水產動物自溶研究進展
2015-05-05 08:48:51宋志遠袁成玉金文剛
食品工業科技 2015年13期
關鍵詞:研究
鄭 杰,宋志遠,于 笛,陳 沖,*,袁成玉,金文剛
(1.遼寧省海洋水產科學研究院,遼寧大連 116023;2.大連海寶生物科技股份有限公司,遼寧大連 116045;3.陜西理工學院生物科學與工程學院,陜西漢中 723001)
水產動物自溶研究進展
鄭 杰1,宋志遠2,于 笛1,陳 沖1,*,袁成玉1,金文剛3
(1.遼寧省海洋水產科學研究院,遼寧大連 116023;2.大連海寶生物科技股份有限公司,遼寧大連 116045;3.陜西理工學院生物科學與工程學院,陜西漢中 723001)
水產動物資源由于水分含量大、自身酶系非常豐富,極易發生自溶,給其儲藏、運輸和加工過程帶來很大不便,是制約水產加工業快速發展的重要瓶頸。本文重點從自溶影響因素、自溶過程生化變化、自溶機理和自溶技術應用四個方面對水產動物自溶的研究現狀進行綜述,并對未來的發展趨勢和研究方向進行了展望,旨在為水產動物自溶的進一步研究提供理論參考。
水產動物,自溶,內源酶
自溶是指當機體受到物理因素(如熱、輻射)、化學因素(如強酸、強堿、有毒物質)和生物因素(如病原體)等環境因素的刺激后,誘發自身酶系將自身的組織結構破壞、降解,從而引起自身死亡的現象[1]。自溶現象在自然界廣泛存在,如動物自溶、植物自溶和微生物自溶等,但不同物種其自溶的定義也有所不同。水產動物通常具有水分含量大、自身酶系非常豐富的特點,因此極易發生自溶現象。水產動物的自溶是指水產動物通過自身體內豐富的酶系成分,在一定條件下使其自身水解的過程[2]。目前,在海洋資源開發和利用已經成為全球各國共同關注的焦點的背景形勢下,國內外學者對水產動物尤其是海參、魚、蝦和貝類等及其加工副產物的自溶進行了廣泛而深入的研究。本文在總結歸納國內外對水產動物自溶的研究成果的基礎上,重點從自溶影響因素、自溶過程生化變化、自溶機理和自溶技術的應用等四個方面對水產動物自溶的研究現狀進行概述,以期為水產動物自溶的研究提供一些新的思路,進而對海洋生物資源的開發利用起到一定的指導和借鑒作用。
1 自溶的影響因素
水產動物自溶是一個系統而又復雜的過程,通常會受到外界環境條件和各種物理化學因素的影響和制約。國內外學者一般采用蛋白質水解度、TCA-可溶性寡肽的含量[3]、可溶性固形物的含量和氨基態氮的變化等作為指標來考察各種因素對水產動物自溶過程的影響,并通過正交實驗、響應面模型等對水產動物自溶條件進行優化,以求能夠實現對自溶過程的有效控制。
1.1 樣品預處理對自溶過程的影響
