CUS致大鼠抑郁行為涉及單胺遞質合成酶表達下調
2015-06-09 14:25:43汪麗佳邱紅梅蔣心惠費慧芝胡小婭周岐新
中國藥理學通報 2015年4期
汪麗佳,邱紅梅,蔣心惠,劉 丹,費慧芝,胡小婭,周岐新
(1.重慶市生物化學和分子藥理學重點實驗室,重慶 400016;2.重慶市中山醫院,重慶 400015;3.重慶市三峽醫學高等專科學校,重慶 404100)
CUS致大鼠抑郁行為涉及單胺遞質合成酶表達下調
汪麗佳1,邱紅梅1,蔣心惠1,劉 丹2,費慧芝3,胡小婭1,周岐新1
(1.重慶市生物化學和分子藥理學重點實驗室,重慶 400016;2.重慶市中山醫院,重慶 400015;3.重慶市三峽醫學高等專科學校,重慶 404100)
目的 探討慢性不可預知性刺激(chronically unpredicted stress,CUS)致大鼠抑郁行為與單胺遞質合成限速酶色氨酸羥化酶(tryptophan hydroxylase,TPH)和酪氨酸羥化酶(tyrosine hydroxylase,TH)表達的關系。方法 ♂SD大鼠30只,隨機分為對照組(control group,CG)和模型組(model group,MG),每組15只。采用連續給予CUS結合孤養28 d建立抑郁模型;對照組每籠5只,正常飼養;以開場試驗和糖水消耗實驗進行行為學評價;以熒光定量PCR測定海馬、前腦皮層TPH 及TH mRNA表達,以Western blot法測定海馬、前腦皮層TPH 及TH蛋白表達。結果 與CG相比,MG大鼠開場實驗水平和垂直活動得分均明顯降低,糖水消耗量也顯著減少,海馬、前腦皮層TPH及TH mRNA和蛋白表達均明顯下降。結論 CUS誘導♂大鼠產生抑郁癥樣行為,其發生機制可能與單胺遞質合成限速酶TPH、TH表達下調相關。
抑郁癥;慢性不可預見性刺激;色氨酸羥化酶;酪氨酸羥化酶;5-羥色胺;去甲腎上腺素
抑郁癥的發病機制存在眾多假說。早期研究認為,抑郁癥的發生與神經單胺遞質缺乏相關。隨著研究的深入,進一步發現氧化/抗氧化應激損傷、下丘腦-垂體-腎上腺軸(hypothalamic- pituitary-adrenal axis,HPA)功能失調、神經營養因子減少及炎癥反應等都參與了抑郁癥的發生[1-3]。其中,“單胺學說”是目前抑郁癥發病機制最重要的假說之一。該學說認為突觸間隙的5-羥色胺(serotonin,5-HT)及去甲腎上腺素(norepinephrine,NE)水平低下是導致抑郁癥的重要原因[1]。單胺神經遞質發揮作用需經歷遞質合成、轉運、代謝過程。通過阻止神經末梢對遞質再攝取或抑制遞質降解而研發出一系列臨床行之有效的經典抗抑郁藥物,如選擇性5-羥色胺再攝取抑制劑(selective serotonin reuptake inhibitor, SSRI)和單胺氧化酶抑制劑等。但即便如此,作為臨床廣泛使用的SSRI等治療抑郁癥通常需要幾周才能完全發揮作用,并且約有60%~70%的病人治療無效[4],其原因尚不明確。有報道認為,大腦單胺遞質缺乏的小鼠對SSRI治療無應答[5]。因此,增加突觸間隙單胺遞質濃度是否可以通過增加遞質合成達到抗抑郁治療值得考慮。TPH和TH分別是合成5-HT以及兒茶酚胺神經遞質(去甲腎上腺素、腎上腺素、多巴胺)的限速酶。有研究發現,TPH基因多態性與抑郁癥關系密切,并提出將TPH作為抗抑郁治療的新靶點[6]。抑制TH活性可產生抑郁樣行為,增加大腦TH水平可用于治療抑郁癥[7]。因此,本文從單胺遞質的合成出發,采用CUS誘導大鼠產生抑郁行為,觀察5-HT和NE的合成限速酶表達是否參與了抑郁癥的發病機制。
