轉(zhuǎn)運(yùn)真核表達(dá)盒的重組T7噬菌體構(gòu)建
2015-06-09 14:20:33王義偉許夢(mèng)薇侯繼波
江蘇農(nóng)業(yè)學(xué)報(bào) 2015年1期
徐 海, 鮑 熹, 王義偉, 盧 宇, 許夢(mèng)薇, 侯繼波
(江蘇省農(nóng)業(yè)科學(xué)院/國(guó)家獸用生物制品工程技術(shù)研究中心,江蘇南京 210014)
轉(zhuǎn)運(yùn)真核表達(dá)盒的重組T7噬菌體構(gòu)建
徐 海, 鮑 熹, 王義偉, 盧 宇, 許夢(mèng)薇, 侯繼波
(江蘇省農(nóng)業(yè)科學(xué)院/國(guó)家獸用生物制品工程技術(shù)研究中心,江蘇南京 210014)
為了構(gòu)建轉(zhuǎn)運(yùn)真核表達(dá)盒的重組T7噬菌體,并分別在體外、體內(nèi)條件下檢測(cè)其標(biāo)簽蛋白質(zhì)EGFP的表達(dá)。以T7噬菌體基因組左側(cè)578 bp處為插入位點(diǎn),取上游400 bp和下游200 bp片段作為左右同源臂,并在其中插入表達(dá)EGFP基因的真核表達(dá)盒(EEB),構(gòu)建同源重組質(zhì)粒pUC-L-EEB-R。將該質(zhì)粒載體轉(zhuǎn)化T7噬菌體宿主細(xì)菌BL21,在噬菌體繁殖過(guò)程中完成同源重組,PCR篩選重組T7-EEB噬菌體。提取該重組噬菌體基因組轉(zhuǎn)染Vero細(xì)胞后體外檢測(cè)EGFP蛋白質(zhì)表達(dá),純化的噬菌體免疫小鼠后體內(nèi)檢測(cè)EGFP蛋白質(zhì)表達(dá)。結(jié)果顯示,通過(guò)同源重組方法成功構(gòu)建了攜帶真核表達(dá)盒的重組T7噬菌體,PCR檢測(cè)和酶切鑒定均證明表達(dá)盒已正確插入。T7-EEB基因組轉(zhuǎn)染真核細(xì)胞可見明顯的EGFP蛋白質(zhì)表達(dá),免疫小鼠后活體熒光檢測(cè)到EGFP蛋白質(zhì)信號(hào),在小鼠肝臟、脾臟組織中RT-PCR檢測(cè)到EGFP基因的mRNA轉(zhuǎn)錄。表明同源重組方法可以用于構(gòu)建重組T7噬菌體,噬菌體能夠轉(zhuǎn)運(yùn)真核表達(dá)盒并實(shí)現(xiàn)蛋白質(zhì)表達(dá)。
同源重組;DNA疫苗;真核表達(dá);T7噬菌體
DNA疫苗是20世界90年代發(fā)展起來(lái)的一種新型疫苗,是指將含有編碼靶抗原基因的質(zhì)粒載體導(dǎo)入宿主,通過(guò)宿主細(xì)胞表達(dá)抗原從而誘導(dǎo)機(jī)體產(chǎn)生免疫反應(yīng)。該免疫途徑不僅能激發(fā)體液免疫反應(yīng),還能誘導(dǎo)高水平的細(xì)胞免疫,尤其是細(xì)胞毒性T淋巴細(xì)胞(CTL)反應(yīng),在疾病的防治中具有較大優(yōu)勢(shì)[1-2]。