在水產品加工過程中,為了改善產品的生產工藝和提高產品質量,通常會采用一些不同的技術工藝對原料進行預處理。不同的預處理方式會對水產動物自溶產生不同程度的影響,如經過去皮和水洗之后,緋鯢鰹魚中內源性蛋白酶的含量和活性均有所下降,導致其自溶程度明顯降低[4],相反經過紫外線照射和超聲波處理后,亨氏仿對蝦和刀額新對蝦及魷魚內臟等的內源性蛋白酶被激活,使其自溶速度加快,蛋白質水解率顯著提高[5-7]。但是,紫外線照射時間過長或者超聲波處理時的功率過高均會使內源酶變性失活,從而對水產動物的自溶過程產生抑制作用。
1.2 環境因子對自溶過程的影響
在能夠影響水產動物自溶過程的眾多環境因子中,溫度和pH的影響最為顯著。部分水產動物自溶的最適溫度和最適pH如表1所示。大多數水產動物及其加工副產物自溶的最適溫度為40~65℃,最適pH為6~8,但是也有例外,如緋鯢鰹魚魚肉、魷魚內臟和南極磷蝦自溶的最適pH則在酸性或堿性較強的pH范圍內,分別為4.0、4.5和8.74。溫度和pH等環境因子對自溶過程的影響呈現出與外源酶酶解相類似的變化趨勢,而且最適宜條件與從水產動物體內制備得到的某些內源酶的酶學性質的研究結果相一致,從而為水產動物自溶機理的研究提供了一定的思路。
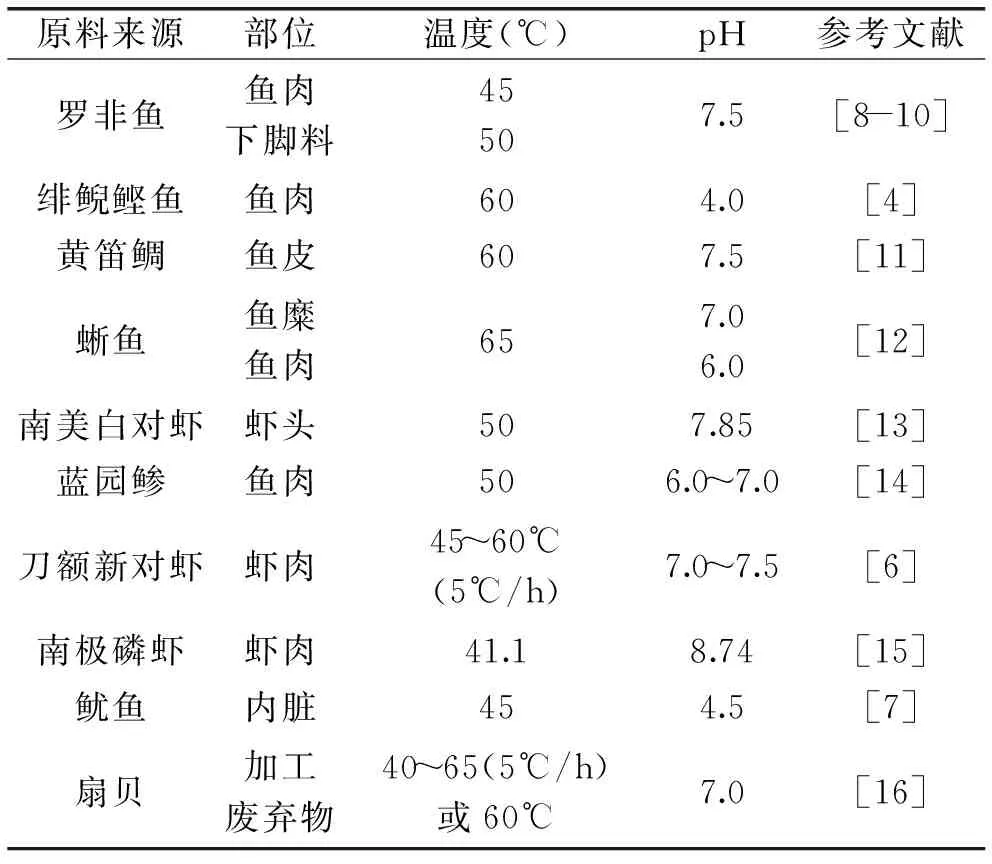
表1 部分水產動物自溶的溫度和pHTable 1 Autolysis temperature and pH of some marine animals
1.3 外源性添加物對自溶過程的影響
在生產過程中,添加食品添加劑是為了能夠獲得質量較高產品而經常采用的一種技術手段。為此,許多學者對多種常用食品添加劑對水產動物自溶過程的影響進行了考察,結果發現,不同種類添加劑對自溶過程有不同程度的影響,而且其作用大小隨添加量的變化而變化,如在葡萄糖≤2.00%、氯化鈉≤10%和檸檬酸≤0.10%的添加量范圍內,對羅非魚蛋白自溶有一定程度的促進作用,超過該范圍則顯示抑制作用[9]。除此之外,其他外源性添加物如雞漿提取物[17]、花生蛋白分離物[18]、乳清蛋白濃縮物(WPC)[19]、卵清蛋白(EW)[20]、牛血漿蛋白(BPP)[21-22]和金屬離子[5]等也可以影響水產動物的自溶過程,而且其影響作用的大小也與添加量有一定的相關性。
2 自溶過程中的生化變化
