東北豆科山羊豆族4屬植物葉形態特征及數量分類學研究
2015-04-15 08:04:20史傳奇劉玫王臣
草業學報 2015年5期
關鍵詞:植物
史傳奇,劉玫,王臣
(哈爾濱師范大學生命科學與技術學院,黑龍江省分子細胞遺傳與遺傳育種重點實驗室,黑龍江 哈爾濱 150025)
?
東北豆科山羊豆族4屬植物葉形態特征及數量分類學研究
史傳奇,劉玫*,王臣
(哈爾濱師范大學生命科學與技術學院,黑龍江省分子細胞遺傳與遺傳育種重點實驗室,黑龍江 哈爾濱 150025)
本研究采用GMA切片、掃描電鏡及葉片透明法,深入研究了東北豆科山羊豆族4屬25種植物葉的形態結構。選取與葉相關的23個形態性狀,參考8個非葉的特征,進行了聚類分析與主成分分析。結果表明,相似系數在0.58處時,4屬植物被分為3組,黃耆屬與米口袋屬為一組,棘豆屬和甘草屬各自為一組。小葉是否具腺體、葉柄橫切面形狀及葉柄細胞是否具單寧等特征具有較高的絕對權重值,可以作為區分刺果甘草與甘草和其他山羊豆族植物的依據。托葉質地、托葉與葉柄聯合程度及小葉葉緣是否反卷等特征可作為棘豆屬的識別特征。托葉形狀在屬下種間具一定的分類學價值。本研究為山羊豆族植物分類學及豆科植物系統學研究提供了葉形態學依據。
山羊豆族;葉;形態;結構;分類學
葉的形態結構(如氣孔類型、表皮細胞垂周壁式樣、葉柄橫切面形狀等)對分類學具有重要的價值[1-8]。Mehrabian等[9]認為伊朗黃耆屬(Astragalus)Incani組葉柄維管束數目、髓部薄壁細胞層數等特征可以作為種的分類依據,但對組間分類的意義不大。康云和李明理[10]指出蔟毛黃耆亞屬(subgenusPogonophace)葉表皮細胞形狀與垂周壁式樣可作為屬下組間的分類依據,角質層紋飾對種的鑒定具有一定的意義。甘草屬(Glycyrrhiza)植物葉表面氣孔有不等型和無規則型,可作為研究甘草屬植物分類和演化的依據[11]。陸嘉惠等[12]研究甘草屬葉表皮特征,提出葉表皮細胞垂周壁式樣,氣孔類型及毛狀體類型可作為劃分甘草屬內種以上等級的依據。劉麗和趙一之[13]根據托葉與葉柄分離或合生、小葉著生方式及小葉數目等形態特征,利用分支分類的方法,探討了內蒙古棘豆屬(Oxytropis)27個物種的系統發育關系。
形態學及分子系統學研究揭示山羊豆族(tribe Galegeae)是一個復系類群,分類學一直存在諸多爭議[14],本研究的目的是通過深入研究東北山羊豆族4屬植物葉的形態結構,選取23個質量性狀,并結合前人研究工作,通過聚類分析與主成分分析,為該族植物的分類及系統學研究提供形態學依據。
1 材料與方法
1.1 材料
研究材料為山羊豆族黃耆屬13種,甘草屬2種,米口袋屬2種及棘豆屬8種。新鮮材料于2014年5-9月采自于中國東北部,經哈爾濱師范大學王臣教授鑒定。憑證標本存放于哈爾濱師范大學植物標本館(Harbin Normal University, HANU),干材料于2014年9月取自哈爾濱師范大學植物標本館和東北生物標本館(Institute of Forestry and Pedology, IFP)。詳細信息見表1。
1.2 方法
