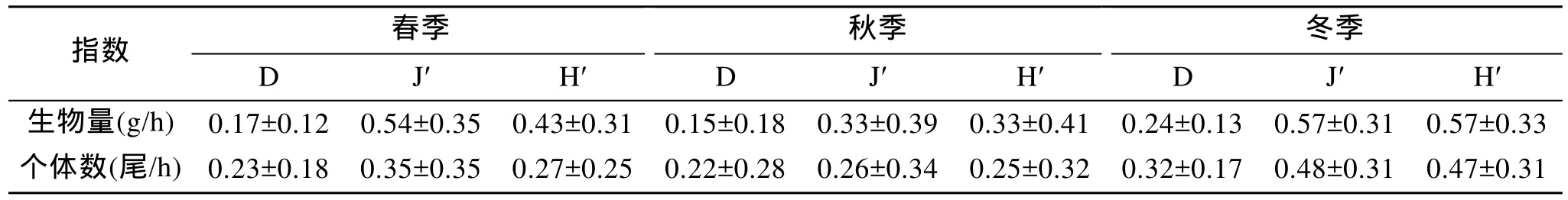
表4 黃海中南部頭足類生物多樣性指數Tab.4 Diversity index of Cephalopoda in central and southern Yellow Sea
2.5 群落結構季節變化
為了便于數據對比, 根據春、秋、冬3個季節共同站位的生物量及個體數資料, 利用 PRIMER軟件中的Similarity程序分析了黃海中南部頭足類的群落結構相似性, 并利用 ANOSIM程序對各季節進行相似性分析檢驗, 結果見表6。根據生物量, 以春季和冬季頭足類群落結構相似性指數最高(66.77), 其次是春季和秋季(62.73), 秋季和冬季相似性指數最低(55.30); 各季節兩兩間差異性均顯著(P<0.05)。 根據個體數, 同樣以春、冬2季黃海中南部頭足類的群落結構相似性指數最高(74.87), 其次是春季和秋季(69.83), 秋季和冬季相似性指數最低(65.15); 各季節兩兩間差異性均顯著(P<0.05)。
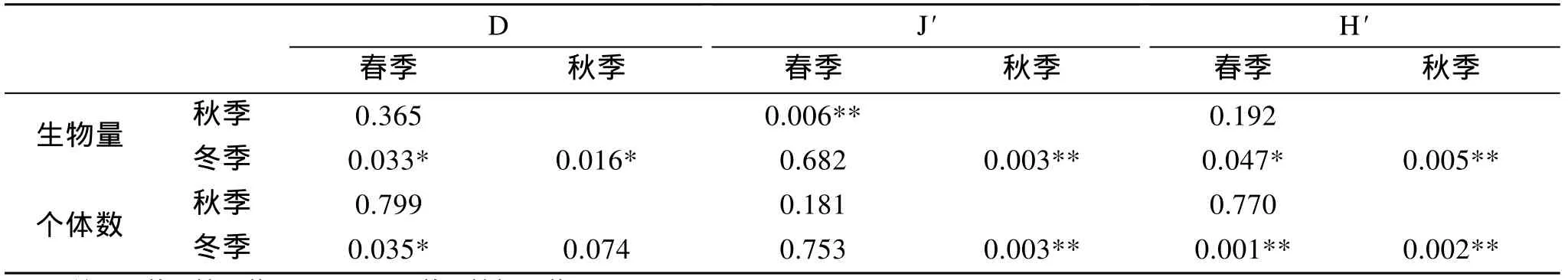
表5 黃海中南部頭足類多樣性指數的季節差異顯著性Tab.5 The significance level (p) of seasonal variation in diversity index of Cephalopoda in central and southern Yellow Sea
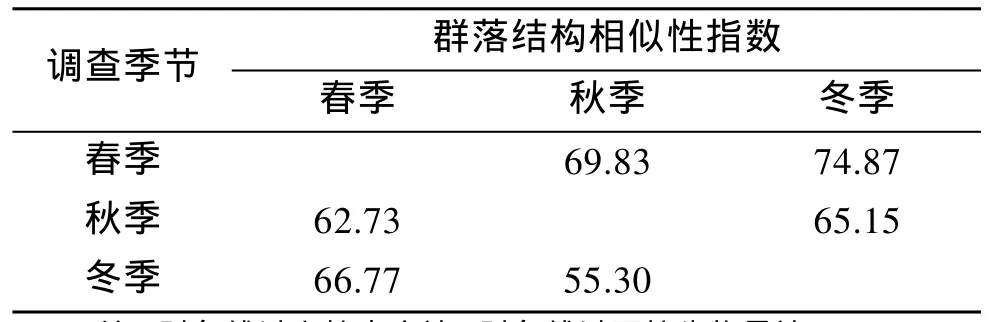
表6 黃海中南部頭足類群落結構相似性指數Tab.6 Similarity index of Cephalopoda community between different seasons in central and southern Yellow Sea
3 討論
3.1 與歷史調查結果的比較
金顯仕等[16]曾于 1998~2000年采用完全相同的方法對黃海中南部頭足類資源進行了調查。表7對比了1998~2000年與2010~2011年春、秋、冬3個季節的調查結果, 與1998~2000年相比, 本次調查采集頭足類種類數增加 5種, 資源密度增長了 12%。1998~2000年優勢種類為槍烏賊及太平洋褶柔魚,分別占頭足類總生物量的 44.20%和 41.50%,2010~2011年槍烏賊的比重上升至 55.61%, 太平洋褶柔魚的比重則下降至 10.40%, 耳烏賊的比重大幅上升至19.93%。
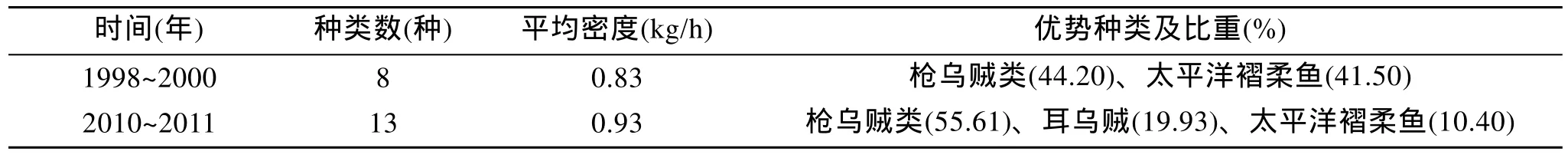
表7 2010~2011年與1998~2000年調查結果的對比Tab.7 Comparison of the survey results between 2010-2011 and 1998-2000