1 材料與方法
1.1 動物清潔級♂Sprague-Dawley(SD)大鼠30只,180~220 g,重慶醫科大學動物實驗中心提供,醫學動物許可證號:SCXK(渝)2007-0001。
1.2 主要試劑與儀器PCR引物(上海鼎安生物技術有限公司);RNA提取試劑盒Total RNA Kit I(OMEGA公司);PrimeScriptTMRT reagent Kit(TaKaRa公司,大連寶生物);逆轉錄試劑盒(TaKaRa公司,大連寶生物);anti-TH、anti-TPH(美國Millipore公司);WB山羊抗兔IgG二抗(北京中杉);ECL發光液:Super Signal Western Blotting Kits(Pierce公司);實時定量PCR儀:美國Bio-rad公司(型號:MJ7500);凝膠成像系統:Bio-rad公司(型號:170-1870);WB垂直電泳儀:美國Bio-rad公司;低溫離心機(Eppendoff)。
1.3 分組及飼養大鼠自由攝食飲水,正常光照節律。在室溫(20±3)℃實驗環境下適應性喂養7 d后,采用open field法篩選大鼠,選取水平活動得分30~120之間的大鼠隨機分為2組,即對照組(CG)和模型組(MG),每組15只;CG組5只/籠, MG組每只大鼠單籠喂養。
1.4 孤養結合CUS方法建立大鼠抑郁模型參照Willner等[8]方法并加以改進,建立實驗性大鼠抑郁模型。刺激方式包括禁食禁水24 h, 夾尾1 min, 間歇照明(光/暗周期為2 h/2 h,時間19 ∶00 ~次日7 ∶00),通宵(12 h)照明,傾斜鼠籠45°結合禁水24 h, 45 ℃熱環境5 min, 4 ℃冰水游泳5 min,電刺激(36 V,4 Hz)5 min, 噪音1 h, 空盒無墊料結合禁食24 h,潮濕墊料24 h,共11種刺激,大鼠每天接受一種刺激,每種刺激累計出現2 ~3次,同種刺激不連續出現,使動物不能預料何種刺激會發生,實驗周期為28 d。
1.5 開場實驗模型建立后,于d 2早上7 ∶30進行開場實驗。參考文獻[9],使用自制100 cm×100 cm×40 cm木箱,底部平均分為5×5個方格,木箱內全部漆成黑色。保持環境安靜,將大鼠置于木箱中央,觀察大鼠5 min內活動情況,記錄大鼠的水平得分(四爪均進入方格的穿越格數)、垂直得分(后肢直立次數)。觀察完畢后,酒精清潔敞箱后再進行下一次觀察,以免受上次大鼠遺留氣味的影響。
1.6 糖水消耗實驗參照文獻,在模型建立d 26,先訓練大鼠試飲10 g·L-1蔗糖水24 h,d 27飲純水24 h,訓練期間正常進食。訓練結束后開始正式實驗:禁食禁水24 h后,所有大鼠單籠放置,同時給予10 g·L-1蔗糖水和純水各1瓶,實驗前稱取帶水瓶重。2 h后,再次稱量帶水瓶重并記算蔗糖水和純水消耗量,最終計算糖水偏好率(糖水偏好率/%=糖水消耗/總液體消耗×100 %)。