DNA疫苗有多種免疫途徑:肌肉、皮內(nèi)、皮下及靜脈等注射,也可通過(guò)基因槍發(fā)送至細(xì)胞內(nèi)[3]。不同DNA導(dǎo)入方式會(huì)引起輔助性T淋巴細(xì)胞(Th)向不同方向極化,產(chǎn)生免疫應(yīng)答的機(jī)制與效果不同[4-5]。這種差異除了DNA疫苗本身抗原的特異性以外,其主要原因是DNA疫苗給藥后,絕大多數(shù)的DNA在轉(zhuǎn)運(yùn)到靶細(xì)胞前被機(jī)體的酶類降解失活,裸露質(zhì)粒DNA轉(zhuǎn)運(yùn)到胞內(nèi)的效率低下,從而導(dǎo)致免疫效果不夠理想[6-7]。因此,提高DNA疫苗轉(zhuǎn)運(yùn)至靶細(xì)胞的效率,是研制高效DNA疫苗的一個(gè)方向。目前,通過(guò)PLGA、海藻酸鈉、殼聚糖及其衍生物等可生物降解材料包裹DNA疫苗來(lái)提高效率[8-9]。
T7噬菌體是感染腸桿菌的烈性噬菌體,其顆粒結(jié)構(gòu)穩(wěn)定能耐受極端的理化環(huán)境,在體液環(huán)境中保持結(jié)構(gòu)完整從而有效保護(hù)衣殼內(nèi)部的基因組不受損傷[10]。噬菌體顆粒的不對(duì)稱性有利于募集T細(xì)胞,使其被免疫細(xì)胞識(shí)別、吞噬、降解;該過(guò)程能夠有效地將噬菌體基因組直接轉(zhuǎn)運(yùn)到免疫細(xì)胞內(nèi)部,是轉(zhuǎn)運(yùn)DNA疫苗的優(yōu)良運(yùn)輸工具。通過(guò)同源重組方法將真核表達(dá)盒插入到T7噬菌體基因組非編碼區(qū),使其隨噬菌體基因組復(fù)制而復(fù)制,構(gòu)建出遺傳穩(wěn)定的重組噬菌體。利用重組T7噬菌體作為真核表達(dá)盒的轉(zhuǎn)運(yùn)工具,既能保護(hù)DNA不受酶類物質(zhì)的降解破壞,又能誘發(fā)細(xì)胞吞噬從而直接將DNA送入免疫細(xì)胞內(nèi)部提高抗原遞呈的靶向性。本研究在T7噬菌體基因組左側(cè)非編碼區(qū)插入表達(dá)EGFP標(biāo)簽蛋白質(zhì)的真核表達(dá)盒,建立構(gòu)建轉(zhuǎn)運(yùn)真核表達(dá)盒重組噬菌體的方法,并驗(yàn)證EGFP蛋白質(zhì)的表達(dá)情況,為T7噬菌體用于DNA疫苗的轉(zhuǎn)運(yùn)奠定基礎(chǔ)。
1 材料與方法
1.1 試驗(yàn)材料
菌株與質(zhì)粒:T7 Select 415-1b Cloning Kit購(gòu)于Merck公司;pUC-19、pEGFP-N1質(zhì)粒由國(guó)家獸用生物制品工程技術(shù)研究中心實(shí)驗(yàn)室保存。
主要試劑:限制性內(nèi)切酶、TaqDNA聚合酶、T4 DNA連接酶、反轉(zhuǎn)錄酶均為大連寶生物公司產(chǎn)品;脂質(zhì)體2000為Invitrogen公司;其余試劑均為分析純。