葉片透明:按照任海燕等[15]的方法,每一物種取兩個植株的成熟葉片,將其置于5%NaOH水溶液中,24 h后取出,50%甘油水溶液封片,Olympus BX53顯微鏡觀察葉表皮細胞特征,重復觀察3次,用Olympus DP26數碼相機照相。參照Dilcher[16]系統鑒定氣孔類型。
葉片掃描:根據陳曦等[17]的方法,切取正方形(4 mm2)成熟葉片,用雙面膠將樣品粘在樣品臺上,Eiko IB-3離子濺射儀導電處理,濺射鍍膜厚度為7~8 nm,在Hitachi S-4800型掃描電鏡下(加速電壓為5 kV)觀察毛狀體形態并照相。
GMA(glycol methacrylate,乙二醇甲基丙烯酸酯)切片[18]:新鮮材料用FAA(Formalin-Acetic acid-70% Alcohol mixture,福爾馬林∶冰醋酸∶70%酒精=5∶5∶90)固定,時間不少于24 h。干材料放入熱水中約30 min,材料充分吸水膨脹后,用FAA固定24 h以上。每個物種各取3份葉柄的中部,經50%酒精、100%酒精、異丙醇、正丁醇脫水處理各2次,每次6 h。進入GMA滲透3次,前2次各為1 d,第3次5~7 d,之后將材料及GMA裝入膠囊,置于60℃溫箱聚合24 h。用Leica Ultralcut R切片,厚度2.0~2.5 μm。將切片置于DNPH(2, 4-dinitrophenylhydrazine,2, 4-二硝基苯肼)冰醋酸水溶液中30 min,流水沖洗10 min。置于高碘酸水溶液中10 min,流水沖洗5 min,將切片烘干后,置于Schiff試劑30 min,流水沖洗1~2 min,切片烘干。最后置于甲苯胺藍溶液中染色1~2 min,流水沖洗10 min,烘干后用加拿大樹膠封片。切片同樣經Olympus BX53顯微鏡觀察,Olympus DP26數碼相機拍照。
根據《中國植物志》[19-20]記載的8個非葉形態特征及本研究所觀察的23個葉的形態特征,參照徐克學[21]的方法,對31個質量性狀編碼:01生長習性:多年生草本0/一或二年生草本1;02花序:葉叢基部葉腋處著生0/總狀1/傘形2;03花萼形狀:管狀鐘形0/鐘形1/管狀2/筒狀3;04花藥藥室頂部:不聯合0/聯合1;05花柱柱頭蔟毛:無0/有1;06龍骨瓣具喙:否0/是1;07果皮質地:革質0/膜質1;08果皮附屬物:無0/有1;09葉著生方式:基生或近基生0/莖生1;10葉型:輪生羽狀復葉0/奇數羽狀復葉1;11托葉質地:草質0/膜質1;12托葉形狀:近卵形0/三角形1/披針形2;13托葉先端形態:漸尖0/銳尖1;14托葉與葉柄聯合情況:離生或僅基部聯合0/中部附近聯合1/大部分聯合2;15小葉基部:近圓形0/圓形或楔形1/楔形2;16小葉先端:多鈍0/鈍或尖1/多尖2;17小葉葉緣:全緣0/微齒1;18小葉葉緣反卷:否0/是1;19小葉多細胞腺體:無0/有1;20小葉表皮毛狀體分布:無0/下表面1/兩面2;21小葉邊緣毛狀體:無0/有1;22小葉非腺毛類型:單毛0/丁狀毛1;23小葉氣孔分布:下表面0/兩面1;24小葉氣孔類型:只具無規則型0/兼具不等型及無規則型1;25小葉上表皮垂周壁式樣:弓形0/弓形-波形1/波形2;26小葉下表皮垂周壁式樣:弓形0/弓形-波形1/波形2;27葉柄橫切面形狀:近圓形0/心形1/戟形2;28葉柄近軸面凹槽:無0/有1;29葉柄維管束排列:不連續0/近連續1;30葉柄橫切面皮層細胞形狀:近圓形0/柱形1;31葉柄細胞內含單寧:否0/是1。