1.7 TPH、TH mRNA表達測定造模結束,每組取5只大鼠,經40 g·L-1水合氯醛(400 mg·kg-1, ip)麻醉后斷頭取腦組織,冰上分離前腦皮層和海馬,置于裝有RNAstore組織保存液(每100 mg組織加入1 mlRNAstore)EP管。4℃過夜,移出液體,組織于-80℃保存。TH引物:上游5′-AGAGGACAGCATCCCACAGC-3′,下游5′-ATCACGGGCGGACAGTAGA-3′;TPH引物:上游5′-TTTGTAGCCAACATTCCTCA-3′,下游5′-ACTATTGAAAGTAGAAACCACCTC-3′;β-actin作內參。擴增產物大小分別為100 bp, 132 bp和100 bp。以TRIzol法按試劑盒說明書提取樣本總RNA, 用TIANGEN試劑盒進行逆轉錄。熒光定量聚合酶鏈反應參見試劑盒說明書:1 μL DNA樣本作為 PCR 擴增模板,陰性對照擴增模板為去離子水;擴增條件為95 ℃預變性2 min; 隨后95 ℃ 10 s,退火15 s,72 ℃ 延伸45s,共40個循環;最后72 ℃延伸10 min。本試驗采用⊿⊿CT法進行實時定量PCR相對定量反應,樣本均采用的三復管。取平均值作為最終試驗結果,然后與內參進行比較,計算相對含量。
1.8 TPH、TH蛋白表達測定按“1.7”方法處理大鼠,冰上分離出前腦皮層和海馬,剪碎組織,加裂解液處理,提取蛋白,采用BCA法定量蛋白濃度。采用Western blot 方法檢測兩腦區TH和TPH蛋白表達。每孔上樣量為50 μg, 進行SDS-PAGE凝膠電泳,轉膜,室溫封閉1 h,加入一抗4℃過夜、洗膜,加入二抗,室溫封閉30 min后洗膜,凝膠成像系統取像,以β-actin蛋白做為內參校正蛋白表達量。
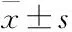
2 結果
2.1 大鼠行為變化開場實驗結果顯示,與CG組相比,CUS致大鼠(MG組)水平得分和垂直得分均明顯降低(P<0.01,P<0.05),見Fig 1。糖水實驗結果顯示,MG組大鼠糖水消耗量明顯低于CG組(P<0.05),見Tab 1,提示CUS成功誘導大鼠產生焦慮、興趣缺失等抑郁樣行為。
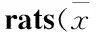
Tab 1 Effect of CUS on the sucrose preference of ±s,n=15)
*P< 0.05vsCG
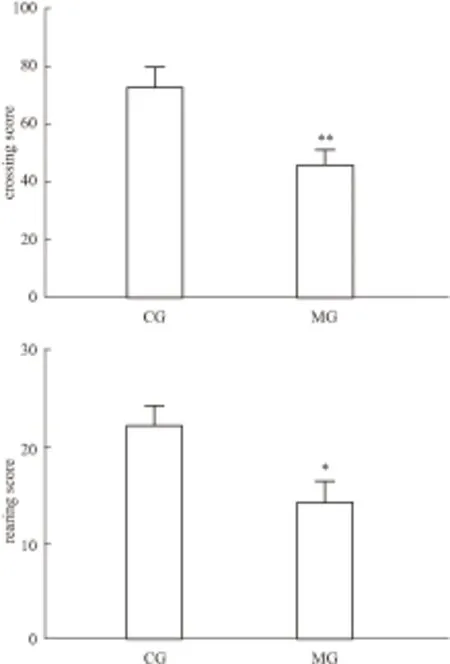
*P<0.05,**P<0.01vsCG
2.2 大鼠腦內TPH mRNA及蛋白表達與CG組比,MG組大鼠海馬和前腦皮層TPH mRNA表達量明顯降低(海馬P<0.01,皮層P<0.05),見Tab 2,同時MG組海馬和前腦皮層TPH蛋白表達也明顯下降(P<0.05),見Fig 2。提示CUS誘導大鼠產生抑郁行為時,使海馬和前腦皮層5-HT合成限速酶表達量明顯降低。
2.3 大鼠腦內TH mRNA及蛋白表達與CG組相比,MG組大鼠海馬和前腦皮層TH mRNA表達量明顯降低(海馬P<0.01,皮層P<0.05),見Tab 2。,同時MG組海馬和前腦皮層TH蛋白表達也明顯下降(海馬P<0.05,皮層P<0.01),見Fig 3,結合TPH表達結果,顯示CUS能明顯抑制大鼠單胺合成限速酶TPH和TH的表達。
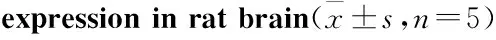
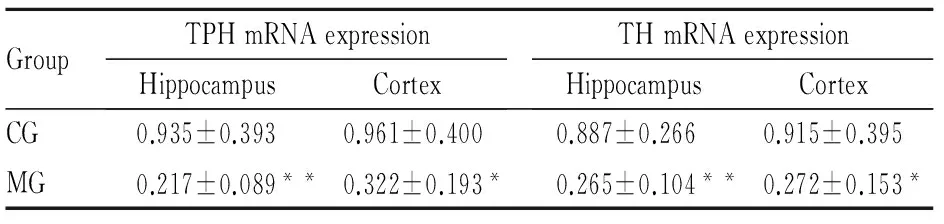