主要儀器:超高速離心機(jī)購(gòu)自美國(guó)Beck-man公司;BTX電轉(zhuǎn)化儀、PCR儀購(gòu)自大連寶生物公司;Geldoc-It Imaging System購(gòu)自美國(guó)UVP公司;熒光顯微鏡購(gòu)自德國(guó)蔡司公司。
1.2 試驗(yàn)小鼠
6周齡ICR小鼠購(gòu)于南京醫(yī)科大學(xué)試驗(yàn)動(dòng)物中心,飼養(yǎng)于江蘇省農(nóng)業(yè)科學(xué)院實(shí)驗(yàn)動(dòng)物中心。
1.3 插入位點(diǎn)的選擇
利用DNAstar軟件比較T7 Select 415-1b與T7野生型噬菌體基因組,選擇缺失位點(diǎn)用于真核表達(dá)盒的插入。
1.4 同源重組質(zhì)粒載體構(gòu)建
以T7 Select 415-1b噬菌體基因組為模板,PCR擴(kuò)增插入位點(diǎn)左側(cè)400 bp(L)和右側(cè)150 bp(R)基因片段作為同源臂,并將其插入pUC-19質(zhì)粒載體構(gòu)建重組質(zhì)粒pUC-L-R。以真核表達(dá)載體pEGFP-N1為模板,PCR擴(kuò)增真核啟動(dòng)子(Pcmv)、EGFP、polyA尾巴等元件并進(jìn)行順序拼接成完整真核表達(dá)盒(Eukaryotic expression box,EEB),將其插入重組質(zhì)粒pUC-L-R構(gòu)建同源重組質(zhì)粒載體pUC-LEEB-R,酶切鑒定正確的克隆送樣測(cè)序鑒定。
1.5 重組噬菌體構(gòu)建與鑒定
將同源重組質(zhì)粒pUC-L-EEB-R導(dǎo)入T7噬菌體宿主E.coliBL21,挑取單菌落重組BL21細(xì)菌,過(guò)夜培養(yǎng)。T7 Select 415-1b噬菌體稀釋至103PFU/ml,分別取10 μl T7噬菌體和BL21重組菌,加到1 ml LB培養(yǎng)液中,37℃培養(yǎng)3 h。共培養(yǎng)液做10倍比稀釋,雙層瓊脂夾心法測(cè)定滴度,挑取單克隆噬菌體斑,用EGFP上、下游引物做菌落PCR鑒定,篩選陽(yáng)性噬菌體斑即為同源重組噬菌體,命名為T7-EEB,提取重組噬菌體基因組酶切鑒定。
1.6 EGFP表達(dá)與鑒定
1.6.1 體外表達(dá)鑒定 將處于對(duì)數(shù)生長(zhǎng)期的Vero細(xì)胞經(jīng)胰酶消化后用無(wú)抗生素含10%胎牛血清(FCS)的MEM培養(yǎng)液輕輕吹下,調(diào)整細(xì)胞密度到1 ml 1.0×106個(gè),然后均勻分布于6孔細(xì)胞培養(yǎng)樣板,用不含抗生素的培養(yǎng)基調(diào)整體積到2 ml每孔,于二氧化碳培養(yǎng)箱37℃過(guò)夜培養(yǎng)。參照Invitrogen公司脂質(zhì)體2000操作說(shuō)明,將提取的重組T7-EEB噬菌體基因組、同源重組質(zhì)粒載體pUC-L-EEB-R分別轉(zhuǎn)染Vero細(xì)胞,培養(yǎng)24 h后熒光顯微鏡觀察。