參考Zhou等[22]的方法,應用NTSYS pc 2.10e軟件包,采用UPGMA法(unweighted pair group method using arithmetic average,非加權配對算術平均法)構建聚類樹,Mantel’s test[23]檢驗聚類樹與相似系數矩陣的一致性。參照李鴻雁等[24]的方法,應用SPSS 17.0做主成分分析,求出各主成分的特征值、貢獻率、累計貢獻率及前3個主成分中各性狀特征的權重值,并基于前2個主成分繪制二維散點圖。
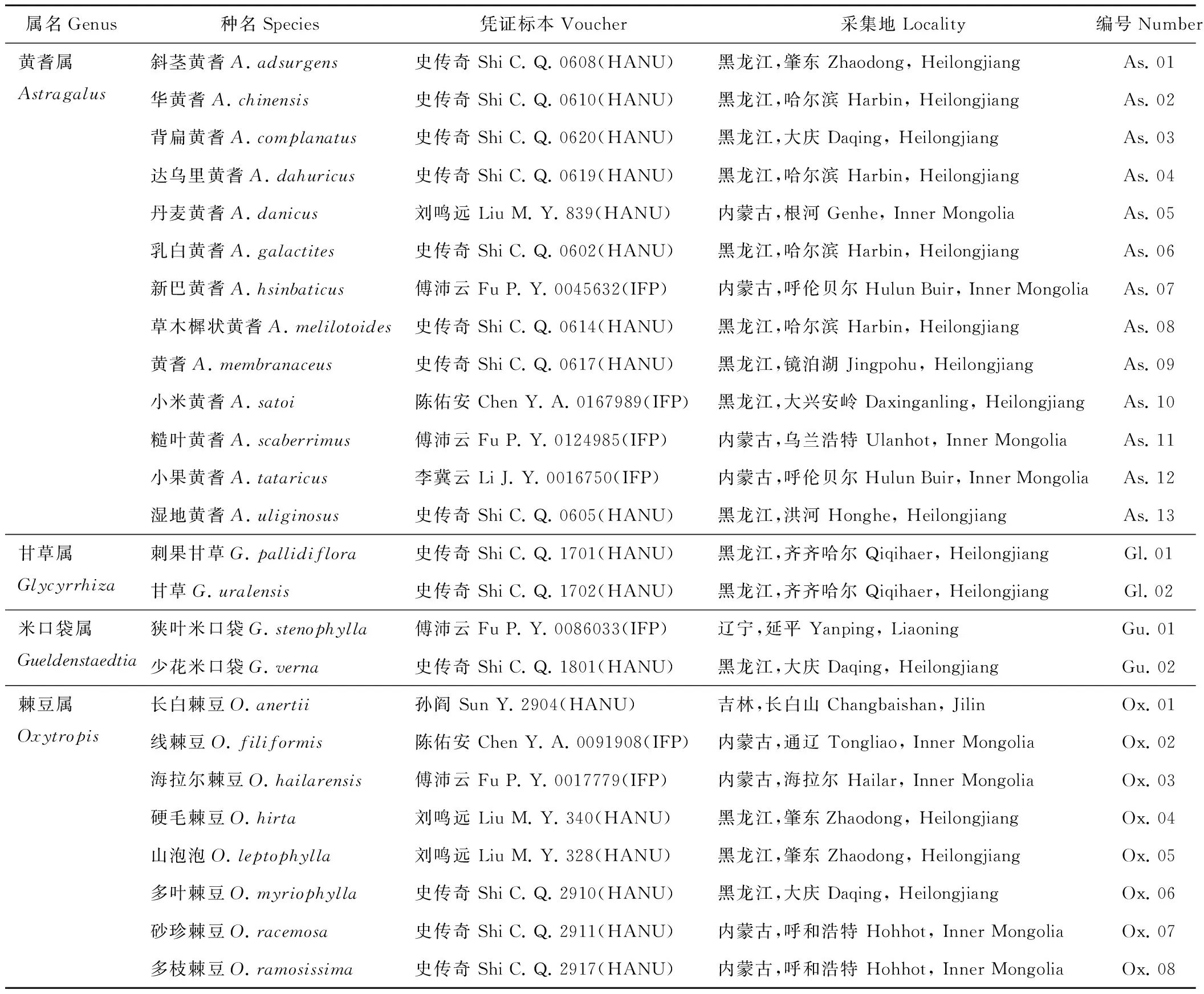
表1 東北山羊豆族4屬植物(用于葉特征研究)基本信息[19-20]
2 結果與分析
2.1 葉片形態及葉柄結構
葉外部形態:山羊豆族葉著生方式分為基生或近基生和莖生,黃耆屬多為莖生,如斜莖黃耆,而乳白黃耆和新巴黃耆的葉為基生或近基生。葉型多為奇數羽狀復葉,也有輪生羽狀復葉,如棘豆屬的海拉爾棘豆。托葉草質或膜質,棘豆屬多為膜質,多枝棘豆的為草質,其他3屬多為草質,而乳白黃耆為膜質。托葉形狀劃分為近卵形、三角形和披針形,在種間有差異,如米口袋屬的狹葉米口袋的為三角形,少花米口袋的為披針形。托葉先端漸尖或銳尖,黃耆屬的達烏里黃耆、小米黃耆和小果黃耆的為銳尖,其余物種的皆為漸尖。托葉和葉柄分離或僅在基部聯合,大部分物種屬于此類,黃耆屬的糙葉黃耆、棘豆屬的硬毛棘豆等聯合至托葉中部,棘豆屬的山泡泡和砂珍棘豆的幾乎全部聯合。小葉基部近圓形或楔形,也有同一植株上不同葉片的小葉基部兼具圓形和楔形的,如狹葉米口袋。甘草屬和棘豆屬小葉先端多尖,米口袋屬的小葉先端形態多樣,鈍或尖,黃耆屬的多為鈍圓,或有時具凹陷,如黃耆。除甘草屬的刺果甘草葉緣具微齒外,其余物種葉緣均為全緣。棘豆屬小葉葉緣反卷(硬毛棘豆除外),為本屬特有。
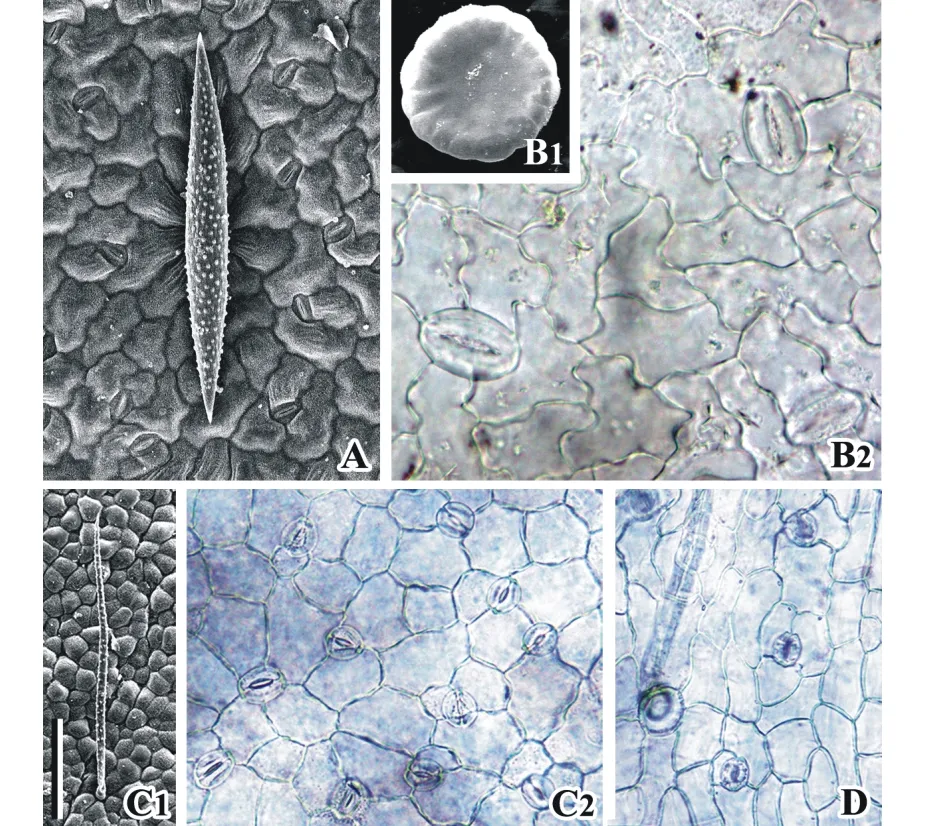
圖1 山羊豆族植物葉表皮毛狀體、垂周壁及氣孔形態Fig.1 Trichome, anticlinal wall and stomata morphology of leaf epidermis of tribe GalegeaeA: 斜莖黃耆葉上表皮 Upper epidermis of A. adsurgens leaf; B1: 甘草葉上表皮腺體 Gland on the upper epidermis of G. uralensis leaf; B2: 甘草葉上表皮Upper epidermis of G. uralensis leaf; C1: 少花米口袋葉下表皮單毛 Single-celled hair on lower epidermis of G. verna leaf; C2: 少花米口袋葉下表皮 Lower epidermis of G. verna leaf; D: 砂珍棘豆葉上表皮 Upper epidermis of O. racemosa leaf. 標尺 Scale bar: 100 μm in A; 50 μm in B1, C2, D; 25 μm in B2; 200 μm in C1.