GroupTPHmRNAexpressionHippocampusCortexTHmRNAexpressionHippocampusCortexCG0.935±0.3930.961±0.4000.887±0.2660.915±0.395MG0.217±0.089**0.322±0.193*0.265±0.104**0.272±0.153*
*P<0.05,**P<0.01vsCG
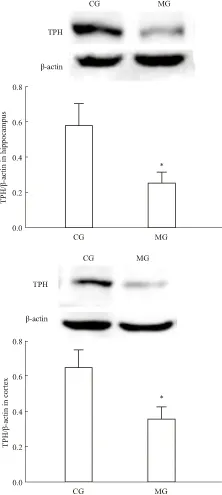
Fig 2 Effect of CUS on TPH protein
*P<0.05vsCG
3 討論
CUS是目前經典、常用的實驗鼠抑郁模型建立方法。其主要原理是模擬人類抑郁癥形成中受到的不同慢性低水平的應激源刺激。開場實驗和糖水消耗實驗可分別測定實驗動物焦慮抑郁行為和是否存在快感缺失。為此本課題組以CUS誘導建立大鼠抑郁模型,對腦內單胺遞質合成、再攝取和代謝進行了系列研究。前期工作發現,CUS誘導的抑郁模型鼠存在轉運體表達異常,文拉法辛和瑞波西汀能逆轉這種表達異常而改善大鼠抑郁癥狀[9-10]。然而,針對臨床使用SSRI等抗抑郁藥物的低治愈率使得人們有理由考慮,抑郁癥發生機制是否也與單胺遞質合成酶表達異常有關。為此,采用經典的CUS結合孤養方法建立大鼠抑郁癥模型是探討抑郁行為與單胺遞質合成酶表達關系的最簡潔明了的途徑。
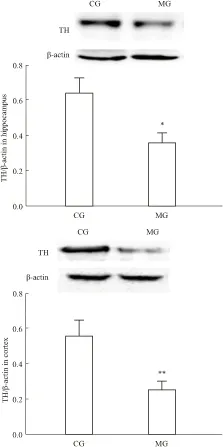
Fig 3 Effect of CUS on TH protein
*P<0.05,**P<0.01vsCG
本研究結果顯示,經CUS刺激的大鼠開場實驗水平得分和垂直得分均明顯低于對照組;糖水消耗實驗也顯示CUS大鼠糖水攝取量明顯低于對照組,提示CUS誘導大鼠出現焦慮、快感缺失等抑郁行為。本研究還發現, 模型組大鼠海馬和前腦皮層TPH以及TH mRNA和蛋白表達均明顯低于對照組。已知大鼠海馬和前腦皮層與認知功能、學習記憶和情感等密切相關,提示CUS致大鼠焦慮抑郁可能涉及該腦區的TPH和TH表達下調。
TPH和TH分別是5-HT和NE合成的限速酶。5-HT作為重要的神經遞質作用于HPA軸、參與涉及交感神經系統的壓力應答,其功能失調會引起精神障礙。TPH作為5-HT的合成限速酶,有報道認為,5-HT前體物質色氨酸不足或限速酶TPH活性降低可誘發抑郁癥狀[11]。采用母嬰分離方法建立幼鼠抑郁模型,發現其成年后中縫核TPH表達降低。此外,也有多篇TPH基因多態性涉及抑郁癥發生的報道,并認為可將TPH作為抗抑郁藥物研究的新靶點[6]。這些均支持TPH表達下調與抑郁癥發生的確有關。最近報道,傳統中藥制劑開心丸在升高海馬及前腦皮層5-HT水平的同時,明顯增加TPH的表達[12]。同樣,有報道提升TH表達可改善在強迫游泳和懸尾實驗中大鼠的絕望行為[13],電休克抗抑郁治療與明顯提高TH基因表達有關[14]。結合TPH的報道,在本次研究中模型組大鼠海馬和前腦皮層TPH、TH mRNA及蛋白表達均降低,意味相關區域的腦神經5-HT、NE遞質合成減少,進而釋放到突觸間隙的5-HT和NE也減少;這可能與抑郁癥的“單胺學說”發生機制不謀而合。因此,單胺遞質合成酶是否可以作為單胺遞質再攝取抑制劑低療效的補充治療靶點值得進一步探討。
[1] Iversen L. Neurotransmitter transporters and their impact on the development of psychopharmacology[J].BrJPharmacol, 2006, 147(S1): 82-8.