水產動物自溶過程中,自身所含游離性小分子物質逐漸得到釋放,同時,在豐富內源酶酶系的作用下,大分子的物質逐漸被降解成為小分子物質如肽和氨基酸等。水產動物自溶過程中的生化變化主要體現在α-氨基氮含量、可溶性總氮含量、蛋白質回收率、氨基氮轉化率、總酸、游離氨基酸和游離脂肪酸等方面,也有研究人員對自溶過程中水分的變化規律進行了研究[23]。不同水產動物自溶過程中主要化學成分的變化規律基本類似,大都隨著自溶時間的延長而呈現先逐漸增加后趨于穩定的趨勢,但具體變化規律與自溶的條件有關,如徐偉[24]等對不同條件下鳀魚自溶水解過程中生化變化規律的研究表明,在室溫(26℃)條件下,鳀魚自溶水解的最佳時間為12h,在初始pH為9.0的條件下,自溶水解液中的α-氨基氮含量、可溶性總氮含量、蛋白質回收率和氨基氮轉化率均較高。
3 自溶的機理
水產動物體內通常含有各種各樣的水解酶,水產動物的自溶主要是由于其體內的碳水化合物、脂肪和蛋白質被自身的磷酸酶、脂肪酶、組織蛋白酶和腸酶等水解所引起的,其中,起主要作用的是內源性蛋白酶類。有研究表明,組織蛋白酶B、H和L在降解內源性蛋白質和外源性蛋白質中起到至關重要的作用。因此,國內外學者主要采用外源添加各種蛋白酶抑制因子的手段,從自溶酶尤其是內源性蛋白酶的角度對藍園鲹、鰱魚、緋鯢鰹魚、蜥魚和南美白對蝦等的自溶機理進行了研究,發現在不同種類水產動物自溶過程中起主要作用的內源性酶類有所不同,對其自溶過程起到抑制作用的因子也不盡相同(見表2)。通常所選用的外源蛋白酶抑制因子為TLCK、TPCK、E-64、Antipain和胰蛋白酶抑制劑及胃蛋白酶抑制劑等。酶在水產動物自溶過程中的作用并不是相互獨立的,而是以協同作用的方式進行的,而且一定程度上也會受到外部環境的影響,如在魚死后的僵直階段,滲透壓和離子強度的增高會導致肌原纖維強度的減弱,從而導致其更易發生降解[29]。雖然已經從眾多水產動物體內分離出了組織蛋白酶B[30]和類半胱氨酸蛋白酶[31]及類組織蛋白酶L[32]等內源蛋白酶,也證實了其在自溶過程中起重要作用,但由于水產動物體內內源酶系非常復雜,至于某種或某類內源性蛋白酶在自溶過程中所起到的具體作用,仍需要進行進一步的探討。
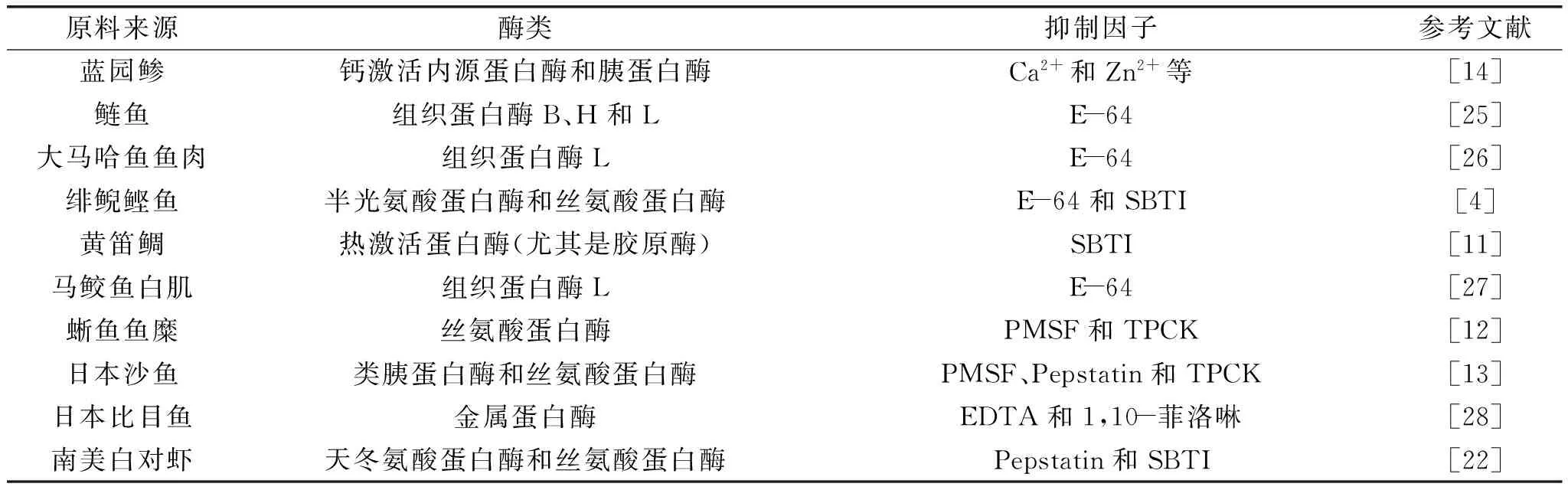
表2 與水產動物自溶相關的部分酶類與抑制因子Table 2 Enzymes and inhibitors related with marine animals autolysis
4 自溶技術的應用
水產動物由于其通常具有水分含量高、酶系豐富的特點而極易發生獨特的自溶現象,然而長期以來,其體內廣泛存在的這種多酶自溶體系一直未得到有效的控制和利用,直到近些年來才逐漸有了改善。由于自溶技術具有成本低、操作簡單和對環境無污染的優點,目前,其已經被廣泛的應用于水產動物中生物活性物質的分離制備、營養成分的回收利用以及相關產品的開發等。