圖2 山羊豆族植物葉柄橫切面結構Fig.2 Petiole structure in transversal section of tribe GalegeaeA: 斜莖黃耆 A. adsurgens; B: 草木樨狀黃耆A. melilotoides; C: 刺果甘草G. pallidiflora; D: 少花米口袋G. verna; E: 砂珍棘豆O. racemosa; F: 多葉棘豆O. myriophylla; G: 多枝棘豆O. ramosissima. c: 柱狀皮層細胞 cylindrical cortex cell; e: 表皮 epidermis; h: 毛狀體 hair; t: 單寧 tannin; v: 維管束 vascular bundle. 標尺 Scale bar: 300 μm in A, C; 200 μm in B, G; 500 μm in D; 450 μm in E, F.
葉表皮特征:葉表面具多細胞腺體、丁狀毛和單毛。刺果甘草和甘草小葉上、下表面均具有多細胞的腺體(圖1B1)。黃耆屬的部分種具丁狀毛,如斜莖黃耆(圖1A)等,多數物種具單毛,如少花米口袋(圖1C1)。毛狀體通常分布于小葉上、下表面及葉緣。小葉上、下表面均具有氣孔分布,僅黃耆屬的黃耆小葉的上表面未見氣孔。氣孔主要為無規則型和不等型,多數物種兼具兩種類型(圖1A,C2,D),刺果甘草和甘草(圖1B2)的氣孔僅具無規則型。葉表皮垂周壁呈弓形、兼具弓形與波形和波形,如少花米口袋(圖1C2)下表皮細胞垂周壁為弓形,斜莖黃耆(圖1A)的為弓形-波形,甘草(圖1B2)的為波形。
葉柄結構:葉柄橫切面為心形、近圓形或戟形,黃耆屬和米口袋屬的皆為心形,如斜莖黃耆(圖2A)、草木樨狀黃耆(圖2B)和少花米口袋(圖2D),刺果甘草的為戟形(圖2C),棘豆屬具心形或近圓形,前者如砂珍棘豆(圖2E),后者如多枝棘豆(圖2G)。多數物種葉柄近軸面具凹槽(圖2A~E)。葉柄具毛狀體,表皮細胞1層,皮層為4~6層薄壁細胞,呈柱狀(其細胞長軸同葉柄橫切面平行)或圓形,前者多見于草木樨狀黃耆(圖2B)和多枝棘豆(圖2G)。黃耆屬、甘草屬和米口袋屬葉柄具3個明顯較大的維管束,分布不連續,近連續分布的只出現在棘豆屬部分物種,如多葉棘豆(圖2F)。單寧僅存在于刺果甘草和甘草葉柄細胞內(圖2C),其余3屬均未見。詳細特征見表2。

表2 山羊豆族植物31個形態學質量性狀編碼
2.2 聚類分析
聚類樹與相似系數矩陣的一致性較好(r=0.82,t=11.57)。聚類結果如圖3,在SM(simple matching,簡單匹配)系數為0.56時,棘豆屬最先被分出,成為獨立的一個類群。在0.58時,甘草屬被分出,而黃耆屬和米口袋屬仍聚在一起。至0.83處,米口袋屬才從黃耆屬區分出來。聚類結果可以明顯看出,本研究選取的31個形態學性狀能較好地區分黃耆屬、甘草屬及棘豆屬,但狹葉米口袋與少花米口袋和黃耆屬的物種區分不十分明顯,并且對屬下個別種的區分效果也不十分顯著,如黃耆屬的華黃耆和丹麥黃耆、棘豆屬的海拉爾棘豆和砂珍棘豆,說明所選取的31個質量性狀在兩者之間有較高的相似性。
2.3 主成分分析