[2] Castren E, Rantamaki T. The role of BDNF and its receptors in depression and antidepressant drug action: Reactivation of developmental plasticity[J].DevNeurobiol, 2010, 70(5): 289-97.
[3] Kim Y K,Na K S,Shin K H,et al. Cytokine imbalance in the pathophysiology of major depressive disorder[J].ProgNeuropsychopharmacolBiolPsychiatry, 2007, 31(5): 1044-53.
[4] de Bodinat C, Guardiola L B, Mocaer E, et al. Agomelatine, the first melatonergic antidepressant: discovery, characterization and development[J].NatRevDrugDiscov, 2010, 9(8):628-42.
[5] Cryan J F, O’Leary O F, Jin S H, et al. Norepinephrine deficient mice lack responses to antidepressant drugs, including selective serotonin reuptake inhibitors[J].ProcNatAcadSci, 2004, 101(21): 8186-91.
[6] Chen G L,Miller G M. Tryptophan hydroxylase-2: An emerging therapeutic target for stress disorders[J].BiochemPharmacol, 2013, 85(9):1227-33.
[7] Thony B, Auerbach G, Blau N. Tetrahydrobiopterin biosynthesis, regeneration and functions[J].BiochemJ,2000, 347(1):1-16.
[8] Willner P, Benton D, Brown E, et al. Depression increases “craving” for sweet rewards in animal and human models of depression and craving[J].PsychopharmacolBerl,1998,136(3):272-83.
[9] 李 娜,王 涵,文 威,等. 改善氧化/抗氧化應激平衡及NET和5-HTT表達可能與瑞波西汀抗抑郁作用有關[J].中國藥理學通報,2011,27(9):593-7.
[9] Li N, Wang H, Wen W, et al. Improvement of oxidative/anti-oxidative stress balance and expressions of NET and 5-HTT involved in antidepressive effect of reboxetine[J].ChinPharmacolBull, 2011, 27(9):593-7.
[10] 胡小婭,費慧芝,蔣心惠,等. 改善氧化應激平衡和逆轉NET及5-HTT表達異常涉及文拉法辛的抗抑郁作用[J].中國藥理學通報,2014,30(6):848-52.
[10] Hu X Y, Fei H Z, Jiang X H, et al. Improvement of oxidative/anti-oxidative stress balance and reversion of NET and 5-HTT abnormal expression involved in the anti-depressive effect of venlafaxine[J].ChinPharmacolBull, 2014, 30(6):848-52.
[11] Yang F Z, Wu Y, Zhang W G, et al. Influence of estradiol on tryptophan hydroxylase and 5-hydroxytryptamine content in raphe nuclei of rats under forced swimming stress[J].ZhonghuaYiXueZaZhi, 2010, 90(27): 1929-32.
[12] Dong X Z, Li Z L, Zheng X L, et al. A representative prescription for emotional disease, Ding-Zhi-Xiao-Wan restores 5-HT system deficit through interfering the synthesis and transshipment in chronic mild stress-induced depressive rats [J].JEthnopharmacol, 2013, 150(3):1053-61.