1.6.2 體內(nèi)表達(dá)鑒定 15只小鼠隨機(jī)分為3組,每組5只,即pUC-L-EEB-R質(zhì)粒免疫組、T7-EEB重組噬菌體免疫組和空白對(duì)照組。于0周免疫組小鼠左后腿肌肉注射鹽酸布比卡因50 μl(0.5 mg/ml)預(yù)處理,3 d后同部位注射pUC-L-EEB-R質(zhì)粒DNA 100 μl(1 μg/μl)和T7-EEB重組噬菌體顆粒100 μl(1×1011PFU/ml);14 d后同等劑量加強(qiáng)免疫1次。分別于首免疫和二免后7 d經(jīng)活體成像系統(tǒng)(南京大學(xué)生命科學(xué)學(xué)院協(xié)助)檢測(cè)EGFP表達(dá)情況;并在二免后14 d處死小鼠,采集肝臟、脾臟提取總RNA,RT-PCR檢測(cè)EGFP基因mRNA轉(zhuǎn)錄水平。
2 結(jié)果
2.1 插入位點(diǎn)及構(gòu)建流程
對(duì)比T7 Select 415 1b噬菌體與野生型T7噬菌體基因組,發(fā)現(xiàn)T7 Select 415 1b基因組有4個(gè)缺失位點(diǎn),其中在左側(cè)578~579 bp有2 168 bp缺失,適合插入外源基因。選擇578 bp處作為插入位點(diǎn),以同源重組的方式將真核表達(dá)盒導(dǎo)入,操作流程見圖1。
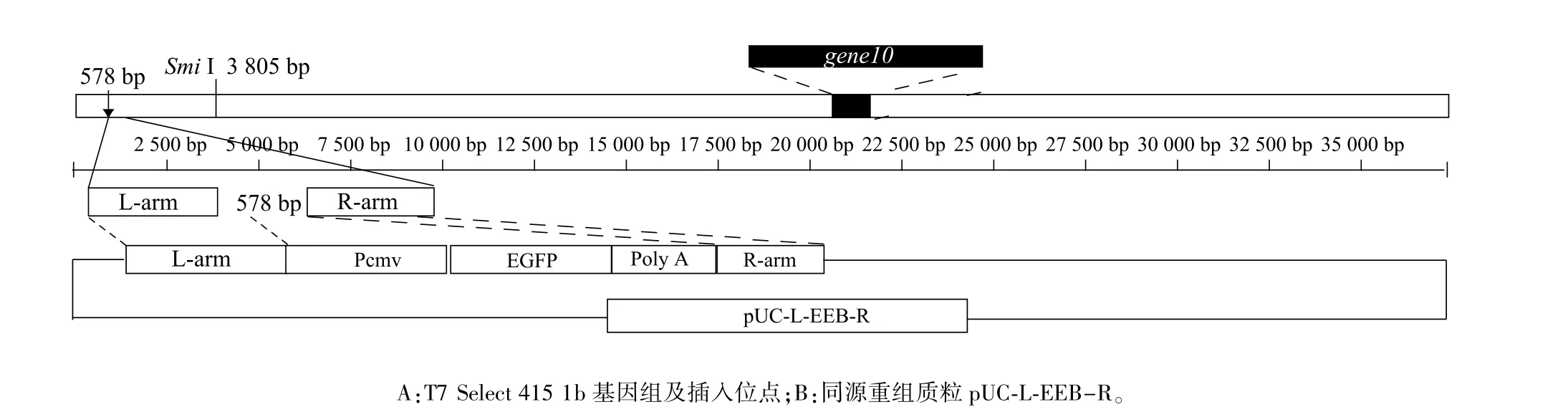
圖1 同源重組示意圖Fig.1 A schematic diagram of homologous recombinanation
2.2 同源重組質(zhì)粒鑒定
構(gòu)建的同源重組質(zhì)粒載體pUC-L-EEB-R分別用左側(cè)同源臂5′端XhoⅠ和右側(cè)同源臂3′端BamHⅠ雙酶切鑒定,觀察到長(zhǎng)度約2 200 bp的預(yù)期條帶,測(cè)序結(jié)果進(jìn)一步證實(shí)所構(gòu)建的載體與設(shè)計(jì)一致。表明該同源重組質(zhì)粒載體pUC-L-EEB-R構(gòu)建成功(圖2)。
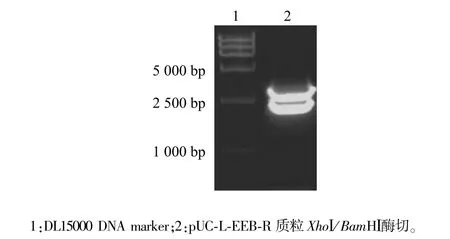
圖2 同源重組質(zhì)粒載體酶切鑒定Fig.2 Identification of homologous recombinant vector by enzyme digestion
2.3 重組噬菌體篩選與鑒定
挑取待鑒定噬菌斑,用針對(duì)EGFP基因的上下游引物做菌落PCR,5個(gè)噬菌斑擴(kuò)增出3個(gè)720 bp目的條帶,初步篩選到插入真核表達(dá)盒的重組噬菌體T7-EEB(圖3)。擴(kuò)繁T7-EEB重組噬菌體并提取基因組,以位于T7 Select 415 1b噬菌體基因組左側(cè)3 805位的SmiⅠ位點(diǎn)進(jìn)行酶切鑒定。T7 Select 415 1b基因組酶切獲得3 800 bp目的條帶,而T7-EEB基因組酶切獲得5 000 bp目的條帶,條帶大小符合預(yù)期(圖4)。通過(guò)菌落PCR初步篩選到陽(yáng)性重組噬菌體,酶切鑒定確定該重組噬菌體成功插入真核表達(dá)盒。