主成分分析結果(表3)可以看出,前3個主成分的累計貢獻率為51.36%,至第6個主成分累計貢獻率達到70%以上。從31個質量性狀中篩選出與葉相關的形態特征,第1主成分中小葉是否具腺體(0.973)、小葉氣孔類型(-0.973)及葉柄細胞是否含單寧(0.973)具有相同的絕對權重值,說明對4屬植物具有同等的分類價值,小葉葉緣形態的權重值為0.788,這些性狀都可作為區分刺果甘草與甘草和其他3屬植物的重要依據。第2主成分中托葉質地(0.818)、托葉與葉柄聯合程度(0.716)及小葉葉緣是否反卷(0.755)的絕對權重值大于0.7,可以作為棘豆屬植物的識別特征。第3主成分中葉柄形狀(0.728)、近軸面是否具凹槽(0.759)及維管束分布(0.675)具有較高的絕對權重值,可作為屬間及棘豆屬屬下種間的分類依據。
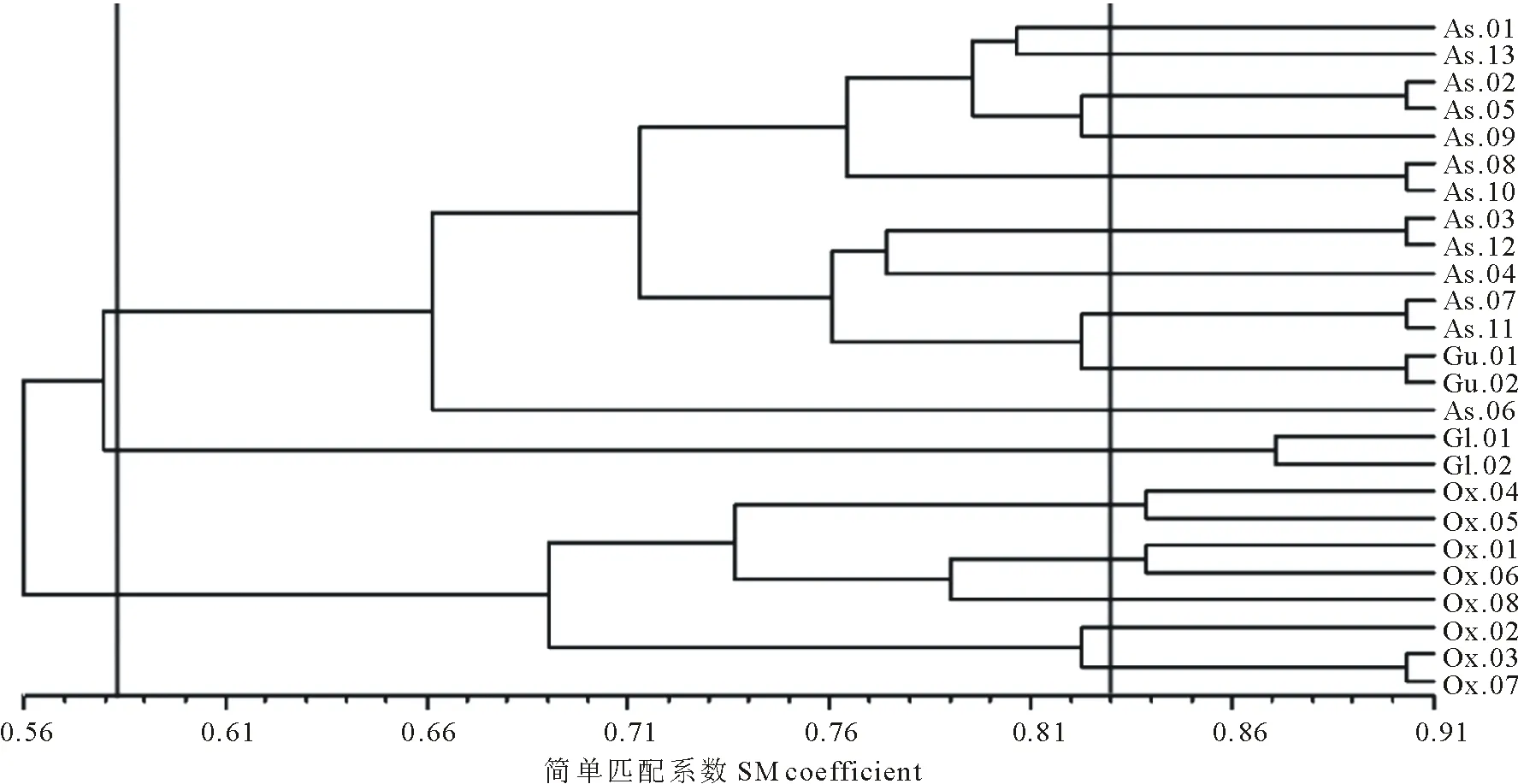
圖3 基于山羊豆族植物31個質量性狀的聚類分析樹狀圖(UPGMA)Fig.3 Cluster analysis dendrogram (UPGMA) based on 31 qualitative characters of tribe Galegeae
基于第1主成分和第2主成分繪制二維散點圖(圖4),與聚類分析結果(圖3)十分相近,4屬植物能較好地被分為3個組。黃耆屬13個物種和米口袋屬2個物種分布比較緊密,區分不明顯。第1主成分坐標軸上甘草屬2個物種和第2主成分坐標軸上棘豆屬8個物種的分布比較分散,說明對此2屬屬下種間具有較高的分類學價值。
3 討論與結論
本研究選取的23個葉形態結構的質量性狀,對多份材料進行分析和歸納,部分葉外觀形態特征與前人研究結果[19-20]基本相似。基于31個質量性狀進行的聚類分析和主成分分析結果,對研究山羊豆族4個屬的屬間及種間關系進行了探討。
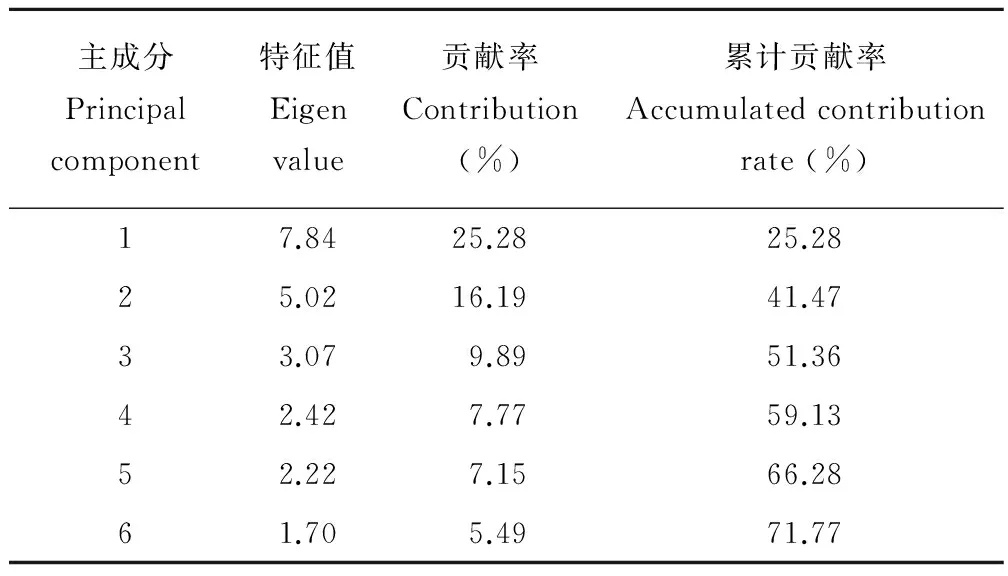
表3 基于山羊豆族植物31個質量性狀的主成分分析
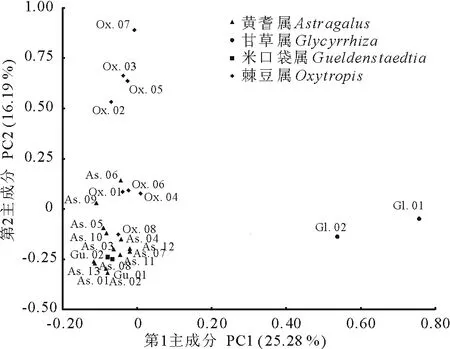
圖4 基于山羊豆族植物31個質量性狀的前2個主成分構建的二維散點圖Fig.4 2-dimensional plot based on the first two principal components of 31 qualitative characters of tribe Galegeae
3.1 黃耆屬
傅坤俊等[19]因小葉具丁狀毛等特征將新巴黃耆、糙葉黃耆及乳白黃耆等劃入裂萼亞屬(subgenusCercidothrix),本研究聚類分析結果中,前兩者分在一起,關系較近,但他們與乳白黃耆較遠。黃耆屬是維管植物第一大屬,形態學與分子系統學研究都認為它是一個多態性類群,現有的分類系統對該屬的屬下類群的劃分尚有爭議[4,14,26-27],而葉的形態特征是否可以作為黃耆屬內的分類依據也有諸多爭論,如Mehrabian等[9]認為黃耆屬Incani組的葉解剖特征與其他組的差異不大,不宜作為組間的分類依據。Haddad和Barnett[25]提出葉柄薄壁細胞和厚角組織細胞數目可以作為黃耆屬一些自然類群的劃分依據。康云和張明理[10]認為葉表皮特征可作為蔟毛黃耆亞屬內組的分類依據。本研究統計分析結果與現有的黃耆屬內亞屬及組的劃分有一些差異,作者認為葉的形態結構可為黃耆屬植物區分種及探討種間親緣關系提供參考。
3.2 甘草屬
本研究揭示刺果甘草和甘草只具無規則型氣孔,可作為區分刺果甘草與甘草和其他物種的依據。但陸嘉惠等[12]提出甘草也具有不等型氣孔,且馮元忠和閆平[11]發現在甘草屬的黃甘草(G.enrycarpa)等物種中也具有不等型氣孔,可見無規則型氣孔只適于探討屬下種間關系,不宜作為區分甘草屬與山羊豆族其他屬的依據。Lewis等[14]通過對甘草屬形態學與分子系統學的研究,認為甘草屬不應歸入山羊豆族。孟雷[28]提出甘草屬與山羊豆族其他屬在系統樹上處于不同的分支,同樣認為將其放在山羊豆族中不太合適。本研究聚類分析表明SM系數為0.58時,甘草屬與其他3屬分開。主成分分析中第1主成分可知,小葉具腺體、葉柄形狀為戟形及葉柄細胞含單寧等特征將刺果甘草與甘草和其他3屬分開,距離較大,這在一定程度上反映甘草屬植物的確與其他3屬有差別。