4.1 在生物活性物質制備方面的應用
利用自溶技術或自溶結合外源酶酶解技術制備生物活性肽已經成為水產動物活性肽制備的主要技術手段。研究人員已經采用自溶技術從凡納濱對蝦、海參腸和太平洋無須鱈魚中分別得到了具有ACE抑制活性的肽[33]和具有抗氧化活性的自溶水解物[34-35]。此外,侯佰立[36]和姜淼[37]等采用自溶法,分別從凡納濱對蝦的蝦頭和蝦殼中提取得到了甲殼素和蝦青素。
4.2 在營養成分回收利用方面的應用
采用自溶技術可以實現水產加工副產物中營養成分的有效回收利用,在使原料價值得到最大化利用的同時,也避免了對環境造成污染,如錢俊青[38]等采用自溶法對黃鲇魚內臟中的魚油進行了回收利用;Liu C C[39]等則采用自溶技術對圓花鯖加工廢棄物中的蛋白成分進行了回收,且發現相比外源酶酶解法而言,自溶法的蛋白回收率較高;Senphan[40]等采用自溶技術從南美白對蝦的肝胰腺中提取脂肪,發現60℃自溶60min可以增加脂肪的提取率,而且對脂肪的質量無任何不良影響。
4.3 在產品開發方面的應用
人們最初只能利用自溶來簡單改善食品原料的風味和質構,任其自溶過程緩慢的進行,如傳統的魚露和蝦油的生產等。隨著多年研究的不斷積累,人們開始逐漸利用自溶來開發一些新的產品,如Liu C C[39]等將通過自溶技術取得的低溫自溶回收物進行活性炭脫苦后制成了一種全天然的、可以用于替代味精的增鮮劑;章超樺等[41]采用蝦組織快速自溶新技術對刀額新對蝦蛋白質進行水解,經減壓濃縮和適當調配后制得2種天然海鮮調味料一黑蝦油和蝦味沙司。
5 總結與展望
水產資源通常營養物質含量較高,尤其是海洋動物資源,由于其獨特的高壓、高鹽的生存環境而使其具備許多特殊的生理活性,因此成為國內外科研人員研究的重點。有關水產動物自溶的研究已經有很多,涉及的原料來源有太平洋鱈魚、凡納濱對蝦、刀額新對蝦、海參、扇貝、魷魚、黃鲇魚和南美白對蝦等,其中,各種魚類和蝦類是研究的主要對象。原料形式包括可食部位和加工副產物兩個部分,但是,隨著相關企業降低成本和使原料價值最大化的需求越來越迫切,研究人員也將重點放在了水產動物加工副產物方面(如魚頭、內臟等)。近些年來,人們已經可以從宏觀角度對水產動物自溶過程實現較為有效的控制,并將其充分運用到水產加工企業的實際生產過程中,成為產品開發的一種重要技術手段。