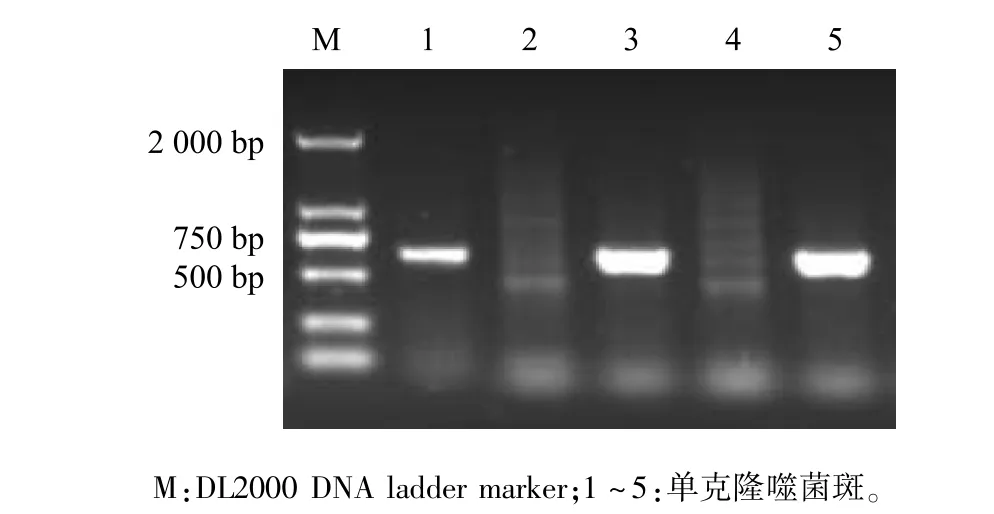
圖3 重組噬菌體PCR鑒定Fig.3 Identification of recombinant phage by PCR
2.4 T7-EEB噬菌體基因組體外表達(dá)
構(gòu)建的真核表達(dá)盒能夠在真核細(xì)胞中表達(dá)EGFP標(biāo)簽蛋白質(zhì)(圖5)。同源重組質(zhì)粒pUC-L-EEB-R(圖5A)轉(zhuǎn)染效率較高,視野內(nèi)可見多個(gè)細(xì)胞表達(dá);T7-EEB基因組(圖5B)分子量較大,轉(zhuǎn)染效率較為低下,僅單個(gè)細(xì)胞可見蛋白質(zhì)表達(dá);細(xì)胞空白對(duì)照(圖5C)未見顯色,沒(méi)有蛋白質(zhì)表達(dá)。
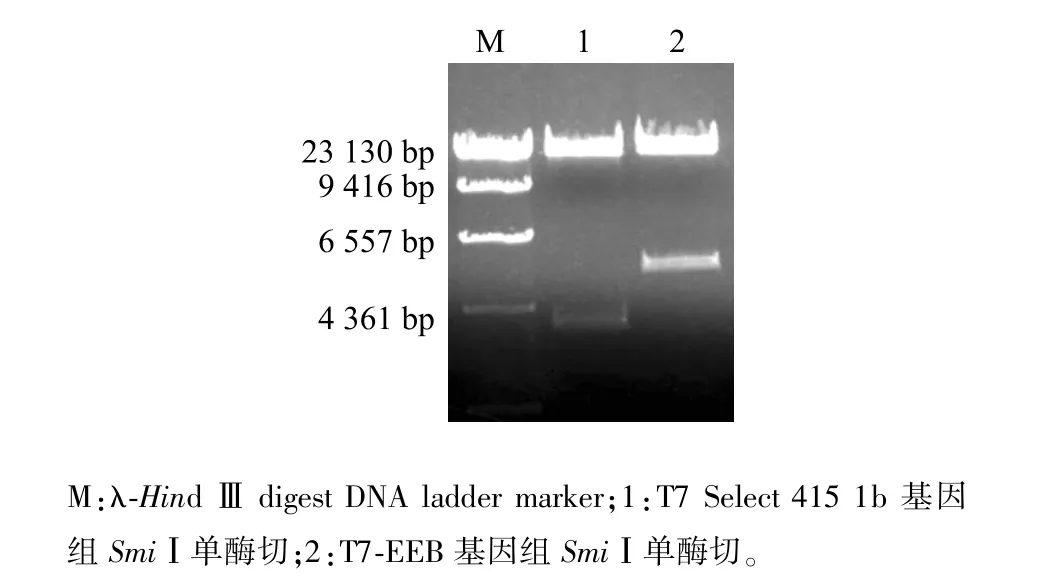
圖4 噬菌體基因組酶切鑒定Fig.4 Identification of phage genome by enzyme digestion
2.5 T7-EEB介導(dǎo)體內(nèi)表達(dá)
首次免疫后7 d經(jīng)活體成像系統(tǒng)檢測(cè)EGFP蛋白質(zhì)表達(dá)情況,T7-EEB免疫小鼠能夠檢測(cè)到EGFP蛋白質(zhì)發(fā)射信號(hào),而pUC-L-EEB-R免疫后未能檢測(cè)到信號(hào)(圖6A);在二次免疫后7 d,T7-EEB和pUC-L-EEB-R免疫小鼠均能檢測(cè)到信號(hào)(圖6B),空白對(duì)照均未檢測(cè)到信號(hào)(圖6A和圖6B)。說(shuō)明T7-EEB免疫后EGFP蛋白質(zhì)的表達(dá)比pUCL-EEB-R免疫后更加迅速。
二次免疫后取小鼠肝臟、脾臟提取總RNA,RTPCR檢測(cè)編碼EGFP蛋白質(zhì)的mRNA轉(zhuǎn)錄水平。結(jié)果(圖7)顯示,電泳條帶可見T7-EEB免疫組mRNA轉(zhuǎn)錄水平高于pUC-L-EEB-R免疫組,空白對(duì)照未能擴(kuò)增出目的條帶。
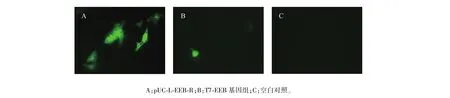
圖5 EGFP真核表達(dá)Fig.5 Eukaryotic expression of EGFP
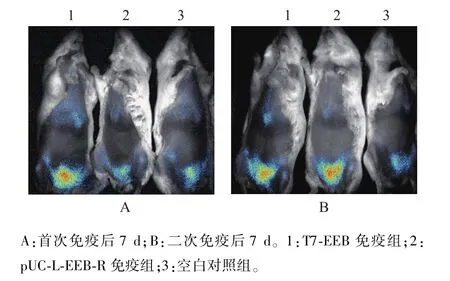
圖6 免疫小鼠活體熒光檢測(cè)Fig.6 Fluorescence images of immunized mice
3 討論