3.3 米口袋屬
聚類分析可知,米口袋屬與黃耆屬關系密切。狹葉米口袋和少花米口袋具傘形花序[29-30],與山羊豆族其他物種不同,而其他形態特征和黃耆屬的新巴黃耆與糙葉黃耆十分相似,因此葉形態結構不能區分米口袋屬與黃耆屬。主成分分析表明少花米口袋與狹葉米口袋不易區分,前者葉為長橢圓形或披針形,而后者葉在夏秋季時為線形[20,29-30]。本研究還發現兩者的托葉形狀、小葉毛狀體分布也有所不同,可以作為區分兩物種的依據。
3.4 棘豆屬
棘豆屬與黃耆屬關系很近[4,14,20]。高靜等[31]和盧萍等[32-33]通過內蒙古棘豆屬植物分子系統學研究,認為棘豆屬與黃耆屬各成一支,前者應為單系類群。本研究聚類分析結果表明棘豆屬最先被分出,單獨成為一個組。主成分分析結果可以看出,第1主成分坐標軸上棘豆屬、黃耆屬及米口袋屬的關系很近,而第2主成分坐標軸上看,棘豆屬與后兩者之間又存在一些差異,如托葉膜質、小葉葉緣反卷都可作為識別棘豆屬的重要特征,故支持棘豆屬為單系類群。硬毛棘豆、多枝棘豆等葉柄近軸面無凹槽、橫切面形狀及維管束排列成圓形,這些特征相對原始[34],在一定程度上說明棘豆屬較其他3屬原始。
本研究揭示了東北豆科山羊豆族不同類群葉的形態結構,對探討屬間及種間的分類及親緣關系具有重要價值,為形態學與分子系統學揭示的山羊豆族植物不是一個自然類群提供了葉形態結構的依據,并為進一步探討山羊豆族系統學提供了形態學資料。
[1] Hare C L. The anatomy of the petiole and its taxonomic value. Proceedings of the Linnean Society of London, 1943, 155: 223-229.
[2] Metcalfe C R, Chalk L. Anatomy of the Dicotyledons (Vol.1)[M]. Oxford: Clarendon Press, 1950.
[3] Howard R A. The Vascular Structure of the Petiole as a Taxonomic Character[C]. Proceedings of the 15th International Horticultural Congress, Nice, 1958. Pergamon Press, 1962: 7-13.
[4] Lu Y M. Study on Taxonomy and Characters of Leaf Epidermis ofAstragalusL. andOxytropisDC. in Altay Region of Xinjiang[D]. Urumqi: Xinjiang University, 2010.
[5] Sun X Q, Liu M, Sun T H,etal. Morphological stduy of the leaf structures ofViolain northeastern China and discussions of their taxonomic values (Violaceae). Acta Prataculturae Sinica, 2014, 23(2): 223-234.