對比兩次調查頭足類生物量的分布狀況: 春季,1998年5月以大沙漁場東北部密度較高, 而2010年5月以石東漁場與連東漁場交界水域和海州灣漁場東南部密度較高; 秋季, 2000年10月頭足類主要集中在石東漁場、石島漁場與連青石漁場的交界水域,2011年10月則以呂泗漁場及大沙漁場密度較高; 冬季, 1999年12月以石東漁場密度較高, 2011年1月則以石島漁場東南部及連青石漁場密度較高。總體來看, 與1998~2000年同期對比, 本次調查黃海中南部春、秋、冬 3個季節頭足類生物量的分布狀況變化較大。
3.2 與其他海區的比較
本次調查, 2010年春季(5月)于黃海中南部共采集頭足類8種, 其中以雙喙耳烏賊(50.10%)和槍烏賊(34.02%)占絕大部分比重。吳強等[11]曾于同期(2010年 5月)對渤海和黃海北部水域進行調查, 于渤海、黃海北部捕獲頭足類均為5種; 其中, 渤海頭足類以槍烏賊(47.97%)和短蛸(28.94%)占大部分生物量比重, 黃海北部頭足類則以短蛸(73.16%)和槍烏賊(13.42%)占大部分比重。
本次調查于黃海中南部共采集頭足類13種, 春季、秋季均以烏賊類比重最高, 冬季則以魷魚占絕大部分比重(圖2)。宋海棠等[21-22]于1994年6月至1996年 5月曾逐月對東海北部的頭足類做了調查研究,共采集頭足類33種, 春(5月)、秋(10月)、冬(1月)3個季節均以烏賊類占大部分比重。綜合看來, 黃海中南部頭足類種類數明顯少于東海北部; 黃海中南部以魷魚和烏賊類主要捕獲對象, 而東海北部則以烏賊類為主要捕獲對象。
3.3 國內外頭足類群落結構的研究方法比較
相對于國內較普遍的基于拖網調查的頭足類群落結構研究[23-25], 國外常采用大型捕食者胃含物分析的方法來研究頭足類的種類組成及生物多樣性[26-28]。例如: Staudinger[26]等研究了新英格蘭近海、中大西洋灣及南大西洋灣頂級捕食者胃含物中的頭足類種類組成及多樣性, 發現南大西洋灣頭足類多樣性指數最高, 其次是新英格蘭近海, 中大西洋灣最低;González[27]等利用擱淺哺乳動物胃含物中角質喙,確定了西班牙西北近海的頭足類隸屬9科、12種, 尾數組成中以槍烏賊科(Loliginidae)(56.9%)和章魚科(Octopodidae)(25.3%)為主, 柔魚科(Ommastrephinae)(11.9%)和耳烏賊科(Sepiolidae)(2.4%)的比例則較小;Velasco[28]等則通過胃含物分析研究了頭足類作為被捕食者對底層魚類的重要作用。綜合對比, 作者認為基于拖網調查的頭足類群落結構分析方法更直接,有利于如實地反映自然海域中頭足類的群落結構;而胃含物分析法則更有利于認識頭足類與大型捕食者攝食關系, 有利于認識頭足類在生態系統能量流動中的重要作用。
[1] 蘇紀蘭, 袁業立.中國近海水文[M].北京: 海洋出版社, 2005: 193-206.