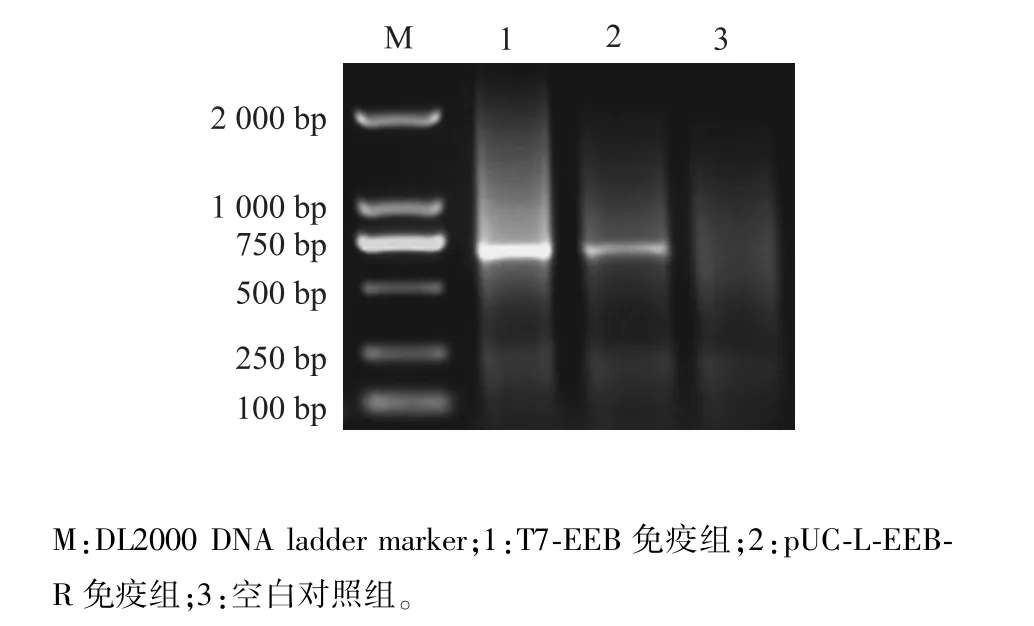
圖7 mRNA轉(zhuǎn)錄RT-PCR檢測(cè)Fig.7 Detection of mRNA transcription by RT-PCR
T7噬菌體載體Novagen公司獨(dú)家產(chǎn)品,是一種烈性噬菌體,在胞漿內(nèi)組裝子代粒子,成熟的噬菌體通過(guò)細(xì)胞裂解而釋放。該T7噬菌體載體對(duì)比野生型T7噬菌體其基因組左側(cè)578 bp處有約2 000 bp缺失,理論上可用于插入外源基因而不影響T7噬菌體生物學(xué)性狀。本研究將攜帶真核表達(dá)盒的質(zhì)粒導(dǎo)入T7噬菌體宿主細(xì)菌BL21,在噬菌體繁殖時(shí)完整基因組與質(zhì)粒的同源重組,進(jìn)而篩選出插入真核表達(dá)盒的重組噬菌體。T7噬菌體繁殖迅速,每個(gè)宿主細(xì)胞釋放約200個(gè)子代噬菌體粒子,本構(gòu)建方法沒(méi)有引入有效的篩選標(biāo)記,如何提高篩選幾率決定該方法的可操作性。本研究中以極低的感染復(fù)數(shù)感染宿主細(xì)胞,增加T7噬菌體在宿主細(xì)菌中的循環(huán)次數(shù)進(jìn)而提高同源重組幾率。重組T7噬菌體由于復(fù)制負(fù)擔(dān)增加,在平皿上表現(xiàn)出較小的噬斑形態(tài),挑選形態(tài)較小的噬斑進(jìn)行菌落PCR鑒定能夠提高篩選陽(yáng)性率。本研究從5個(gè)單噬斑中篩選到3個(gè)疑似陽(yáng)性克隆,證明同源重組方法構(gòu)建重組噬菌體具有可行性。
利用完整噬菌體粒子作為真核表達(dá)盒的轉(zhuǎn)運(yùn)載體是一個(gè)比較新的途徑。在這個(gè)過(guò)程中,克隆到噬菌體基因組中的真核表達(dá)盒攜帶保護(hù)性抗原基因,以完整的噬菌體粒子免疫動(dòng)物,在噬菌體被免疫細(xì)胞吞噬、裂解后釋放出的真核表達(dá)盒利用宿主細(xì)胞表達(dá)元件啟動(dòng)保護(hù)性抗原的表達(dá)。Clark等[11-13]用包含乙肝表面抗原表達(dá)盒的完整的λ噬菌體顆粒免疫動(dòng)物發(fā)現(xiàn)其免疫保護(hù)效果是裸露的核酸疫苗的10倍左右。噬菌體轉(zhuǎn)運(yùn)真核表達(dá)盒與傳統(tǒng)質(zhì)粒DNA疫苗傳遞系統(tǒng)相比在對(duì)核酸酶的穩(wěn)定性方面有明顯優(yōu)勢(shì);而且噬菌體顆粒被免疫細(xì)胞吞噬,抗原遞呈效率更高。本研究中,基因組的體外瞬時(shí)轉(zhuǎn)染證明噬菌體攜帶的真核表達(dá)盒可以實(shí)現(xiàn)蛋白質(zhì)的表達(dá),由于T7-EEB基因組分子量遠(yuǎn)大于pUC-LEEBR,其轉(zhuǎn)染效率較低所表達(dá)的EGFP蛋白量也相對(duì)較少。相反,T7-EEB免疫組相比pUC-L-EEB-R免疫組,其EGFP表達(dá)更為迅速、表達(dá)量更高,間接說(shuō)明噬菌體顆粒轉(zhuǎn)運(yùn)真核表達(dá)盒作為DNA疫苗免疫的新途徑效率更高效果更好。