[6] Cheng X Y, Liu M, Zhang X X,etal. Vegetative organ structures of Ranunculaceae in northeastern China and notes on systematic implications. Acta Prataculturae Sinica, 2014, 23(3): 62-74.
[7] Sun T H, Liu M, Sun X Q,etal. Morphology study on the leaf structures ofPotentillain Northeastern China and its taxonomic value (Rosaceae). Acta Prataculturae Sinica, 2014, 23(3): 75-84.
[8] Shi C Q, Liu M, Wang C,etal. Taxonomic values for leaf structure in the tribe Vicieae (Leguminosae) in northeastern China. Acta Prataculturae Sinica, 2014, 23(6): 157-166.
[9] Mehrabian A R, Zarre S H, Azizian D,etal. Petiole anatomy inAstragalusSect. Incani DC. (Fabaceae) in Iran (a phylogenetical approach). Iran Journal of Botany, 2007, 13(2): 138-145.
[10] Kang Y, Zhang M L. Leaf epidermal feature ofAstragalusL. SubgenusPogonophaceBunge (Leguminosae) from China and its systematic significance. Bulletin of Botanical Research, 2009, 29(1): 25-32.
[11] Feng Y Z, Yan P. Observation on the leaf epidermis stomatal patterns ofGlycyrrhiza. Journal of Shihezi Agricultural College, 1995, 29(1): 1-4.
[12] Lu J H, Li X Y, Zhou L L,etal. Characters of leaf epidermis and their systematic significance inGlycyrrhiza. Acta Botanica Yunnanica, 2005, 27(5): 525-533.
[13] Liu L, Zhao Y Z. A preliminary study on the species ofOxytropisin NeiMonggol by cladistic taxonomic method. Acta Scientiarum Naturalium Universitatis NeiMonggol, 1996, 27(1): 72-82.
[14] Lewis G M, Schrire B, Mackinder B,etal. Legumes of the World[M]. London: Kew Press, 2005.
[15] Ren H Y, Pang Y L, He X J,etal. Characters of leaf epidermis and systematic research inPleurospermumfrom China. Acta Botanica Boreali-Occidentalia Sinica, 2009, 29(1): 49-60.
[16] Dilcher D L. Approaches to the identification of angiosperm leaf remains. The Botanical Review, 1974, 40(1): 154-157.
[17] Chen X, Xing Y, Wang M,etal. Morphology structure of leaf epidermis of genusRubusin Heilongjiang. Acta Botanica Boreali-Occidentalia Sinica, 2012, 32(10): 2035-2039.
[18] Feder N, O’ Brien T P. Plant microtechnique: some principles and new methods. American Journal of Botany, 1968, 55(1): 123-142.
[19] Fu K J, He Y Q, He S B,etal. Flora Tomus 42(1)[M]. Beijing: Science Press, 1993.
[20] Cui H B, Zhang Z W, Xu L R,etal. Flora Tomus 42(2)[M]. Beijing: Science Press, 1998.
[21] Xu K X. Numerical Taxonomy[M]. Beijing: Science Press, 1994.
[22] Zhou H, Liao J, Xia Y P,etal. Morphological characteristics for classifying evergreenAzalea(Ericaceae) cultivars in China using numerical taxonomy. Pakistan Journal of Botany, 2013, 45(2): 593-598.
[23] Mantel N A. The detection of disease clustering and a generalized regression approach. Cancer Research, 1967, 27: 209-220.
[24] Li H Y, Li Z Y, Shi W G,etal. A study on leaf anatomic traits and drought resistance ofMedicagorutenicain Inner Mongolia. Acta Prataculturae Sinica, 2012, 21(3): 138-146.
[25] Haddad R S, Barnett J R. Variation in petiole anatomy of the European spiny species ofAstragalusL. (Leguminosae-Papilionoideae-Galegeae). Botanical Journal of the Linnean Society, 1989, 101: 241-247.
[26] Ma J J. The Taxonomical Study ofAstragalusL. in Inner Mongolia[D]. Hohhot: Inner Mongolia Normal University, 2004.
[27] Wojciechowski M F.Astragalus(Fabaceae): A molecular phylogenetic perspective. Brittonia, 2005, 57(4): 382-396.
[28] Meng L. Systematics ofGlycyrrhizaL. (Fabaceae) - With a Special Reference to Its Relationship toGlycyrrhizopisisBoiss. & Bal.[D]. Beijing: Institute of Botany, the Chinese Academy of Sciences, 2005.
[29] Institute of Forestry and Pedology. Flora Plantarum Herbacearum Chinae Boreali-orientalis Tomus 5[M]. Beijing: Science Press, 1976.
[30] Fu P Y. Clavis Plantarum Chinae Boreali-orientalis Editio Secunda[M]. Beijing: Science Press, 1995.
[31] Gao J, Lu P, Wang J N,etal. Molecular phylogeny of several species ofOxytropisDC. based on 5. 8SrDNA/ITS sequence. Acta Agriculturae Boreali-sinica, 2009, 24(6): 168-173.
[32] Lu P, Gao J, Wang J N,etal. Molecular phylogenetic analysis of severalOxytropisDC. species in Inner mongolia based on 5.8S rDNA/ITS sequences. Acta Botanica Boreali-Occidentalia Sinica, 2011, 31(12): 2420-2428.
[33] Lu P, Gao L X, Jin F,etal. Molecular phylogeny of ten species ofOxytropisbased on psbA-trnH sequences. Plant Diversity and Resources, 2014, 36(3): 279-284.
[34] Rodriguez R L. Systematic anatomical studies onMyrrhidendronand other woody umbellates. University of California Publications in Botany, 1957, 29: 145-318.
參考文獻:
[4] 逯永滿. 新疆阿勒泰地區黃耆屬和棘豆屬分類及葉表皮特征研究[D]. 烏魯木齊: 新疆大學, 2010.