水產動物的自溶過程非常復雜,所發生的生化變化主要是由內源酶系對蛋白和結構組織的降解所導致的,在這一過程中起作用的酶類主要有具有多催化活性的蛋白酶體、溶酶體酶(包括天冬氨酸蛋白酶和組織蛋白酶等)、鈣激活酶、氨基肽酶、堿性蛋白酶和結締組織水解酶如彈性蛋白酶和膠原酶等。雖然目前已經從自溶酶的角度對水產動物自溶過程的機理進行了較多研究,奠定了一定的基礎,但是每種酶在自溶過程中的確切作用目前尚不明確。同時,由于水產動物自溶過程中可能會存在細胞自吞噬現象[42],還應該利用細胞生物學和分子生物學手段分別對自溶過程中的超微細胞結構變化和起關鍵作用的自溶酶基因及其調節作用機制進行進一步的研究,以便從微觀角度更好的揭示自溶這一特殊生理現象的本質。
[1]啟航. 海參體壁酶的研究[D].大連:大連輕工業學院,2007.
[2]鄭杰. 海參自溶過程中生化變化及抗氧化活性寡肽的研究[D].鎮江:江蘇大學,2012.
[3]鄭杰,吳海濤,董秀萍,等. 基于主要化學成分變化建立的海參自溶評價指標[J]. 大連工業大學學報,2010,29(6):391-395.
[4]Yarnpakdee S,Benjakul S,Visessanguan W,et al. Autolysis of goatfish(Mulloidichthysmartinicus)mince:Characterisation and effect of washing and skin inclusion[J]. Food Chemistry,2009,114(4):1339-1344.
[5]章超樺,鄧尚貴,楊麗明,等. 紫外線和溫度對蝦快速白溶的影響-水產品快速自溶影響因素探討之一[J]. 湛江海洋大學學報,1994,14(2):51-56.
[6]章超樺,鄧尚貴. 刀額新對蝦的快速自溶技術[J]. 水產學報,1999,23(4):387-391.
[7]司偉蘭,辛綺婷,余穎兒,等. 魷魚內臟成分分析及自溶條件的研究[J]. 農產品加工·學刊,2010(8):11-14.
[8]黃冬香,李林,于新. 羅非魚蛋白自溶水解條件優化[J]. 西南大學學報:自然科學版,2009,31(5):93-97.
[9]黃冬香,于新,李林. 食品添加劑對羅非魚蛋白自溶作用的影響[J]. 食品科學,2010,31(5):73-77.
[10]孔美蘭,吉宏武,章超樺. 羅非魚下腳料自溶條件的初步探討[J]. 湛江海洋大學學報,2005,25(4):27-31.
[11]Intarasirisawat R,Benjakul S,Visessanguan W,et al. Autolysis study of bigeye snapper(Priacanthusmacracanthus)skin and its effect on gelatin[J]. Food Hydrocolloids,2007,21(4):537-544.