噬菌體進(jìn)入體內(nèi)會(huì)很快被循環(huán)系統(tǒng)清除,但是在脾臟可以檢測(cè)到存活的噬菌體,其存活周期可以達(dá)到14~21 d[14]。在噬菌體治療過(guò)程中,兔子口服噬菌體后血漿中能檢測(cè)到活性噬菌體,并且能夠檢測(cè)到針對(duì)噬菌體衣殼蛋白質(zhì)的特異性體液免疫反應(yīng)[15]。本研究中所用T7噬菌體衣殼蛋白質(zhì)p10B還可以用于外源多肽的表面展示,可在其表面展示抗原表位激發(fā)體液免疫反應(yīng),而基因組中嵌合的真核表達(dá)盒可以表達(dá)蛋白質(zhì)激發(fā)細(xì)胞免疫反應(yīng),可以進(jìn)一步升級(jí)該系統(tǒng)達(dá)到體液、細(xì)胞免疫的雙重激發(fā)。因此,基于該理念開發(fā)噬菌體疫苗并且可以通過(guò)口服途徑進(jìn)行免疫,將會(huì)是一種具有良好前景的疫苗研發(fā)策略。
[1] MA L.DNA vaccines:an historical perspective and view to the future[J].Immunol Rev,2011,239(1):62-84.
[2] MICHELE A K,DAVID B W.DNA vaccines:ready for prime time[J].Nature Reviews Genetics,2008(9):776-788.
[3] PASETTI M F,BARRY E M,LOSONSKY G,et al.AttenuatedSalmonella entericaserovar Typhi andShigella flexneri2a strains mucosally deliver DNA vaccines encoding measles virus hemagglutinin,inducing spcific immune responses and protection in cotton rats[J].J Virol,2003,77(9):5209-5217.
[4] PACHUK C J,MCCALLUS D E,WEINER D B,et al.DNA vaccines-challenges in delivery[J].CurrOpin Mol Therapy,2000,2 (2):188.
[5] DUBENSKY T W,LIU M A,ULMER J B.Delivery systems for gene based vaccines[J].Mol Med,2000,6(9):723.
[6] ERTL H C,XIANG Z.Novel vaccine approaches[J].J Immunol,1996,156(10):3579-3582.