[2] 董正之.黃海太平洋褶柔魚資源現狀與開發[J].海洋科學, 1996, 20(6): 34-38.
[3] 宋海棠, 丁天明, 余匡軍, 等.東海北部頭足類的種類組成和數量分布[J].浙江海洋學院學報(自然科學版), 1999, 18(2): 99-106.
[4] 覃濤, 俞存根, 陳全震, 等.舟山漁場及鄰近海域頭足類種類組成和數量分布[J].海洋與湖沼, 2011, 42(1):124-130.
[5] 丁天明, 宋海棠.東海中北部海區頭足類資源量的評估[J].水產學報, 2011, 25(3): 215-221.
[6] 凌建忠, 嚴利平, 李建生.應用資源密度面積法估算東海區頭足類資源量[J].海洋漁業, 2008, 30(1): 56-61.
[7] 董正之, 呂榮書, 李永明.南海深海區頭足類調查簡報[J].海洋科學, 1982, 6(2): 45.
[8] 黃梓榮.南海北部陸架區頭足類的種類組成和資源密度分布[J].南方水產, 2008, 4(5): 1-7.
[9] 吳耀泉.萊州灣主要無脊椎動物資源及其群聚多樣性特征[J].海洋與湖沼, 1995, 26(6): 606-609.
[10] 程濟生.黃海無脊椎動物資源結構及多樣性[J].中國水產科學, 2005, 12(1): 68-75.
[11] 吳強, 王俊, 金顯仕, 等.中國北部海域主要無脊椎動物群落結構及多樣性[J].中國水產科學, 2011, 18(5):1152-1160.
[12] 國家技術監督局.GB/T 12763.2-1991.海洋調查規范—海洋生物調查[S].北京: 中國標準出版社, 1991.
[13] 董正之.中國動物志(軟體動物門頭足綱)[M].北京:科學出版社, 1988: 81-84.
[14] Pinkas L, Oliphamt M S, Iverson I L K.Food habits of albacore, bluefin tuna, and bonito in California waters [J].Calif Dep Fish Game Fish Bull, 1971, 152: 1-105.
[15] Aglen A, Foyn L, Godos O R, et al.Surveys of the marine fish resources of Peninsular Malaysia, June-July,1980[A].Bergen: Institue of Marine Research, 198l: 9:l-69.
[16] 金顯仕, 趙憲勇, 孟田湘, 等.黃渤海生物資源與棲息環境[M].北京, 科學出版社, 2005: 369-379.
[17] Margalef R.Information theory in ecology [J].General System, 1958, 3: 36-71.
[18] Wilhm J L.Use of biomass units in Shannon's formula [J].Ecology, 1968, 49: 153-156.
[19] Pielou E C.Ecological Diversity [M].New York: Wiley,1975: 35-86.
[20] Simpson E H.Measurement of diversity [J].Nature,1949, 163: 688.
[21] 宋海棠, 余匡軍.太平洋褶柔魚在東海的分布和洄游[J].海洋水產科技, 1999, 1: 9-14.
[22] 宋海棠, 丁天明.東海北部頭足類的種群組成和數量分布[J].浙江海洋學院學報(自然科學版), 1999,18(2): 99-105.
[23] 李成林, 張艷萍, 趙斌, 等.膠州灣曼氏無針烏賊資源量與季節變化的調查研究[J].漁業科學進展, 2009,30(3): 62-67.
[24] 俞存根, 虞聰達, 寧平, 等.浙江南部外海頭足類種類組成和數量分布[J].海洋漁業, 2009, 31(1): 27-33.
[25] 陳強, 王家樵, 張雅芝, 等.福建閩江口及附近海域和廈門海域頭足類種類組成的季節變化[J].海洋學報,2012, 34(3): 179-184.
[26] Staudinger M D, Juanes F, Salmon B, et al.The distribution, diversity, and importance of cephalopods in top predator diets from offshore habitats of the Northwest Atlantic Ocean [J].Deep Sea Research Part II:Topical Studies in Oceanography, 2013, 95: 182-192.
[27] González A F, López A, Guerra A, et al.Diets of marine mammals stranded on the northwestern Spanish Atlantic coast with special reference to Cephalopoda[J].Fisheries Research, 1994, 21(1-2): 179-191.
[28] Velasco F, Olaso I, Sánchez F.The role of cephalopods as forage for the demersal fish community in the southern Bay of Biscay[J].Fisheries Research, 2001,52(1-2): 65-77.