[12]Yongsawatdigul J,Piyadhammaviboon P. Inhibition of autolytic activity of lizardfish surimi by proteinase inhibitors[J]. Food Chemistry,2004,87(3):447-455.
[13]曹文紅,章超樺,洪鵬志,等. 響應面法優化南美白對蝦蝦頭自溶工藝的研究[J]. 中國食品學報,2009,9(1):158-164.
[14]崔春,趙謀明,林偉鋒. 藍園鲹快速自溶機理研究[J]. 食品工業科技,2005,26(2):85-87.
[15]薛勇,趙明明,王超,等. 響應面法優化南極磷蝦蛋白自溶工藝的研究[J]. 食品工業科技,2012(4):346-348,373.
[16]鄭麗,汪秋寬,謝智芬,等. 扇貝加工廢棄物自溶技術的研究[J]. 大連水產學院學報,2007,22(1):49-52.
[17]Rawdkuen S,Benjakul S,Visessanguan W,et al. Fractionation and characterization of cysteine proteinase inhibitor from chicken plasma[J]. Journal of Food Biochemistry,2005,29(5):486-503.
[18]Oujifard A,Benjakul S,Ahmad M,et al. Effect of bambara groundnut protein isolate on autolysis and gel properties of surimi from threadfin bream(Nemipterusbleekeri)[J]. LWT-Food Science and Technology,2012,47(2):261-266.
[19]Rawdkuen S,Benjakul S. Whey protein concentrate:Autolysis inhibition and effects on the gel properties of surimi prepared from tropical fish[J]. Food Chemistry,2008,106(3):1077-1084.
[20]Yongsawatdigul J,Piyadhammaviboon P. Inhibition of autolytic activity of lizardfish surimi by proteinase inhibitors[J]. Food Chemistry,2004,87(3):447-455.
[21]Benjakul S,Visessanguan W,Tueksuban J,et al. Effect of some protein additives on proteolysis and gel-forming ability of lizardfish(Sauridatumbil)[J]. Food Hydrocolloids,2004,18(3):395-401.
[22]Eakpetch P,Benjakul S,Visessanguan W,et al. Autolysis of pacific white shrimp(Litopenaeusvannamei)meat:characterization and the effects of protein additives[J]. Journal of Food Science,2008,73(2):S95-S103.
[23]儀淑敏,王雪琦,紀曉林,等. 刺參自溶過程中水分變化規律的研究[J]. 食品工業科技,2014,35(11):49-51,56.
[24]徐偉,于剛,薛勇. 鳀魚自溶水解過程的生化變化[J]. 食品科技,2010(4):136-139.
[25]李樹紅,張楠,劉歡,等. 鰱魚背肌肌原纖維蛋白自溶與內源組織蛋白酶 B,L,H的關系[J]. 中國農業大學學報,2004,9(5):71-75.
[26]Yamashita M,Konagaya S. Differentiation and localization of catheptic proteinases responsible for extensive autolysis of mature chum salmon muscle(Oncorhynchus keta)[J]. Comparative Biochemistry and Physiology Part B:Comparative Biochemistry,1992,103(4):999-1003.
[27]Aoki T,Ueno R. Involvement of cathepsins B and L in the post-mortem autolysis of mackerel muscle[J]. Food Research International,1997,30(8):585-591.
[28]Kubota M,Kinoshita M,Kubota S,et al. Possible implication of metalloproteinases in post-mortem tenderization of fish muscle[J]. Fisheries Science,2001,67(5):965-968.
[29]Delbarre-Ladrat C,Cheret R,Taylor R,et al. Trends in postmortem aging in fish:Understanding of proteolysis and disorganization of the myofibrillar structure[J]. Critical Reviews in Food Science and Nutrition,2006,46(5):409-421.
[30]Sun L M,Zhu B W,Wu H T,et al. Purification and characterization of cathepsin B from the gut of the sea cucumber(Stichopusjaponicas)[J]. Food Science and Biotechnology,2011,20:919-925.