[7] FYNAN E F,WEBSTER R Q,FULLER D H,et al.DNA vaccines:Protectivev immunizations by parenteral mucosal,and genegun inoculations[J].Proc Natl Acad Sci USA,1993,90(24): 478-482.
[8] HAO T,MCKEEVER U,HEDEY M L.Biological potency of microsphere encapsulated plasmid DNA[J].J Controlled Release, 2000,69:249.
[9] GUPTA R K,CHANG A C,SIBER G R.Biodegradable polymer microspheres as vaccine adjuvants and delivery systems[J].Deve BiolStand,1998,92:63.
[10]徐 海,王義偉,鮑 熹,等.表面展示GnRH重組T7噬菌體構(gòu)建及其免疫效果[J].江蘇農(nóng)業(yè)學(xué)報(bào),2014,30(4):809-813.
[11]CLARK J R,MARCH J B.Bacteriophage-mediated nucleic acid immunization[J].FEMS Immunol Med Microbiol,2004,40 (1):21-26.
[12]CLARK J R,MARCH J B.Bacterial viruses as human vaccines? [J].Expert Rev Vaccines,2004,3(4):463-476.
[13]CLARK J R,BARTLEY K,JEPSON C D,et al.Comparison of a bacteriophage-delivered DNA vaccine and a commercially available recombinant protein vaccine against hepatitis B[J].FEMS Immunol Med Microbiol,2011,61(2):197-204.
[14]KELLER R,ENGLEY F B.Fate of bacteriophage particles introduced into mice by various routes[J].Proc Soc Exp Biol Med, 1958,1:577-580.
[15]REYNAUD A,CLOASTRE L,BERNARD J,et a1.Characteristics and diffusion in the rabbit of a phage forEscherichia coli0103.Attempts to use this phage for therapy[J].Vet Microbial, 1992,30(2-3):203-212.
(責(zé)任編輯:袁 偉)
Construction ofrecombinantbacteriophageT7deliveringeukaryotic expression box
XU Hai, BAO Xi, WANG Yi-wei, LU Yu, XU Meng-wei, HOU Ji-bo
(Jiangsu Academy of Agricultural Sciences/National Veterinary Biological Medicine Engineering Research Center,Nanjing 210014,China)
To construct a recombinant T7 phage delivering eukaryotic expression box and to detect the EGFP(enhanced green fluorecent protein)tag protein expressionin vivoandin vitro,respectively,a 400-bp fragment(L)located at the upstream of 578 bp of T7 genome and a 200-bp fragment(R)located at the downstream were used as homologous arms. Eukaryotic expression box encoding EGFP protein(EEB)was inserted into left and right arm to construct homologous recombinant plasmid vector pUC-L-EEB-R.Plasmid vector was transformed into bacterium BL21 where homologous recombinant took place during the T7 phage propagation.EGFP protein expression was identified by phage genome transfection and phage particle immunization.PCR and restriction enzyme digestion analysis demonstrated that eukaryotic expression box was inserted into T7 phage genome correctly.Visible EGFP protein expression was detected bothin vivoandin vitro.In mice liver and spleen,mRNA transcription ofEGFPgene was detected by RT-PCR as well.These results indicate that homologous recombinanation is capable to construct recombinant T7 phage which could deliver eukaryotic expression box and achieve protein expression.
homologous recombination;DNA vaccine;eukaryotic expression;T7 phage
Q939.48
A
1000-4440(2015)01-0117-05
徐 海,鮑 熹,王義偉,等.轉(zhuǎn)運(yùn)真核表達(dá)盒的重組T7噬菌體構(gòu)建[J].江蘇農(nóng)業(yè)學(xué)報(bào),2015,31(1):117-121.
10.3969/j.issn.1000-4440.2015.01.018
2014-08-07
江蘇省農(nóng)業(yè)科技自主創(chuàng)新基金項(xiàng)目[CX(11)4073]
徐 海(1982-),男,江蘇揚(yáng)州人,碩士研究生,主要從事預(yù)防獸醫(yī)學(xué)方面的研究。(Tel) 13645161013;(E-mail)hai_x @126.com
侯繼波,(E-mail)houjibo@jaas.ac.cn
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級(jí))(2025年8期)2025-08-18 00:00:00
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
學(xué)苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
小學(xué)生學(xué)習(xí)指導(dǎo)(低年級(jí))(2017年10期)2017-10-10 01:00:05
海峽科技與產(chǎn)業(yè)(2016年3期)2016-05-17 04:32:12
云南中醫(yī)學(xué)院學(xué)報(bào)(2014年3期)2014-07-31 18:57:34