[13] Fu A L, Wu S P, Dong Z H, et al. A novel therapeutic approach to depression via supplement with tyrosine hydroxylase[J].BiochemBiophysResCom, 2006,351(1):140-5.
[14] Brady L S, Lynn A B, Glowa J R, et al. Herkenham,Repeated electroconvulsive shock produces long-lasting increases in messenger RNA expression of corticotropin-releasing hormone and tyrosine hydroxylase in rat brain [J].TherImplJClinInvest, 1994, 94(3):1263-8.
[15] Zhang J, Darling R D, Paul I A, et al. Altered expression of tyrosine hydroxylase in the locus coeruleus noradrenergic system in citalopram neonatally exposed rats and monoamine oxidase a knock out mice[J].AnatRec(Hoboken), 2011,294(10):1685-97.
☆ ☆ ☆ ☆ ☆ ☆ ☆ ☆ ☆ ☆ ☆ ☆ ☆ ☆ ☆ ☆ ☆ ☆ ☆ ☆ ☆ ☆ ☆ ☆
《中國藥理學通報》投稿信箱:zgylxtb8@163.com;
《中國藥理學通報》修回稿信箱:zgylxtb88@163.com;
《中國藥理學通報》稿件查詢專用信箱:zgylxtb@163.com
Down-regulation of monoamine neurotransmitter synthetase expression involved in CUS-induced depression of rats
Wang Li-jia1, Qiu Hong-mei1, Jiang Xin-hui1, Liu Dan2, Fei Hui-zhi3, Hu Xiao-ya1, Zhou Qi-xin1
(1.DeptofBiochemistryandMolecularPharmacology,Chongqing400016,China;2.ChongqingZhongshanHospital,Chongqing400015,China;3.ChongqingThreeGorgesMedicalCollege,Chongqing404100,China)
Aim To investigate whether chronically unpredictable stress(CUS)-induced depression-like behaviors of rats is associated with the variant expression of tryptophan hydroxylase (TPH) and tyrosine hydroxylase (TH). Methods 30 male SD rats were randomly divided into depression model group(MG) and control group(CG),the former was established using CUS plus solitary condition for 28 d, whereas the latter was fed normally as five rats per cage without CUS. The open field test(OFT)and the sucrose preference test were used to evaluate depressive behaviors. Both mRNA and protein expressions of TPH and TH in hippocampus and forebrain cortex were determined by real-time fluorescent quantitative PCR and western blot (WB), respectively. Results MG rats showed obvious depressive behaviors with much lower locomotive activity and sucrose preference than CG. Meanwhile, the mRNA and protein expressions of TPH and TH also significantly decreased in MG rats, compared with CG rats. Conclusion The depression behaviors of rats induced by CUS may be associated with down-regulation of TPH and TH expression.
depression;CUS;TPH;TH;5-HT;NE
時間:2015-3-16 15:41 網絡出版地址:http://www.cnki.net/kcms/detail/34.1086.R.20150316.1541.026.html
2014-11-03,
2015-02-22
國家自然科學基金資助項目(No 31400881);重慶市教委科學技術研究項目(No KJ1400208)
汪麗佳(1987-),女,碩士生,研究方向:神經精神藥理學,E-mail:492759231@qq.com; 周岐新(1947-),男,教授,博士生導師,研究方向:神經精神藥理學,E-mail:cqzhouqx@cqmu.edu.cn
10.3969/j.issn.1001-1978.2015.04.018
A
1001-1978(2015)04-0527-05
R-332;R322.81;R338.1;R345.47;R749.42;R977.3
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
作文周刊·小學二年級版(2022年20期)2022-05-05 01:33:06
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
創新作文(小學版)(2019年10期)2019-09-25 08:12:28
小學生學習指導(低年級)(2017年5期)2017-05-04 04:14:38
光學精密工程(2016年6期)2016-11-07 09:07:19
發明與創新(2016年38期)2016-08-22 03:02:52