[31]Qi H,Dong X P,Cong L N,et al. Purification and characterization of a cysteine-like protease from the body wall of the sea cucumberStichopusjaponicus[J]. Fish Physiology and Biochemistry,2007,33:181-188.
[32]Zhou D Y,Chang X N,Bao S S,et al. Purification and partial characterisation of a cathepsin L-like proteinase from sea cucumber(Stichopusjaponicus)and its tissue distribution in body wall[J]. Food Chemistry,2014,158:192-199.
[33]郝更新,章超樺,曹文紅. 對蝦蛋白自溶制備ACE 抑制肽的工藝條件優化[J]. 湛江海洋大學學報,2006,26(4):59-62.
[34]鄭杰,吳海濤,朱蓓薇,等. 海參腸自溶水解物抗氧化活性的研究[J]. 大連工業大學學報,2011,30(5):313-317.
[35]Samaranayaka A G P,Li-Chan E C Y. Autolysis-assisted production of fish protein hydrolysates with antioxidant properties from Pacific hake(Merlucciusproductus)[J]. Food Chemistry,2008,107(2):768-776.
[36]侯佰立,吉宏武,王燕,等. 凡納濱對蝦蝦頭制備甲殼素工藝的研究[J]. 食品工業科技,2011,(10):273-276.
[37]姜淼,楊賢慶,李來好,等. 內源酶輔助提取蝦殼蝦青素的研究[J]. 南方水產科學,2011,7(2):55-60.
[38]錢俊青,單昱東,廖啟元. 黃鲇魚內臟自溶酶解提取魚油的工藝[J]. 食品與發酵工業,2009,35(12):66-69.
[39]Liu C C,Morioka K,Itoh Y,et al. Autolysis:An effective way to recover protein from fish solid wast[J]. Journal of Shanghai Fisheries University,2003,12(suppl):71-77.
[40]Senphan T,Benjakul S. Compositions and yield of lipids extracted from hepatopancreas of Pacific white shrimp(Litopenaeusvannamei)as affected by prior autolysis[J]. Food Chemistry,2012,134(2):829-835.
[41]章超樺,鄧尚貴. 蝦組織快速自溶技術在海鮮調味料生產上的應用研究[J]. 食品與發酵工業,2000,26(2):36-39.
[42]Zhu B W,Zheng J,Zhang Z S,et al. Autophagy plays a potential role in the process of sea cucumber body wall “melting” induced by UV irradiation[J]. Wuhan University Journal of Natural Sciences,2008,13(2):232-238.
Research progress in the autolysis of aquatic animals
ZHENG Jie1,SONG Zhi-yuan2,YU Di1,CHEN Chong1,*,YUAN Cheng-yu1,JIN Wen-gang3
(1.Aquatic Products Processing Laboratory,Liaoning Ocean and Fisheries Science Research Institute,Dalian 116023,China;2.Dalian Haibao Biotechnology Co,Ltd.,Dalian 116045,China;3.School of Bioscience and Engineering,Shaanxi University of Technology,Hanzhong 723001,China)
The aquatic animals are easily subjected to autolysis because of its high content of water and endogenous enzymes,and this phenomenon brings great difficulties to its storage,transportation and processing. It has become the bottleneck of aquatic products processing. In this paper,the research status of autolysis was summarized from the aspects of influence factors,biochemical changes,autolytic mechanism and autolysis technology applications. The developing trend was also prospected,and this may provide theoretic basis for the further studies of aquatic animals autolysis.
aquatic animals;autolysis;endogenous enzyme
2014-09-24
鄭杰(1981-),男,博士,助理研究員,研究方向:水產品精深加工與綜合利用。
*通訊作者:陳沖(1963-),男,本科,研究員,研究方向:水產品精深加工與綜合利用。
遼寧省海洋與漁業廳科研項目(201207)。
TS254.4
A
1002-0306(2015)13-0397-04
10.13386/j.issn1002-0306.2015.13.076
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