[5] 孫雪芹, 劉玫, 孫天航, 等. 東北堇菜屬植物葉形態結構的研究及其分類學價值的探討. 草業學報, 2014, 23(2): 223-234.
[6] 程薪宇, 劉玫, 張欣欣, 等. 東北毛茛科植物營養器官結構及其系統學意義. 草業學報, 2014, 23(3): 62-74.
[7] 孫天航, 劉玫, 孫雪芹, 等. 東北委陵菜屬植物葉形態結構的研究及其分類學價值的探討. 草業學報, 2014, 23(3): 75-84.
[8] 史傳奇, 劉玫, 王臣, 等. 東北野豌豆族植物葉形態結構的研究及其分類學意義. 草業學報, 2014, 23(6): 157-166.
[10] 康云, 張明理. 中國黃耆屬蔟毛黃耆亞屬的葉表皮特征及其系統學意義. 植物研究, 2009, 29(1): 25-32.
[11] 馮元忠, 閆平. 甘草屬植物葉表面氣孔類型的觀察. 石河子農學院學報, 1995, 29(1): 1-4.
[12] 陸嘉惠, 李學禹, 周玲玲, 等. 甘草屬植物葉表皮特征及其系統學意義. 云南植物研究, 2005, 27(5): 525-533.
[13] 劉麗, 趙一之. 內蒙古棘豆屬植物分支分類的初步探討. 內蒙古大學學報(自然科學版), 1996, 27(1): 72-82.
[15] 任海燕, 逄云莉, 何興金, 等. 中國棱子芹屬植物葉表皮微形態特征及其系統學研究. 西北植物學報, 2009, 29(1): 49-60.
[17] 陳曦, 邢怡, 王蒙, 等. 黑龍江懸鉤子屬植物葉表皮形態結構的研究. 西北植物學報, 2012, 32(10): 2035-2039.
[19] 傅坤俊, 何業祺, 何善寶, 等. 中國植物志第42(1)卷[M]. 北京: 科學出版社, 1993.
[20] 崔鴻賓, 張振萬, 徐朗然, 等. 中國植物志第42(2)卷[M]. 北京: 科學出版社, 1998.
[21] 徐克學. 數量分類學[M]. 北京: 科學出版社, 1994.
[24] 李鴻雁, 李志勇, 師文貴, 等. 內蒙古扁蓿豆葉片解剖性狀與抗旱性的研究. 草業學報, 2012, 21(3): 138-146.
[26] 馬建軍. 內蒙古黃芪屬(AstragalusL.)植物的分類學研究[D]. 呼和浩特: 內蒙古師范大學, 2004.
[28] 孟雷. 甘草屬(GlycyrrhizaL.)的系統學研究-兼論與假甘草屬(GlycyrrhizopsisBoiss. & Bal.)的關系[D]. 北京: 中國科學院植物研究所, 2005.
[29] 遼寧省林業土壤研究所. 東北草本植物志第五卷[M]. 北京: 科學出版社, 1976.
[30] 傅沛云. 東北植物檢索表(第二版)[M]. 北京: 科學出版社, 1995.
[31] 高靜, 盧萍, 王金妞, 等. 基于5. 8SrDNA/ITS序列的幾種內蒙古棘豆屬植物分子系統學研究. 華北農學報, 2009, 24(6): 168-173.
[32] 盧萍, 高靜, 王金妞, 等. 基于5. 8SrDNA/ITS序列的內蒙古棘豆屬植物分子系統學分析. 西北植物學報, 2011, 31(12): 2420-2428.
[33] 盧萍, 高利霞, 金鳳, 等. 基于psbA-trnH序列的十種棘豆屬植物分子系統學研究. 植物分類與資源學報, 2014, 36(3): 279-284.
Leaf morphology and numerical taxonomy of 4 genera of the tribe Galegeae in northeastern China
SHI Chuan-Qi, LIU Mei*, WANG Chen
KeyLaboratoryofMolecularCytogeneticsandGeneticBreedingofHeilongjiangProvince,CollegeofLifeScienceandTechnology,HarbinNormalUniversity,Harbin150025,China
The leaf morphological structure of 25 species,representing 4 genera from the tribe Galegeae, northeastern China, was determined using GMA, SEM and leaf transparency methods. Twenty-three morphological features related to leaf and 8 non-leaf characters were selected for cluster analysis and principal component analysis. Three groups were identified;AstragalusandGueldenstaedtiawere placed in one group, andGlycyrrhizaandOxytropisin separate groups. Features including the presence of glands, petiole shape and tannin content of petiole cells were able to distinguishGlycyrrhizapallidifloraandG.uralensisfrom other Galegeae species. Stipule texture, connectivity between stipule and petiole and leaflet margin features are all useful identification characteristics ofOxytropis. Stipule shape is valuable for interspecific classification. This study provided the leaf morphological evidences for taxonomy of the Galegeae and Leguminosae systematics.
Galegeae; leaf; morphology; structure; taxonomy
10.11686/cyxb20150522
http://cyxb.lzu.edu.cn
2014-11-25;改回日期:2015-01-07
哈爾濱師范大學博士研究生創新科研項目(HSDBSCX2013-06)和國家自然科學基金(31270235及31070169)資助。
史傳奇(1986-),男,黑龍江齊齊哈爾人,在讀博士。E-mail:chuanqi0730@163.com *通訊作者Corresponding author. E-mail:hsd_liumei@163.com
史傳奇,劉玫,王臣. 東北豆科山羊豆族4屬植物葉形態特征及數量分類學研究. 草業學報, 2015, 24(5): 182-189.
Shi C Q, Liu M, Wang C. Leaf morphology and numerical taxonomy of 4 genera of the tribe Galegeae in northeastern China. Acta Prataculturae Sinica, 2015, 24(5): 182-189.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
